
Abstract
The use of arbuscular mycorrhizal fungi (AMF) to enhance soil phosphate uptake is a biological technique considered to cure phosphate deficiency in soils. This study investigated association of Aspilia pruliseta Schweif shrub with AMF in Kenya. The study aims at profiling a tropical shrub with multiple ecological benefits that could reduce addition of chemical phosphatic fertilizer into the soil and reverse negative consequences of eutrophication. Sampling was purposive to have areas with or without Aspilia pruliseta vegetation growing. A small amount (10 g) of the soil from 27 composite samples was used for spore count determination and a similar amount for next generation sequencing. Spore counts varied significantly among soil textural types, sample locations, and soil depth. Sandy loam had the highest spore counts with a mean average of 404 spores. The spore count decreased significantly (P < .05) with the depth of soil from a mean of 514 spores to 185 along the rhizosphere. The intensity of spore morphotypes was significantly higher at P < .05 for soils whose vegetation was covered with Aspilia pruliseta than those without. Aspilia pruliseta vegetation used together with sandy loam soil could culture commercial mycorrhiza fungi production for use in agrisystems.
Introduction
Global reserves of phosphorus are running out fast.1-3 This poses a great challenge for global food production in the foreseeable future because phosphate is one of the macronutrients needed for plant growth.4,5 Phosphorus (P) deficiency occurs in many soils of East Africa not only due to P depletion through crop harvest and erosion but also due to P-fixing soils in the region. 6 A natural method of solubilizing fixed phosphorus to make it available to plants alleviates this problem. 7 Replenishment of phosphate in the soil is mainly dependent on chemical fertilizer.8-12 In Kenya, the most common phosphate-enhancing fertilizers added include diammonium phosphate, single superphosphate, and double superphosphate. This is a challenge to small-scale farmers who cannot afford frequent replenishment of phosphatic fertilizer due to their low income. Soils in the central highlands region of Kenya are among the affected ones13,14 and the lower parts of Embu and Tharaka-Nithi counties though arid and semiarid are located within this P-deficient zone. Soil degradation caused by the usual farming of sorghum, millets, cowpeas, and green grammes in these fragile ecosystems pose an added threat to agricultural production15,16 and therefore much attention must be paid to restoring and maintaining soil health, particularly in arid and semiarid environments. A suitable cover crop, with multiple benefits17-20 growing during the dry season would be ideal for these environments. Phosphorus is a macronutrient needed by crops for growth. It makes up about 0.2% of dry weight in terrestrial plants, but it is one of the most difficult nutrients for plants to acquire.21-23 In soil, P occurs mainly as insoluble phosphates and is not readily available. 24 However, with the help of micro-organisms, this insoluble phosphate is solubilized to become available to plants. 25
Arbuscular mycorrhizal fungi (AMF) can provide an effective pathway by which phosphorus is scavenged and rapidly delivered to cortical cells within the root.26,27 Arbuscular mycorrhizal fungi enhance P utilization in soils that makes it available to plants.28-30 In this work, we sought to evaluate the intensity of AMF in the rhizosphere of Aspilia pruliseta Schweif in the different depths of sandy loam, silt loam, and silty clay soils and profile the plant as both mycorrhizing as well as a cover crop in arid and semiarid environments. Aspilia pruliseta is a flowering plant in the Asteraceae family and was hypothesized to grow in coexistence with the mycorrhiza fungi. It is hypothesized that this complex association leads to availability of phosphorus in the soils in usable forms to plants. The shrub is common and grows naturally in the open woodlands and grasslands in western, southern, central, and eastern Africa. Farmers in central eastern Kenya have reported good crop yields, particularly cereals, in farms previously growing Aspilia pruliseta. When uprooted, Aspilia pruliseta (Figure 1A) presents huge surface area of white mycelium and a robust rooting system that is visible without any magnification (Figure 1B). There seems to be a relationship between the good crop yields and the mycelium on the roots of Aspilia pruliseta. Formation of “dense” cluster roots that produce organic anions are some of the strategies that some plants use to enhance P availability or uptake,27,31,32 a phenomenon that seem to be exhibited by Aspilia pruliseta. Availability and uptake of P from natural and fertilizer sources remains the subject of active research. 33 This research will investigate the role of mycorrhiza fungi in mediating P availability within the rhizosphere of Aspilia pruliseta and its availability for utility by other follower plants. The work reported here tested on the superiority of Aspilia pruliseta to aggregate AMF spores that could be useful to subsequent cultivated crops during the wet season.

(A) Aspilia pruliseta Schweif. shrub growing in the wild in Tunyai, Kenya, during the wet season (Photo taken by James Muchoka on November 10, 2017) and (B) roots of a newly uprooted Aspilia pruliseta exposing the white mycelia (photo taken by James Muchoka on November 10, 2017).
Materials and Methods
Study area
This study was carried out in Embu and Tharaka-Nithi counties of eastern Kenya with a semiarid climatic set up. The 3 study sites were situated at co-ordinates 00°21′00″ S, 37°28′30″ E (Figure 2A), 00°10′00″ S, 37°50′00″ E (Figure 2B) and 00°12′00″ S, 37°51′00″ E (Figure 2C). Elevation for all the studied sites was below 1000 m above sea level. Rainfall in the selected areas was below 700 mm/year with 2 distinct but unreliable wet seasons in the months of March to May and October to December. Dry spells were more prolonged with temperature mean of 27°C. Scanty natural tree and shrub vegetation found was mainly Aspilia pruliseta, Cassia sp, Euphorbia sp, Acasia sp, Balanites aegyptiaca. Cenchrus ciliaris and Hyperhenia rufa grasses were interspersed in trees and shrubs. The selected sites had silty clay, silt loam, and sandy loam soil textural types and all of them had Aspilia pruliseta growing in the natural environment (Figure 2D) for the period of study.

(A) Collection of soil samples at Kanyuambora, 00°21′00″ S, 37°28′30″ E (photo by James Muchoka, September 12, 2017), (B) soil samples collection at Tunyai site, 00°10′00″ S, 37°50′00″ E (photo by James Muchoka, September 12, 2017), (C) soil samples collection at Gakurungu, 00°12′00″ S, 37°51′00″ E (photo by James Muchoka, September 13, 2017), and (D) silt loam soil sampling along the rhizosphere of Aspilia pruliseta in Tunyai site (photo by James Muchoka, September 28, 2017).
Soil AMF spores extraction, isolation, and counting method
Soils were sampled from Gakũrũngũ, Tunyai, and Kanyuambora field sites. A reconnaissance survey was initially carried out in which natural seed-bearing Aspilia pruliseta plots were mapped out for each of the 3 sites. Each of the site measured 4 km2 and was an administrative subunit. In each site, 3 subsites measuring 100 m by 50 m were identified which formed the plots. Soil was sampled in the rhizosphere of Aspilia pruliseta vegetation and from patches without as a control. Soils from these subsites were combined to form one composite mixture for each depth sampled (depth1, 0-20 cm; depth2, 21-40 cm, depth3, 41-60 cm).
Soil sampling was purposive for each of the selected sites based on the soil textural types in the area: sandy loam, silty clay, and silt loam soils. To qualify for sampling, the specific soil textural types had to have at least 70% of the soil particle under consideration based on soil categorization through use of soil sieves with known particle diameter.34,35 A tape measure was used to delineate the clearance area of 1 m2 of the selected natural site where mature test plants were located. The underneath vegetation was cleared using a panga, and a standard soil auger (SOD-GP Dormer sampling equipment) was used to collect soil samples taken from the test plant rhizosphere at depths 0 to 60 cm. The patches without the test plant were either bare or covered with Hypharhenia rufa grass. One-metre length tape measure was used to take vertical measurements (in centimetres) along the profile. Three samples for each parameter of interest was taken, each with a volume of 40 cm3 (the volume the soil auger can scoop) for the various depths (0-20 cm, 21-40 cm, and 41-60 cm) in different subsites and were mixed together to form a single sample. Three soil samples from each subsite (plots) and for every soil depth under consideration were taken, giving the number of soil samples taken as 27 in every soil textural group for the plots that the Aspilia pruliseta grew. Similar procedure was followed for the sampling plots without this vegetation type. Overall number of soil samples taken was 486. Samples were carefully labelled and put into separate sterilized hessian bags before taking them to the laboratory for analysis. This soil was then thoroughly mixed but maintaining the site, subsite, and soil depth of the collected samples. About 100 g of the mixture was put into khaki paper bags for laboratory analysis.
Laboratory analysis
A small amount (10 g) of the composite soil sample from each of the designated soil depth, soil textural class and in the 3 field sites were weighed using an electronic balance and soaked in 1 L of water before washing it several times by passing it through a 710 µ sieve placed on top of a 45 µ sieve. 36 The 710 µ sieve collected the roots and course debris while the 45 µ prevented the spores from passing through. The sieved water and soil mixture was then collected in a 1-L cylinder. The washing and decanting process was done several times until near-clear water was obtained. This was followed by filling the centrifuge tubes with the sieved content. Centrifugation was done for 5 min at 1500 r/min, and the filtrate was poured off while the supernatant remained at the bottom of the tube. About 48% sucrose solution was added to the supernatant at equal volumes (50 mL) and centrifuged for 1 min at 1500 r/min. The filtrate was collected on the 45 µ sieve while the supernatant was disposed off. 36 The filtrate was then washed with slowly flowing tap water to wash off the sucrose. The washed content was then collected in a 50-mL plastic cylinder. To collect and separate the spores of different morphotypes, a Petri dish with grid lines was used to hold the water content after the centrifugation process. Using a dissecting microscope to view the contents in the Petriplate, a fine forceps was used to collect the spores. The available spores were collected, separated, and placed in a small round watch glass. 36 The Edinburgh Botanic Gardens colour chart for fungi was used in the determination of spore colour. The mycorrhizal spores were then separated according to the size, shape, colour, hyphal attachments, and surface appearance. Based on these characteristics, the spores were quantified, mounted separately on glass slides using polyvinyl lactoglycerol (PVLG) and Meltzer reagent. The spores mounted on the PVLG were mounted whole and the ones on Meltzer reagent were slightly crushed to reveal finer details. 36
This process was repeated in the adjacent site in soils where Aspilia pruliseta (test plants) were not growing but other test variables remained the same.
Root colonization by mycorrhiza fungi on Aspila pruliseta
Roots of Aspilia pruliseta were cleared in 25% KOH and autoclaved for 15 min. 37 After cooling, the roots were washed in tap water and then bleached in ammonia, hydrogen peroxide and water solution for 2 h. After bleaching the roots, they were rinsed in water before adding 1% hydrochloric acid. The acidified roots were then stained using 0.05% acidified glycerol stained with trypan blue and autoclaved for 3 min. The roots were then rinsed in water and stored temporarily in acidified glycerol without the stain. A minimum of 30 × 1 cm roots were chosen randomly and 15 fragments placed parallel to each other on the slide. Polyvinlylactoglycerol (PVLG) was used as a mounting reagent and the slide was covered using a 24 × 50 mm coverslip. The roots were squashed gently to reveal the AMF infection. The roots were assessed under the compound microscope using a linear eyepiece moving along each root fragment. The roots were rated according to the ranges of classes by Trouvelot et al. 37 The classes gave a rapid estimation of the abundance of arbuscules and the level of mycorrhizal colonization of each root fragment.
Soil and rootlets total DNA extraction
A small amount of (10 g) of composite soil from the rhizosphere of Aspilia pruliseta comprising of the plant’s rootlets was weighed using an electronic balance. The weighed soil was put into 100 mL beaker and about 50 mL tap water was added. The mixture was placed on an electronic stirrer overnight. After 12 h, the mixture was washed several times by passing it through a 710 µ sieve placed on top of a 45 µ sieve. 36 The 710 µ sieve collected the roots and course debris while the 45 µ prevented the spores from passing through. The roots and course debris from the 710 µ sieve were put into a mortar and air-dried in a hood while the process of sieving continued by collecting sieved water and soil mixture on a 1-L cylinder. 36 The washing and decanting process was done several times until near-clear water was obtained. This was followed by filling the centrifuge tubes with the sieved content. Centrifugation was done for 5 min at 1500 r/min and the filtrate was poured off while the supernatant remained at the bottom of the tube. A 48% sucrose solution was added to the supernatant at equal volumes (50 mL) and centrifuged for 1 min at 1500 (r/min). 36 The filtrate was collected on the 45 µ sieve while the supernatant was disposed off. The filtrate was then washed with slowly flowing tap water to wash off the sucrose. The washed content was then collected in a 50 mL plastic cylinder and the contents poured into a filter paper. Using a fine pair of forceps, the contents were picked and transferred to eppendorf tubes. The dried plant roots in the mortar were crushed into a fine powder using a pestle and the contents added to fungal spore cells in the eppendorf. 38 The content in the eppendorf was resuspended in 100 µL of solution A {100 mM Tris-HCL (pH 8.0), 100 Mm EDTA (pH 8.0); added to 5 µL of lysozyme (from a 20 mg/mL solution) and incubated at 37°C for 30 min in a water bath}. A small volume (400 µL) of lysis buffer (solution B) comprising 400 mM Tris-HCL (pH 8.0), 60 mM EDTA (pH 8.0), 150 mM NaCl, 1% sodium dodycyl sulphate and the tube was left at room temperature for 10 min. A small volume (10 µL) of Proteinase K (20 mg/mL) was mixed gently and incubated at 65°C for 1 h in a water bath. 39 An equal volume of chloroform/isoamyl alcohol was added and centrifuged at 13 200 r/min for 5 min at 4°C. The supernatant was transferred to new tubes. In the new tubes, 150 µL of sodium acetate (pH 5.2) and an equal volume of isopropanol alcohol were added accordingly. The tubes were briefly mixed through inversion. The mixture was then incubated at −20°C overnight. The tubes were then spun at 13 200 r/min for 30 min and the supernatant was discarded. The resultant DNA pellets were washed in 300 µL of 70% ethanol. The pellets were then spun at 10 000 r/min for 1 min, and the supernatant was discarded. The resultant DNA pellet was air dried in the hood and dissolved in 50 mL of 13 Tris-EDTA. Genomic DNA (5-15 ng) in 10 µL of ddH2O was used for RAPD amplification using 1.5% agarose gels and images obtained confirming the presence of DNA. 38 Six samples (3 samples each from depth 1-3 in soils from Aspilia pruliseta vegetated soils and 3 from adjacent soils without the vegetation) were dried using LABCONCO machine. About 30 µL of the confirmed DNA was shipped to mrdnalabs (USA) for next-generation sequencing with the primers as diversity assay bTEFAP average inhouse ITSwanda. Illumina was used as the sequencing technology method.
Data analysis
Data obtained from the factors that were tested was subjected to 3-way analysis of variance (ANOVA) using SAS Edition 9.2 and differences between treatments mean was examined using least significant difference (LSD) at P < .05. The spore counts data from the soil textural types, rhizosphere depths, and locations with Aspilia pruliseta were compared to their controls. Arbuscular mycorrhiza fungi root colonization was determined to assess the mycorrhizal infectivity potential of the shrub. Mycorrhiza spores were identified using total genomic sequencing method to give an indication of the type and numbers in the shrubs’ rhizosphere.
Results and Discussions
The 3 tested soil textural types (silty clay, silt loam, and sandy loam) promoted proliferation of mycorrhizal fungal spores differently as evidenced by spore counts (Table 1). The highest spore count was in sandy loam soils, and the lowest was in silty clay soils for all sites studied (Figure 3, mean of 404.2 and 260.7 respectively). Spore counts for silty clay soils were statistically lower than those of sandy loam and silt loam soil in all the studied sites (Figure 4), with 224.5 spores for silty clay soil at Gakurungu compared to 299 and 394 mycorrhiza fungal spores for silt loam and sandy loam soils, respectively, for the same site. Similar observation was noted for Tunyai and Kanyuambora site. In all the soil textural types, mycorrhiza spores were recorded, albeit in differing numbers, confirming ubiquitousness of mycorrhiza fungi microflora. 39 Soil textural types seemed to play a significant role in determining the number of mycorrhiza spores in the rhizosphere. Other scholars40-44 corroborate this finding on the influence of soil type on mycorrhizal populations. The difference in spore counts in sandy loam and silt loam soils were not statistically significant but intensity levels in silty clay soils were significantly lower at P ⩽ .05 compared with the other type of soils. Past research asserts that this variance between soil textural types influencing AMF communities is still a subject of study. However, the study by Rodríguez-Caballero et al, 45 del Mar Alguacil et al 46 concluded that soil pH and levels of 2 micronutrients (Mn and Zn), play significant roles in triggering AMF populations.
Arbuscular mycorrhiza fungal spores count for the studied factors at different soil sampling sites (different superscript letters denote significant differences; LSD test, P = .05 between groups).

Abbreviation: LSD, least significant difference.
LSD is the smallest significance difference between 2 calculated means at P < .05.
‘a’, ‘b’, and ‘c’ values having the same letter under each factor are not significantly different at P < .05.

Arbuscular mycorrhiza fungi average counts for soil samples in sampling sites. Error bars denote SE, P < .05.
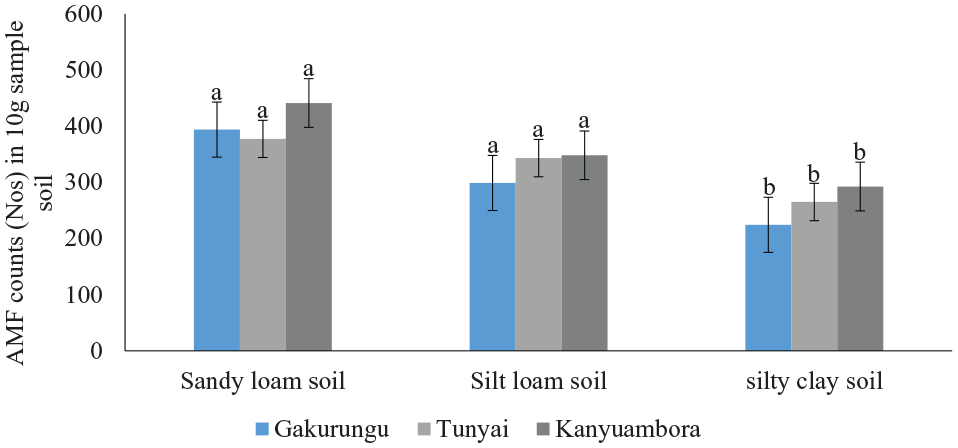
Arbuscular mycorrhiza fungi count variation for different soil textural types in 10 g of sampled soils. Error bars denote SE, P < .05.
Soils inhabited by Aspilia pruliseta in all the 3 sites showed a higher number of morphotypes compared to those without this plant species (Table 1). Plant, soil, and climatic factors determine the development of mycorrhiza fungi and influence establishment of the mycorrhizal symbiosis and its efficiency.47-49 The ability to produce mycorrhizal propagules (spores, hyphae, and colonized roots) is enhanced through the inoculation of plants with AMF. This is an important part of the process of soil microbiota recuperation.30,50 Although different factors were considered in determining AMF spore production, different plant species were tested in different locations, giving different results from this current experiment.
Furthermore, the experiment demonstrated that mycorrhiza spore count in the rhizosphere of Aspilia pruliseta decreased with increase in soil depth (Figure 5). The intensity of mycorrhiza varied significantly with soil depth ranging from 514 to 185 counts for 0 to 20 cm and 41 to 60 cm depths, respectively (Table 1 and Figure 5). This inverse relationship on mycorrhiza fungi population and soil depth was exhibited in all the sites tested (Figure 5). On comparison of the 3 sites, AMF population was the highest in Kanyuambora and the lowest in Gakurungu (Figure 6). It is not rather obvious for mycorrhiza population to decrease with an increase in soil depth as suggested51-53 because studies by Gucwa-Przepióra et al 54 got different results. Gucwa-Przepióra et al 54 found out that there was an increase in arbuscular mycorrhiza colonization with increasing soil depth because of chemophytostabilization amendments. An increase in AM colonization with increasing soil depth was observed in soils with spontaneously growing Deschampsia cespitosa. Similar work by Mejstrik, 51 but using different plant species indicated that the frequency and intensity of vesicular-arbuscular mycorrhiza in Molinietum coeruleae was soil depth dependent and that the higher the depth, the less the intensity of AMF. Rillig and Field, 52 in their research report indicated that arbuscular mycorrhiza responds to plants exposed to elevated atmospheric carbon dioxide and that this gas is a function of soil depth. In addition, Asghari et al, 53 while on the study of growth response of Atriplex nummularia to inoculation with AMF at different salinity levels showed higher infectivity rate with decreased salinity levels which in itself, was negatively correlated to soil depth. Curiosity of results from these scholars drove us to study more on depth aspects for Aspilia plant. Indeed this study agrees with similar studies by Shukla et al, 55 on soil depth as an overriding factor for AMF. In other studies, although not related to Aspilia pruliseta, mycorrhiza fungi populations have been shown to decrease with an increase in soil depth.56-58 Some researchers suggested that it might be due to the less organic content 57 and low availability of oxygen in deeper soil zones 59 because fungi are sensitive to low oxygen pressure which prevails at lower depths. 60 According to Anderson et al 61 and Bever et al, 62 AMF are generally scarce where the plant roots are sparse. Besides, this phenomena 63 conclude that strong mycorrhizal plants will always have more rhizosphere spore counts compared to nonmycorrhizing plants. Based on these earlier findings, Aspilia pruliseta shrub satisfies the criteria of a strong mycorrhizal plant.
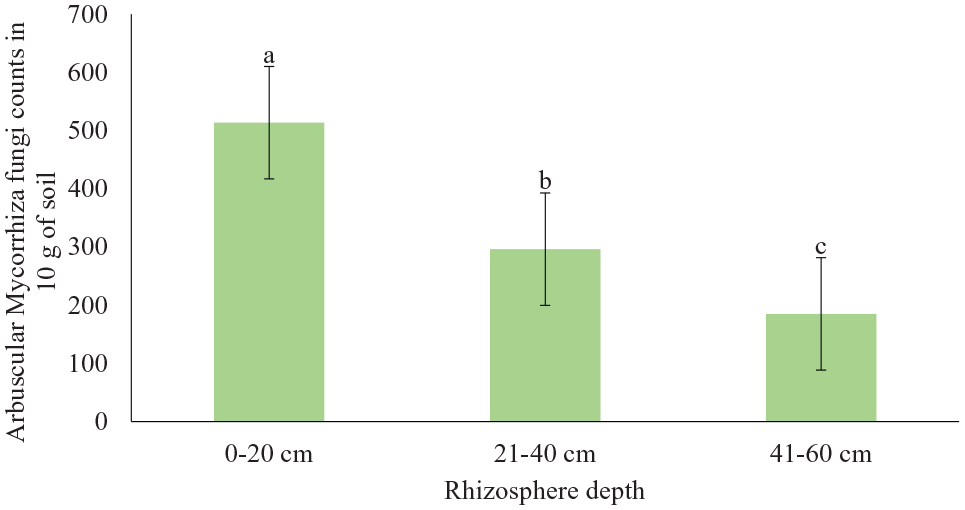
Arbuscular mycorrhiza fungi average counts in the rhizosphere of Aspilia pruliseta in different soil depth. Error bars denote SE, P < .05.

Site effect on Arbuscular mycorrhiza fungi counts along the rhizosphere of Aspilia pruliseta. Error bars denote SE, P < .05.
Higher spore counts were observed where Aspilia pruliseta vegetation was growing compared to soils that did not have this plant species. Spore counts in 10 g of soil was 420.6 spores, 381.3 and 417.8 in Gakurungu, Tunyai and Kanyuambora, respectively, in sample sites with Aspilia pruliseta compared to 191.1 spores, 275.8 and 303.8 spores for the same sites where there was no Aspilia pruliseta plant species growing (Figure 7). Mycorrhizal root colonization percentage in Aspilia pruliseta shrub was 96.7 (Table 2). Molecular work (described above on DNA extraction) identified 35 different mycorrhizal fungal species in the rhizosphere where Aspilia pruliseta vegetation grew compared to adjacent soils without this plant species (Table 3). Species composition and diversity varied along the rhizosphere with more abundance in depth 1 (0-20 cm). The lower rhizosphere zone (depth 3, 41-60 cm) was dominated by Archaeospora trappei mycorrhizal fungi. From this experiment, Glomus sp was the most abundant in the rhizosphere of Aspilia pruliseta shrub. Mycorrhizal fungi species in the rhizosphere of Aspilia pruliseta shrub was important to this experiment as both their abundance and diversity would enhance the shrub’s potent in soil phosphate uptake by follower crop. Studies by Radhika and Rodrigues, 64 Torrecillas et al, 65 concluded that relative diversity of rhizosphere mycorrhiza fungi depended on host plant species. Although these case studies mentioned did not evaluate Aspilia pruliseta shrub, it is evidently clear that this shrub is host to various mycorrhizal fungi of importance to ecological reconstruction.

Arbuscular mycorrhiza fungi counts in sites with and without Aspilia pruliseta. Error bars denote SE, P < .05.
Aspilia pruliseta mycorrhiza fungi root colonization.

Abbreviations: a%, arbuscule abundance in mycorrhizal parts of root fragments; A%, arbuscules abundance in the root system; F%, the frequency of mycorrhiza in the root system; m%, the intensity of AMF colonization in the root fragments; M%, the intensity of the mycorrhizal colonization in the root system.
F% = 96.67, M% = 42.17, m% = 43.62, a% = 56.13, A% = 23.67.
Mycorrhiza fungal species and populations in the rhizosphere of Aspilia pruliseta, next-generation sequencing from soil and rootlets samples, species count; d1 (0-20 cm); d2 (21-40 cm); d3 (41-60 cm); Ap-Aspilia pruliseta.

Conclusions
The presence of Aspilia pruliseta shrub increased the level of colonization by AMF in soils. Since AMF is associated with phosphorus availability, Aspilia pruliseta could be grown and used as a soil fertility-enhancing shrub and for improved fallow systems. Sandy loam soils support AMF colonization better than silt loam and silty clay soils, thus, Aspilia pruliseta could be used to enhance phosphate levels in sandy loam soils hence improving productivity in the semiarid areas. It is difficult to culture obligate soil microbes for commercial exploitation. Mycorrhiza fungi are obligate saprophytes and this study has demonstrated that Aspilia pruliseta vegetation used together with sandy loam soil could culture commercial mycorrhiza fungi production for use in agrisystems.
Footnotes
Acknowledgements
The authors wish to thank the teams from the University of Embu and Kenyatta University in Kenya for the close collaboration in carrying out this research work. The team also wishes to thank the national research fund (NRF), a kenya government kitty for providing the initial funds to carry out this research.
Funding:
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
All authors to this research article contributed equally