
Abstract
There is growing public concern surrounding traumatic brain injury (TBI). TBI can cause significant morbidity, and the long-term sequelae are poorly understood. TBI diagnosis and management rely on patient-reported symptoms and subjective clinical assessment. There are no biologic tools to detect mild TBI or to track brain recovery. Emerging evidence suggests that microRNAs (miRNAs) may provide information about the injured brain. These tiny epigenetic molecules are expressed throughout the body. However, they are particularly important in neurons, can cross the blood-brain barrier, and are securely transported from cell to cell, where they regulate gene expression. miRNA levels may identify patients with TBI and predict symptom duration. This review synthesizes miRNA findings from 14 human studies. We distill more than 291 miRNAs to 17 biomarker candidates that overlap across multiple studies and multiple biofluids. The goal of this review is to establish a collective understanding of miRNA biology in TBI and identify clinical priorities for future investigations of this promising biomarker.
Introduction
Head injuries are a common concern in the United States, especially among children and adolescents, whose developing brains may be at an increased risk for long-term sequelae. It is estimated that 20% of adolescents 1 have been diagnosed with at least 1 concussion—the sequelae of symptoms induced by mild traumatic brain injury (TBI). 2 Despite growing public concern about concussion, there are limited tools available for diagnosis and prognosis. 3 Currently, concussion diagnosis is based on subjective assessment. 3 There are few biologic measures available to personalize anticipatory guidance or therapeutic decision-making. A lack of sensitive, objective assessment tools hinders the ability of physicians to provide accurate diagnoses and effective treatments.4,5 The resulting one-size-fits-all approach may increase anxiety about brain health for patients and families.6,7 Thus, establishing mild traumatic brain injury (mTBI) biomarkers could have profound clinical impact on the diagnosis and management of concussion.8,9 Emerging evidence suggests that microRNAs (miRNAs) may fill this clinical need.
miRNAs are implicated in the pathophysiology of many diseases, but they are critical for neurodevelopment and brain function. 10 Through the regulation of gene activity, miRNAs control cellular processes essential to neuronal injury and repair: differentiation, proliferation, apoptosis, and metabolism.11-13 Disruption in miRNA levels have been reported in numerous disease processes of the central nervous system (CNS).14-16 Here, we review the miRNAs altered in human patients with mild, moderate, and severe TBI, identifying the most consistent findings and proposing avenues for future investigations to validate this promising molecular technology.
Clinical Aspects of TBI
Neuropathology
TBI is defined as an alteration in brain function, or other evidence of brain pathology, caused by an external force. 17 The external force can lead to permanent or temporary injuries from the neuronal insult. Primary damage is characterized by the effects of the immediate impact on neuroanatomy and function, whereas secondary damage consists of the biochemical responses to injury that result in neuronal repair or apoptotic cell death.18-21 miRNAs are implicated in both the primary and secondary damage responses to TBI. 22 Injured neurons may release miRNAs into the extracellular space, where their small size allows them to navigate the blood-brain barrier (BBB), facilitating peripheral sampling.23,24 In the cellular response to secondary damage,25,26 neurons use miRNA signaling to regulate synaptogenesis and neuroplasticity. As a result, miRNA profiles during the subacute period may also telegraph the trajectory of brain recovery. 27
Clinical diagnosis
The Glasgow Coma Scale (GCS) is a scoring system based on verbal, motor, and eye-opening reactions to external stimuli. GCS has been used to classify TBI as mild (GCS ⩾ 13), moderate (GCS 9-12), or severe (GCS ⩽ 8). 28 Mild, moderate, and severe TBI involve a spectrum of similar mechanical forces as well as partially overlapping pathophysiology. However, patients with these conditions often display disparate symptomology, and their diagnosis and management can entail distinct approaches. Consideration of the commonalities and differences across TBI severity may help inform biomarker development.
The most common type of TBI is mTBI, accounting for approximately 80% of cases. 29 Diagnosis of mTBI is complex, requiring a multi-modal approach. Recent guidelines from the Centers for Disease Control (CDC) recommend neuropsychological tools and age-appropriate symptom scales alongside standard clinical assessment for mTBI diagnosis. 3 Although cognition and balance measures may be used, routine use of imaging modalities is discouraged, and objective biomarkers are not recommended outside of a research setting. Recently, the Food and Drug Administration (FDA) approved serum measurements of ubiquitin C terminal hydrolase (UCH-L1) and glial fibrillary acidic protein (GFAP) to identify adult patients at risk for intracranial injury.30,31 The utility of these measures is limited to differentiating patients with moderate-severe TBI who benefit from cranial imaging. This may be because protein biomarkers require moderate-to-severe CNS damage for reliable peripheral detection.32-34 In contrast, miRNAs can be released as signaling vectors by live cells35,36 and do not require cell injury for release. This may make them more amenable to mTBI detection.37-39 This idea is supported by findings that peripheral miRNA levels reflect subtle structural changes in the CNS identified by magnetic resonance imaging. 40
Clinical symptoms
Even mTBI can impact cognition, sleep, mood, or physical wellbeing (eg, fatigue, nausea, dizziness, and headache). These sequelae are commonly called “concussion,” a term used herein when stressing the patient-oriented experience of mTBI symptoms. 41 Most of the neurologic, cognitive, and behavioral changes associated with mTBI resolve in days to weeks, but for 10% to 30% of those affected, symptoms can persist for months.42-44 Symptom burden at initial presentation45,46 and clinical prediction tools47,48 have demonstrated fair utility for identifying patients at risk for prolonged concussion symptoms, but there are no objective measures currently available. A recent study of saliva miRNA levels in children shows promise for predicting prolonged concussion symptoms. 49
Clinical management
Because no single tool strongly predicts outcome in patients with mTBI, physicians are forced to rely on subjective symptom surveys to formulate management plans. 3 Such surveys may be manipulated by patients who wish to expedite return-to-play, 50 or prolong return-to-work/school activities. 51 Although management plans may be partially informed by individual risk factors (eg, previous mTBI, learning difficulties, psychosocial stressors), most patients receive standard anticipatory guidance about sleep, exercise, and headache management. The expectation is that 70% to 80% of patients will recover within 1 month. 3 In patients with protracted recovery, referral for multi-disciplinary evaluation is recommended. Thus, patients with persistent headache or dizziness may wait a full month for individualized therapy, during which a critical window for neuroplasticity and intervention may have passed. Currently, there are no specific therapies for mTBI, and many therapeutic trials have provided inconclusive results.52,53 This may result because mTBI is a heterogeneous condition and therapeutic efficacy depends on the recognition of phenotypes and early, personalized intervention. 54 The biological diversity and pathologic specificity of miRNAs make them ideal candidates for mTBI phenotyping. Indeed, studies have shown that concussion-related miRNAs measured at the time of injury are associated with chronic symptom characteristics. 49
TBI biomarkers
Biomarkers, such as proteins, metabolites, and neuronal imaging, have been extensively explored in patients across the spectrum of TBI severity. 9 In particular, studies of protein biomarkers as a means for diagnosing, monitoring, and predicting the course of concussions has increased markedly over the past decade. 55 Here we briefly focus on 2 proteins with clinical approval.30,31,46 GFAP and UCH-L1 have demonstrated utility for discerning adult patients with TBI who may benefit from neuroimaging.56-59 Protein biomarkers may require BBB disruption to migrate from cerebral spinal fluid (CSF) into blood. 60 As a result, the sensitivity of protein biomarkers for TBI detection may be limited in individuals with mTBI. 61 Second, unlike miRNAs, which are packaged in protective exosomes, 62 extracellular proteins may be subject to degradation by endogenous proteases. 61 This could potentially limit their utility to acute and subacute periods of TBI. Finally, a number of protein biomarkers are released following damage to myocytes and osteocytes, and this could limit their specificity in patients with poly-trauma. 63 This is not to say that protein biomarkers have no role in TBI assessment. Rather, we wish to highlight potential limitations that miRNAs may be uniquely suited to address.
miRNA Physiology
miRNA processing
miRNAs are short (19-28 nucleotides), endogenous, non-coding RNAs that act at the post-transcriptional level to regulate protein synthesis. 64 They are initially transcribed in the nucleus as long (>1000 base pairs) precursor molecules called primary microRNA (pri-miRNA) which fold into a hairpin structure. The pri-miRNA is trimmed by the RNase Drosha into stem-loop structures called precursor microRNA (pre-miRNA) that are 60 to 100 nucleotides long. The pre-miRNA is exported into the cytoplasm where the miRNA processing enzyme Dicer cleaves them into miRNA:miRNA duplexes. One half of this duplex will become a mature miRNA molecule, whereas the other half, known as the minor miRNA, will be degraded.65,66 The mature miRNA strand is bound within the microRNA-induced silencing complex (miRISC) where it suppresses the expression of specific gene targets by inhibiting protein translation or promoting messenger RNA (mRNA) degradation. 67 The miRISC complex recognizes target genes through “seed sequences” at the 5′ end of the miRNA that are complementary to the 3′ region of the intended mRNA target. The seed sequence of the miRNA can recognize hundreds of different mRNAs, and there are several miRNAs that share the same seed sequence. 68 miRNAs are organized into families based on similarities in the sequence of the hairpin molecules. 69
miRNAs in the brain
The CNS contains the highest concentration and highest diversity of miRNAs. It is estimated that 70% of all miRNAs are expressed in the brain, spinal cord, or peripheral nerves. 70 Expression of miRNA evolves throughout neurodevelopment and varies across the brain regions. 71 Within neurons, miRNAs also display intracellular variation in localization. 72 Enrichment of certain miRNAs in the axonal or dendritic compartments suggests that individual miRNAs may have unique functions regulating local protein expression, synapse maturation, or neural circuit formation. 73 Expression of miRNAs is critical to the development and function of the CNS. Dysregulated miRNA levels have been linked to impaired learning, memory, and cognition as well as a host of neuropsychiatric disorders. 74 These characteristics have led to rising interest in the utility of miRNAs as biomarkers for CNS pathology. 75
The role of miRNAs in neuronal injury and repair
Numerous studies have examined physiologic alterations in miRNA expression after TBI. Redell et al 22 first described altered levels of hippocampal miRNAs in rats exposed to controlled cortical impacts. Using microarray techniques to interrogate more than 400 miRNAs, they verified changes in 8 miRNAs with quantitative real-time polymerase chain reaction (RT-PCR; miR-107, miR-130a, miR-223, miR-292-5p, miR-433-3p, miR-451, miR-541, and miR-711). Liu et al 26 extended miRNA findings to the rat cerebral cortex and established that miRNA levels change dynamically during the first 72 hours after TBI. Notably, the study identified 1 miRNA (miR-21) which remained chronically elevated following the initial injury.
The biochemical cascade that occurs following brain injury consists of mechanical insult, oxidative stress, apoptotic cell death, subacute repair, and chronic remodeling. 76 miR-21, a miRNA biomarker identified in both human and animal TBI studies, may inhibit apoptosis and target angiogenesis factors critical to BBB maintenance.77,78 miR-16 is another biomarker candidate identified in animal studies that may regulate apoptosis. In addition, miR-16 has putative targets critical to cell cycle regulation, including Bcl-2 and cyclin dependent kinase 6. Together these molecules may facilitate neurogenesis and acute repair responses following TBI. 79 Unlike proteins, which are typically increased after TBI, some miRNAs undergo down-regulation after injury (eg, miR-107, mir-27a). Decreases in miR-107 may be critical for inflammatory processes, allowing transcription of granulin. 80 Whereas decreases in miR-27a may facilitate programmed cell death by allowing expression of pro-apoptotic Bcl-2 proteins. 81
The exact mechanisms that drive individual miRNA alterations after TBI are not fully understood. As with proteins, endothelial cells and astrocytes forming the BBB may release miRNAs following TBI-induced disruption. 82 In addition, some miRNAs may be released from damaged neurons as a result of the shearing forces that occur in axonopathy. 83 Although peripheral transport of neuron-derived exosomes containing miRNA has not been firmly established, several studies suggest that nucleic acid content can cross the BBB with the help of vesicular transport.84,85 Glia also express miRNAs, and may alter their expression in response to oxidative, metabolic, and apoptotic demands of traumatic injury. 86 Peripheral miRNAs can also undergo concentration shifts in response to sympathetic, hormonal, or neuro-immune mechanisms that regulate the physiologic response to TBI. In this latter scenario, miRNAs could play an important role in neuroplasticity, 87 and differential miRNA expression among TBI patients in the subacute period could portend long-term prognosis. Whether miRNA changes are passive artifacts of primary injury or integral players in the secondary injury response, their dynamic expression and versatile role in the human brain provides a distinct opportunity to gauge the CNS response to trauma. Furthermore, the same properties that allow miRNA cross-talk between neurons (small size, vesicle protection) may permit sampling of these brain-related molecules in peripheral biofluids.
Peripheral measurement of brain-related miRNAs
miRNAs can be transported through the extracellular space by exosomes and microvesicles, or bound to protective proteins such as high-density lipoprotein. 88 Such carriers facilitate protective transfer and targeted docking of miRNAs at distant cells, where they may traverse the cell membrane and influence gene expression. 89 Protection by exosomes and microvesicles also permits ease of miRNA measurement in various biofluids. miRNAs can be quantified in serum, plasma, CSF, urine, and saliva.61,90 Due to their abundance, stability at fluctuating pH levels, resistance to enzymatic degradation, and essential role in transcriptional regulation, miRNAs make ideal biomarker candidates. 91 Brain-related miRNAs have been detected in serum less than 1 hour after TBI. 92
miRNAs as Biomarkers in Human TBI
Overlapping miRNAs in human TBI
This review summarizes findings from 14 studies that examined miRNA levels in human patients with TBI. Together, the 14 studies have identified a host of miRNA candidates that may herald CNS injury across a spectrum of severity and time (Supplemental Figure 1). Although most of the studies explored miRNA levels in peripheral blood,23,38,39,61,92-99 a handful of recent investigations have also identified saliva as a novel and easily accessible biofluid with potential CNS overlap.37,38,49 Cumulatively, the 14 studies distinguish 291 miRNAs with prospective roles in TBI. Of these, 17 miRNAs are altered in multiple studies, across 3 biofluids (CSF, blood, and saliva; Figure 1A and B). A host of factors may contribute to this heterogeneity, including TBI severity, timing of sample collection, biofluid chosen, method of RNA analysis, and participant ages. Below, we consider such factors and attempt to untangle the web of biomarker complexity pervading miRNA research. The goal of this review is to identify gaps in existing miRNA literature that can be addressed through cohesive methodological approaches to promote valid, reliable results with near-term clinical applications.

(a) Acute injury response of miRNAs: across 3 biofluids (cerebrospinal fluid, saliva, and blood) in the acute period (<96 hours post injury); (b) secondary injury response of miRNAs: across 3 biofluids (cerebrospinal fluid, saliva, and blood) in the subacute or chronic period (>5 days post injury).
The evolution of miRNA exploration in human TBI
Initial investigations of miRNA expression in TBI patients predominantly focused on serum miRNA differences between small numbers of injured participants and controls. A 2012 study by Pasinetti and colleagues used microarray to detect 13 small RNAs in blood mononuclear cells from 9 veterans with mTBI compared with 9 veterans without mTBI (73% male, mean age: 30 years). The authors reported 1 miRNA change (miR-671-5p) that was validated with PCR. 96
Another study compared vesicular RNA in the CSF of 11 severe TBI patients and 17 controls to detect differential expression (53% male, mean age: 46 years). The authors determined that most of the RNA packaged in CSF microparticles was non-coding RNA, and that 2 of these non-coding RNAs (miR-9 and miR-451) were differentially expressed in severe TBI patients. They also found that the transfer of microparticles in vitro resulted in the regulation of specific neuronal genes. 95
A study by Yang et al 94 explored serum miRNA levels in an expanded cohort of adult patients with severe TBI (n = 76), compared with healthy (n = 38) controls (mean age: 47 years). They found increased levels of miR-93, miR-191, and miR-499 in TBI patients, which peaked on days 2 to 7 post injury, and was correlated with injury severity. 94
You and colleagues described CSF miRNA levels in 26 comatose patients with severe TBI compared with 21 controls (55% male, mean age: 50 years). They detected 14 miRNAs with distinct expression levels in severe TBI patients, and 1 of these (miR-431-3p) had an associated single-nucleotide polymorphism in its promoter region. 93
Studies have also incorporated multiple biofluids to compare miRNA levels. A study by Bhomia and colleagues is notable for its use of both CSF and serum from severe TBI patients, as well as its use of controls with orthopedic injury. The authors used RT-PCR to measure miRNA from 8 patients with mTBI, 8 patients with severe TBI, 7 orthopedic controls, and 8 healthy controls. The severe TBI serum was collected on average 33.8 hours after injury, and the CSF was collected 26.3 hours after injury, on average. Those with mTBI and orthopedic controls had their serum collected on average 3.1 hours after injury. They identified 10 miRNAs up-regulated in the serum of both mild and severe TBI groups relative to the orthopedic controls: miR-151-5p, miR-328, miR-362-3p, miR-486, miR-505, miR-451, miR-30d, miR-20a, miR-195, and miR-92a. Changes of 4 of these miRNAs were validated in the CSF of severe TBI patients. 61
In 2018, Hicks and colleagues used a similar approach to identify overlapping CSF and saliva miRNA profiles among children with severe (n = 8) and mTBI (n = 60; mean age: 14 years). The authors used RNA sequencing to identify 6 salivary miRNAs with overlapping CSF alterations (miR-182-5p, miR-221-3p, miR-26b-5p, miR-320c, miR-29c-3p, and miR-30e-5p) <24 hours after injury. Furthermore, they showed that these miRNAs could predict mTBI status relative to healthy controls and that saliva miRNA levels were correlated with the severity of subjective symptoms. 37
However, this was not the first study to demonstrate the diagnostic utility of peripheral miRNA levels. A study by Redell et al 23 used RT-PCR to explore 108 miRNAs in the plasma of 10 adults with severe TBI, and 10 adults with mTBI (<10 after injury). There were 33 miRNAs increased and 19 miRNAs decreased in the plasma of TBI patients relative to healthy controls (8 were unique to severe TBI patients). Two miRNAs (miR-92a and miR-16) demonstrated diagnostic potential in both severe and mTBI relative to healthy and orthopedic injury controls. 23
Recently, a study of saliva miRNA levels from 32 rugby players (mean age: 24 years) detected 5 miRNAs (miR-27b-3p, miR-142-3p, let-7i, miR-107, and miR-135b-5p) with differential expression 48 to 72 hours after sports-related concussion. Patterns for these 5 miRNAs correlated with reaction time on ImPACT testing and were able to predict a TBI diagnosis better than 92 protein biomarker candidates. 38
Several studies have also focused on the longitudinal response of miRNAs to TBI. A study by Taheri and colleagues used RT-PCR to analyze 740 miRNAs in the serum of 38 patients (71% male, mean age: 43 years) receiving intensive care for severe TBI at 1, 7, and 28 days post injury, as well as 25 patients 5 years post injury. In patients with TBI-induced hypopituitarism, 2 miRNAs (miR-126-3p and miR-3610) demonstrated alterations during both the acute and chronic phases. 97
Di Pietro and colleagues screened 754 miRNAs in the serum of 5 mTBI, 5 severe TBI, and 5 healthy controls at 1 and 15 days post injury. They identified 10 miRNA candidates that changed over time. In 30 additional patients with TBI, 2 of the miRNA candidates (miR-425-5p and miR-502) were validated at earlier time-points (0-1 hour and 4-12 hours post injury) and 2 (miR-21 and miR-335) were validated relative to healthy and orthopedic controls. 92
Zhangjie et al 98 also examined the time course of miRNA levels following sports-related concussion in 15 athletes (relative to 8 healthy peers). At both 1 and 2 weeks post injury, serum levels of miR-425-5p were down-regulated in concussed patients and correlated with the time since injury. 98
The ability to measure dynamic miRNA responses after TBI has led some groups to posit that miRNA levels may be used to predict medical outcomes. One study measured plasma miRNA levels on arrival to the emergency department, and again 5 and 30 days post injury in adults with TBI (mean age: 43 years). The authors reported that plasma levels of miR-142-3p and miR-423-3p differentiated patients with mild head injury from those with higher post-concussive syndrome (PCS) risk, based on standardized post-traumatic amnesia testing. 39
A study published in JAMA Pediatrics by Johnson and colleagues used RNA sequencing to identify 5 saliva miRNAs (miR-320c, miR-133a-5p, miR-769-5p, let-7a-3c, and miR-1307-3p) that differentiated risk for prolonged concussion symptoms among 52 children with mTBI (58% male, mean age: 14 years). Levels of 3 miRNAs at the time of injury were also associated with the presence of specific symptoms (eg, memory difficulty, headaches, fatigue) 1 month later. 49
Recently, a large-scale study of 50 adult mixed martial arts fighters (mean age: 27 years) compared the utility of serum and saliva miRNAs for detecting hits-to-the-head, relative to protein markers. Serum and saliva were collected immediately before fight and again after fight (15 minutes, 2-3 days, 1 week, and 3-4 weeks). Saliva and serum miRNA levels showed an equivalent ability to detect the number of hits-to-the-head a fighter experienced. This ability exceeded the accuracy of serum protein markers. Intriguingly, saliva miRNA levels demonstrated a more acute response to head trauma than serum miRNA, and acute saliva miRNA levels were correlated with functional measures of cognition and balance. 99
A Sampling of Top miRNA Candidates and Their Putative Functions in TBI
miR-320c represents one intriguing biomarker candidate in patients with TBI. miR-320c is altered in the blood of adults with severe TBI and the saliva of children with mTBI.23,49,97 Furthermore, this miRNA demonstrates longitudinal variation in CSF after severe TBI and predicts the duration of TBI symptoms. Previous studies in depressed adults have found that miR-320c levels change in the cerebral cortex following suicide. 100 Such findings are intriguing given the association of prolonged concussive symptoms with depressed mood. Such an association might be facilitated by the role of miR-320c in neuroplasticity. 37 For example, an investigation of miRNA expression in neuronal precursor cells has demonstrated that miR-320c levels are critical for neuronal differentiation and axonal outgrowth. 101
Another compelling TBI biomarker candidate is miR-92a. miR-92a was up-regulated in the blood of patients with mTBI and the blood and CSF of patients with severe TBI.23,61 This microRNA was up-regulated in the acute (hours) and subacute (days) period after injury. Studies have shown that up-regulated miR-92a blocks angiogenesis after an ischemic event.102,103 Thus, acute up-regulation of miR-92a in patients with severe TBI may be critical in those with intracranial bleeding or neurovascular compromise.
Changes in the miR-30 family have been detected in several human TBI studies. Changes in miR-30 expression were noted in the blood of patients with severe TBI, both hours and days after injury. miR-30 changes have also been detected in the CSF weeks after brain injury. In patients with mTBI, subacute changes in miR-30 expression have been reported in both saliva and blood.23,38,61,93 The miR-30 family plays a role in neuroinflammation and is negatively regulated by pro-inflammatory cytokines. A decreased expression of miR-30 has been associated with changes in cell morphology, loss of cell adhesion, and increased cell migration due to tight junction disruption.104-106 Therefore, miR-30 may play a critical role in the maintenance of the BBB.
The biologic rationale for peripheral miRNA biomarkers in TBI can be inferred from the roles of their putative mRNA targets in brain-related processes. Here, we use DIANA mirPath bioinformatics along with the microT-CDS database to interrogate the 17 miRNAs implicated in the primary (hours-to-days) and secondary (days-to-weeks) damage responses to TBI (Figures 1A and B). The 17 miRNAs target 2014 mRNA transcripts with high confidence (microT-CDS ⩾ 0.975). These mRNAs demonstrate enrichment for 22 KEGG (Kyoto Encyclopedia of Genes and Genomes) pathways (false discovery rate [FDR] < 0.05), including 8 implicated in brain-related mechanisms (Table 1). The pathway targets included those specific to neuronal outgrowth (neurotrophin signaling, FDR = 0.021) and brain function (amphetamine addiction, FDR = 2.6E-05; cocaine addiction, FDR = 0.020), as well as those with broad-ranging implications in cell signaling (ErbB signaling, FDR = 0.0056). Among the 17 miRNAs, miR-27b-3p targets the largest number of transcripts from brain-related pathways (43 mRNAs, accounting for 14% of its 313 total targets). Five genes (COL27A, KMT2C, ACVR1C, ACVR2A, and ITGA5) were targeted by at least 2 of the 17 miRNAs. It is worth noting that these targets and functions are putatively based on predicted miRNA-mRNA interactions, or observed in vitro targeting experiments. Additional mechanistic studies defining the specific physiologic contributions of individual miRNAs in TBI will be required to enhance our understanding of the exact role peripheral miRNAs play in CNS injury.
Putative targets of the 17 miRNAs with the highest TBI biomarker potential.
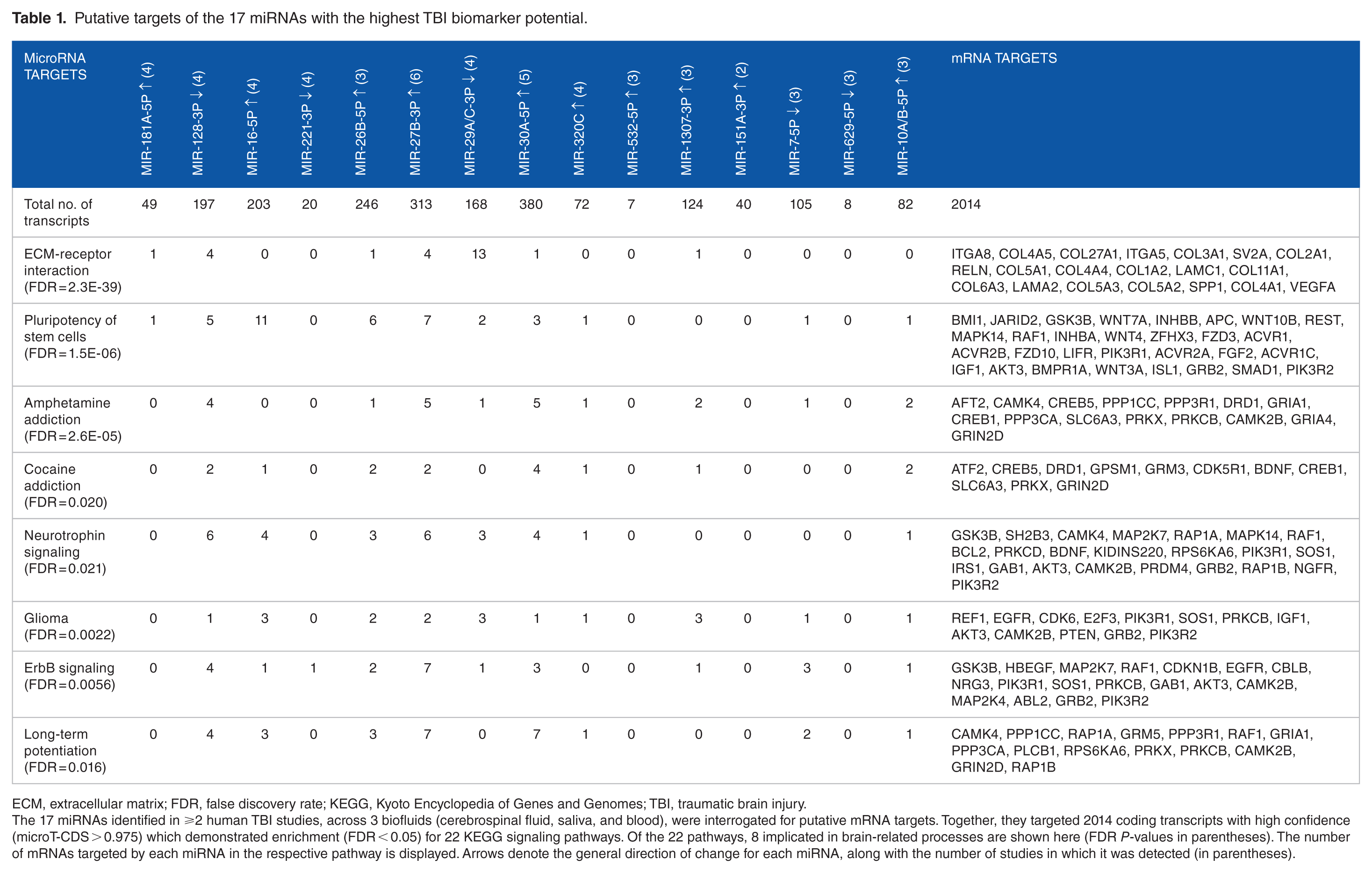
ECM, extracellular matrix; FDR, false discovery rate; KEGG, Kyoto Encyclopedia of Genes and Genomes; TBI, traumatic brain injury.
The 17 miRNAs identified in ⩾2 human TBI studies, across 3 biofluids (cerebrospinal fluid, saliva, and blood), were interrogated for putative mRNA targets. Together, they targeted 2014 coding transcripts with high confidence (microT-CDS > 0.975) which demonstrated enrichment (FDR < 0.05) for 22 KEGG signaling pathways. Of the 22 pathways, 8 implicated in brain-related processes are shown here (FDR P-values in parentheses). The number of mRNAs targeted by each miRNA in the respective pathway is displayed. Arrows denote the general direction of change for each miRNA, along with the number of studies in which it was detected (in parentheses).
A close examination of miRNA gene targets reveals numerous transcripts with potential biologic significance in TBI. For example, vascular endothelial growth factor A (VEGFA) induces proliferation and migration of vascular endothelial cells and is essential for angiogenesis. 107 VEGFA is targeted by miR-29a-3p (microT-CDS = 0.989) which rises in the acute TBI response (when the brain might wish to block VEGFA protein expression and prevent further bleeding), but drops in the subacute/chronic periods (when allowing VEGFA expression might be beneficial). The reelin transcript (RELN) encodes a secretory matrix protein that is imperative for the cell-cell positioning involved in neuronal migration. 108 Expression of RELN is disrupted in temporal lobe epilepsy and major depressive disorder. 109 RELN is targeted by miR-27b-3p (microT-CDS = 0.999), which is elevated across CSF, blood, and saliva in TBI patients.24,38,39,93,95,98 In addition, miR-27b-3p levels have been shown to correlate with reaction time on ImPACT testing. 38 Putative miRNA-mRNA interactions provide a simplistic biologic construct for the potential function of miRNA expression following TBI. Additional work to elucidate the exact role of individual miRNAs in the CNS injury response will help guide the selection of ideal biomarker candidates in various clinical circumstances (eg, severe-versus-mild injury; prognostic-versus-diagnostic utility).
Future Directions: Providing Clarity and Clinical Applicability Through miRNA Research
Methodology
Pilot studies have now established a putative role for miRNAs in TBI pathophysiology.22,77 Moreover, it is clear that the mechanistic findings in animal models extend to human patients with TBI, 95 allowing measurement of brain-related miRNAs in peripheral biofluids.23,39,61,92 Studies of children and adults with severe TBI have identified the miRNAs that are altered in the CNS and deserve primary consideration as TBI biomarkers.37,93,95 The miRNAs identified in this review (that overlap across multiple studies, or demonstrate dynamic longitudinal expression patterns across multiple biofluids) may provide high yield targets for future hypothesis-driven investigations. In animal models, such investigations might manipulate miRNA expression to define cell- or tissue-specific actions, or explore TBI resilience, as Ge et al 77 have done with miR-21.
It will be critical that future investigations eschew the small sample sizes employed in pilot research. Hundreds of samples from TBI patients will likely be required to control for the inter-individual variation that exists in baseline miRNA expression 110 and produce reliable results. Alternatively, as LaRocca et al 99 recently demonstrated, pre-injury miRNA baseline levels can be employed, although this approach is costly and time-consuming. Starting from hypothesis-driven miRNA targets, future studies should consider building diagnostic/prognostic miRNA algorithms in a training cohort, then validating the findings in large, external hold-out sets. This rigorous approach was recently demonstrated with success by Di Pietro et al 38 in their study of salivary miRNA among concussed athletes. To ensure the reliability and reproducibility of such findings, investigators should provide explicit details for sample collection, sample storage, miRNA isolation, and miRNA quantification whenever possible. Selecting reagents and instruments from companies that have established miRNA protocols with test-retest reliability will allow others to reproduce findings and validate the clinical promise of miRNA biomarkers.111-113
Considering context-specific miRNAs
Severity of injury, timing of sample collection, biofluid selection, participant age, and medical comorbidities can all impact miRNA patterns following TBI. Careful consideration of these factors when designing future miRNA trials and interpreting results will be necessary to discern reliable biomarkers from statistical artifact. For example, a number of miRNAs demonstrate overlapping expression patterns in both severe and mTBI,23,37,92,94 but it is likely that distinct miRNA panels will be necessary to provide the clinical specificity and sensitivity required for these disparate conditions. After all, mTBI and severe TBI involve different pathophysiologic mechanisms and unique medical management considerations. Among patients with severe TBI, GCS and clinical endpoints have been used with some success to delineate clinical severity, whereas in mTBI GCS does not provide sufficient granularity, so employing sensitive measures of balance and cognition38,99 will provide objective measures of functional deficits while aligning with clinical guidelines for mTBI care.
Mounting evidence demonstrates that saliva miRNAs may provide a novel window into the brain,114-117 and that salivary miRNA levels may have utility in TBI.37,38,49,99 Although saliva and blood share some common miRNA biomarkers following TBI (Figures 1A and B), the 2 biofluids also demonstrate distinct signatures after injury. 99 This may result from the origin of miRNAs in these 2 fluids. Saliva, which can receive exosomal miRNAs directly from cranial nerves in the oropharynx, demonstrates a rapid miRNA response after TBI. In comparison, blood-based miRNAs must traverse the BBB, which may account for their relative delay in temporal patterns. 99 In the secondary response to TBI, it is likely that serum miRNAs carry information about the inflammatory mechanisms underlying neuronal injury (although this response may be muted without disruption of the BBB). Salivary miRNAs, on the other hand, appear to reflect the secondary neuroplasticity response. This may explain why saliva miRNA levels measured 1 week after injury predict prolonged concentration/memory difficulties with such accuracy. 49 Saliva collection allows immediate sideline assessment by coaches and athletic trainers without venipuncture training. 118 Non-invasive saliva collection also has the added benefit of room-temperature miRNA stabilization (when using RNA-specific devices).119,120 Ultimately, serum and saliva miRNA profiles are likely to provide complementary information following TBI. Investigations that harness both biofluids in parallel may help clarify the clinical role for each approach.
Clinical utility must also be considered when selecting time-points for miRNA collection. Numerous studies have demonstrated that CNS 37 and peripheral38,39,94,97-99 miRNA profiles fluctuate across the primary and secondary phases of TBI. The miRNAs responding within hours of injury are likely to be most suitable as a diagnostic tool. Studies with this clinical focus should employ large control groups, matched by age, sex, and ethnicity. For translation to a clinical setting, control groups must include individuals with non-cranial orthopedic injury,23,61 as well as post-exercise participants (for field-side applicability in concussed athletes).121,122 Conversely, miRNAs involved in the subacute response (days to weeks after injury) may provide more information for prognosis and clinical outcomes. Investigations geared toward prognostic biomarkers should consider timing of typical follow-up visits in outpatient TBI management (7-14 days) and use controls whose medical comorbidities may mirror prolonged TBI symptoms (eg, patients with depression, attention deficits, or disordered sleep). 96 Finally, with the rising concern over traumatic encephalopathy and the long-term implication of TBI, measuring miRNA levels at chronic time-points alongside sensitive functional outcomes (eg, memory, balance) and imaging may help fill critical gaps in the current miRNA TBI literature (see gray boxes in Figure 1).
Clinical applications
The primary emphasis of most miRNA studies has been TBI diagnosis. Certainly, an objective marker of mTBI would provide clinical benefit in emergency department and outpatient settings. Currently, clinicians in these settings rely on subjective symptom reports because neurologic examination and imaging are typically normal. Recent epidemiologic research suggests that health care providers may have previously under-diagnosed mTBI. 123 As such, addition of an objective diagnostic biomarker could improve mTBI recognition. In cases of negative testing, it would also provide valuable reassurance to patients and families that head trauma did not result in biologically detectable brain injury.
This sort of reassurance will require development of markers sensitive enough to detect even subconcussive hits to the head. It will also require that we understand how such blows impact long-term brain health. The interplay between repeated minor head trauma and chronic traumatic encephalopathy necessitates examining athletes in contact sports to identify miRNAs that change from minor head impacts. The study by LaRocca and colleagues begins this critical work, by identifying saliva and blood-based miRNA signatures in mixed martial arts fighters with subconcussive hits to the head. By treating the miRNA response as a function of the number of blows to the head, and not an “all-or-none” concussion experience, the study defines miRNAs that portend minor brain disruption.
What happens after TBI diagnosis? Surveys of health care providers demonstrate a critical need for objective toolsets that guide prognosis and clinical management.4,124,125 This need extends from mTBI through severe TBI. Clinical risk scores using information about injury mechanisms and patient factors are currently used to predict outcomes,126,127 but these tools can be time-consuming and inaccurate. In intensive care settings, separate sets of biomarkers will likely be required to predict outcomes involving morbidity and mortality. It would be naïve to expect such biomarkers to completely overlap with those used for mTBI diagnosis.
In mTBI, there is a unique opportunity to harness miRNA profiles to divide patients into phenotypic subgroups and guide individualized treatment plans. For example, recent findings that salivary miRNAs identify children at risk for PCS and that specific miRNAs may be associated with individual symptoms could be used for initiation of early individualized therapy. 49 In patients with miRNA profiles that portend PCS characterized by chronic headaches, this might mean early initiation of aggressive triptan therapy. In patients with miRNA levels implicated in prolonged concentration difficulties, this might inform individualized return-to-learn decisions or trigger prescription of stimulant medication.
Accumulating research demonstrates that graded exercise routines can accelerate TBI recovery in a subset of patients identified through treadmill testing.128,129 Interestingly, some of the same miRNAs that demonstrate alterations in patients with TBI show contrary patterns with endurance exercise. 121 This concept has also been validated in mouse studies, where voluntary running mitigates the miRNA response to TBI, 130 and post-injury running triggers miR-21 alterations associated with cognitive improvements. 131 In this context, miRNAs represent a means for phenotypic assessment, as well as potential therapeutic targets in and of themselves.
Conclusions
Despite extensive research involving TBI biomarkers, significant gaps remain. There are limited tools for diagnosis, prognosis, and therapeutic decisions after TBI. Emerging research suggests that miRNAs are ideally suited to fill this need. miRNAs are highly expressed in the brain, can cross the BBB, and are stable in peripheral biofluids. Moreover, the unique function of miRNAs in neuronal communication may provide a distinct window into the injured brain. The miRNA response to TBI is complex, and at times messy. However, with careful consideration of the physiologic factors involved, it is possible to discern commonalities across studies. Focusing on the miRNA candidates with reproducible findings across multiple biofluids and time-points may yield clinical utility in the near future. It will be critical that future miRNA investigations employ large sample sizes, while considering confounding factors such as patient ages, or injury severity. If these factors are addressed, miRNAs may one day guide diagnostic, prognostic, and therapeutic decision making in patients with TBI.
Supplemental Material
Supplemental_Figure_1A – Supplemental material for A Review of MicroRNA Biomarkers in Traumatic Brain Injury
Supplemental material, Supplemental_Figure_1A for A Review of MicroRNA Biomarkers in Traumatic Brain Injury by Hamna Atif and Steven D Hicks in Journal of Experimental Neuroscience
Supplemental Material
Supplemental_Figure_1B – Supplemental material for A Review of MicroRNA Biomarkers in Traumatic Brain Injury
Supplemental material, Supplemental_Figure_1B for A Review of MicroRNA Biomarkers in Traumatic Brain Injury by Hamna Atif and Steven D Hicks in Journal of Experimental Neuroscience
Supplemental Material
Supplemental_Figure_1C – Supplemental material for A Review of MicroRNA Biomarkers in Traumatic Brain Injury
Supplemental material, Supplemental_Figure_1C for A Review of MicroRNA Biomarkers in Traumatic Brain Injury by Hamna Atif and Steven D Hicks in Journal of Experimental Neuroscience
Footnotes
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests:
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: SDH is a paid consultant for Quadrant Biosciences Inc. He is named as an inventor on intellectual property using miRNA technology in patients with traumatic brain injury. AF has no conflicts of interest to disclose.
Author Contributions
SDH conceived of the study. AH and SDH contributed equally to the literature review and drafting the manuscript.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.