
Abstract
Nitric oxide (NO) is a ubiquitous, endogenously produced, water-soluble signaling molecule playing critical roles in physiological processes. Nitric oxide plays pleiotropic roles in cancer and, depending on its local concentration, may lead to either tumor progression or tumor suppression. Addition of NO group to a cysteine residue within a protein, termed as S-nitrosylation, plays diverse regulatory roles and affects processes such as metabolism, apoptosis, protein phosphorylation, and regulation of transcription factors. The process of S-nitrosylation has been associated with development of different cancers, including breast cancer. The present review discusses different mechanisms through which NO acts, with special emphasis on breast cancers, and provides detailed insights into reactive nitrogen species, posttranslational modifications of proteins mediated by NO, dual nature of NO in cancers, and the implications of S-nitrosylation in cancers. Our review will generate interest in exploring molecular regulation by NO in different cancers and will have significant therapeutic implications in the management and treatment of breast cancer.
Keywords
Introduction
Breast cancer, although termed as one disease, is one of the most diverse cancers displaying heterogeneous expression of progesterone receptor (PR), estrogen receptor (ER), and human epidermal growth factor receptor 2 (HER2) in patients with different breast cancers, which also give rise to intertumor and intratumor heterogeneity and often lead to different clinical presentations in terms of histology, prognosis, and responsiveness to treatment. 1 One of the main reasons behind the broad heterogeneity is variation in transcriptional programs which could help in providing a distinct molecular profile for each tumor. 2 Early onset of breast cancer with an aggressive phenotype has been observed in young women, making breast cancer awareness and screening among younger women a priority. 3 Research performed in the last few decades has highlighted the role of genomic alteration in driving breast cancers and has drawn special attention to role of genomic drivers, impact of DNA repair defects, and resistant clones in the disease. 4 One of the reasons behind DNA damage and impaired DNA repair is oxidative stress. Increasing evidence suggests that a situation of oxidative stress arises when an imbalance in the rate of generation and disposal of reactive oxygen species (ROS) or reactive nitrogen species (RNS) occurs. The generated reactive species are capable of playing a dual role and can cause oxidative damage as well as act as molecular signals and activate stress responses, beneficial for the organism. 5 Reactive nitrogen species play a crucial role in physiological regulation in cells by displaying pleiotropic effects on cellular targets. An elevated level of RNS induces nitrosative stress and has been implicated in cell injury and death. 6
The development of cancers including breast cancer is affected by various intracellular and extracellular factors, including reactive oxygen and nitrogen species (RONS). The cellular accumulation of RNS is implicated to play a role in cancer initiation and progression by causing alterations in gene expression profile, deregulating signal transduction pathways, and inducing abnormal protein modifications. 7 As cancer cells are adapted to grow in low oxygen concentration or hypoxia, they undergo metabolic reprogramming to meet their elevated demands of nutrients and energy for proliferation and survival. Metabolic adaptations of an increased rate of anaerobic glycolysis, decreased oxidative phosphorylation generation, and overall mitochondrial dysfunction govern cancer growth. 8 As reported in different cancers, hypoxic condition results in an increased production of RONS. 9 Cancer cells with chronic inflammatory conditions and elevated concentration of reactive species face oxidative (imbalance between generation of ROS and antioxidants) and nitrosative stress (imbalance between production and elimination of RNS), leading to DNA damage and impaired DNA repair. 10 Elevated levels of RONS have high reactivity and, hence under normal physiologic conditions, require proper redox balance between RONS generation and elimination by the internal antioxidant system. 11 The clean-up of elevated intracellular RONS concentration controls the reversible modifications of regulatory proteins caused by oxidative/nitrosative stress and acts as redox switches controlling activities of intracellular downstream effectors of different cell signaling pathways via ROS/RNS signaling. 12 Reactive nitrogen species have been implicated in disturbed cellular homeostasis leading to deleterious consequences by deregulation of signaling pathways causing protein modifications. 13
Nitric Oxide, Nitric Oxide Synthase, and RNS
Nitric oxide (NO) is a highly diffusible and reactive diatomic free radical. Present in gaseous state at room temperature, NO has pleiotropic functions and plays a critical role in multiple biological processes, such as neurotransmission, vasodilatation, and macrophage-mediated immunity. In addition, NO can also act a messenger molecule and play a role in promoting as well as inhibiting cancer. 14
Nitric oxide synthase (NOS; EC 1.14.13.39) enzyme helps in the synthesis of NO from
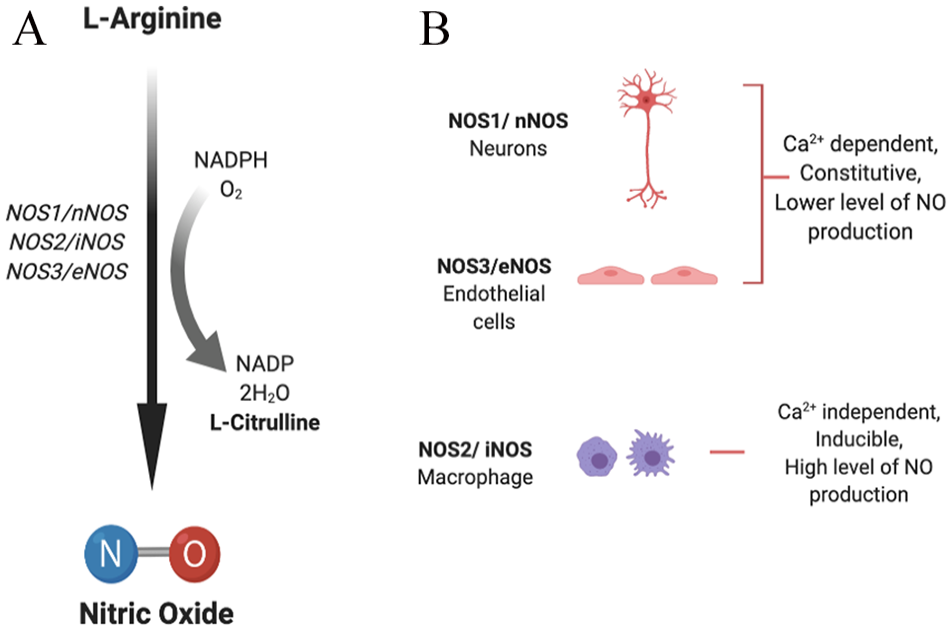
(A) Synthesis of nitric oxide. (B) Nitric oxide synthase (NOS) isoforms: neuronal (NOS1/nNOS), inducible (NOS2/iNOS), and endothelial (NOS3/eNOS) isoforms catalyze the process of NO generation in the presence of cofactors via oxidation of
NOS1 and NOS3 are dependent on calcium levels for activity and hence produce lower but transient concentrations of NO. They are continuously expressed in neurons and endothelial cells, whereas NOS2 is a calcium-independent, inducible isoform and, once induced, results in continuous production of higher concentrations of NO. 13 The expression of nNOS and eNOS can also be activated or inhibited via different protein kinase–mediated phosphorylations, while expression of iNOS can be regulated transcriptionally by multiple factors, including cytokines (interleukin [IL]-1β, interferon [IFN]-γ, tumor necrosis factor [TNF]-α), bacterial endotoxin (lipopolysaccharide [LPS]), and oxidative stress. 14
Nitric oxide is the primary, common progenitor for all RNSs, and different RNSs are formed by NO-dependent reactions. Peroxynitrite (ONOOH−) is formed by a fast reaction between NO and O−2, which forms secondary RNS by further reaction. Nitric oxide reacts with its intracellular environment to form other reactive metabolites including peroxynitrite, nitrite, nitrate, or S-nitroso-thiols that induce genotoxic effects leading to DNA damage. Peroxynitrite species are capable of causing single-strand DNA breaks by attacking the sugar-phosphate backbone of DNA (Figure 2). 14
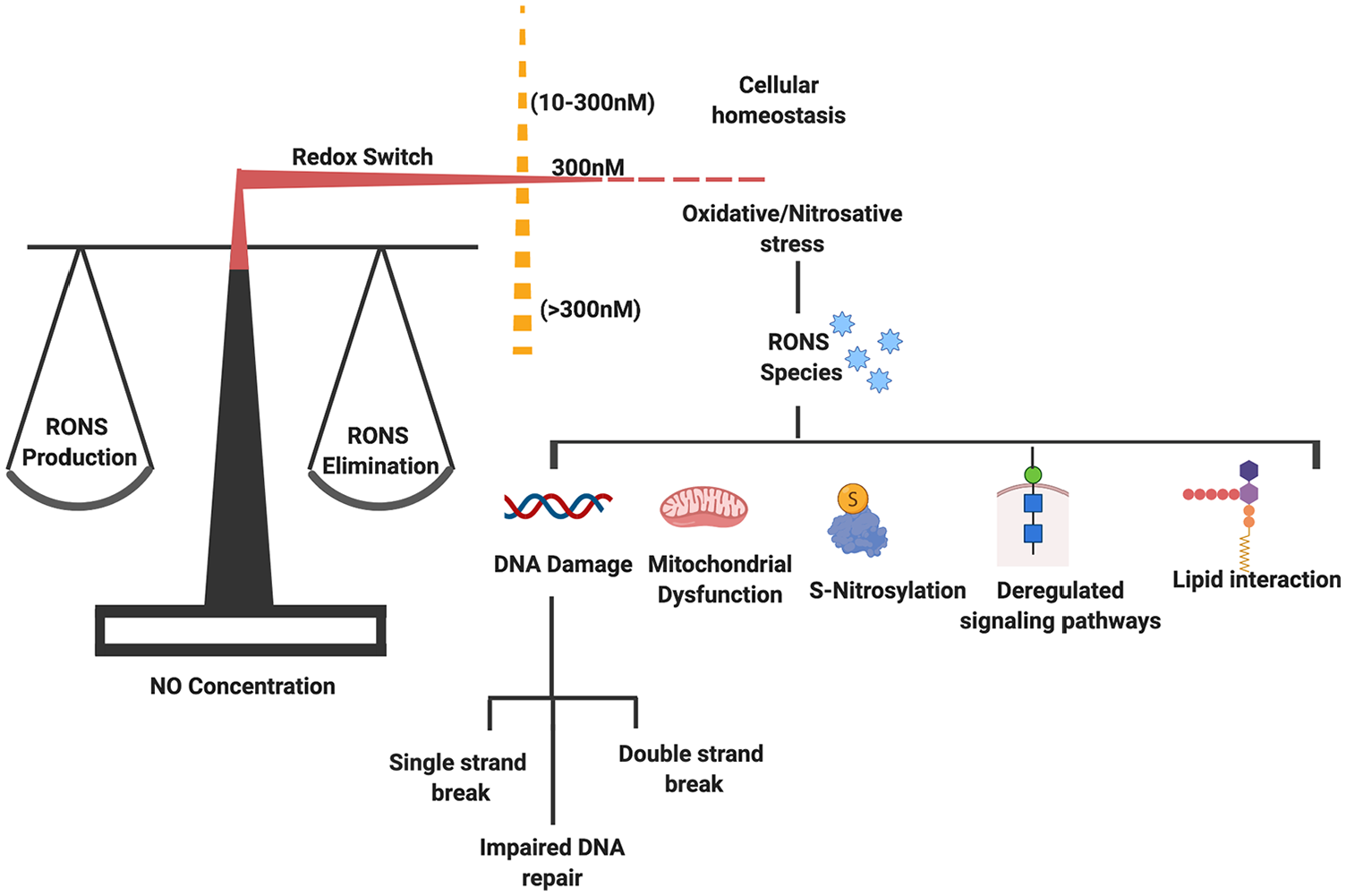
A redox switch mechanism helps in maintaining a fine balance between ROS/RNS generation and elimination. However, a lower concentration of NO helps in maintaining cellular homeostasis. An imbalance in their level results in generation of oxidative and nitrosative stress. The reactive species (RONS) play a role in development of pathological condition via acting on different pathways and causing varied effects in the form of DNA damage, mitochondrial dysfunction, deregulated S-nitrosylation, and lipid interaction. NO indicates nitric oxide; RNS, reactive nitrogen species; RONS, reactive oxygen and nitrogen species; ROS, reactive oxygen species.
The products formed after oxidation reactions (oxidation of glutathione [GSH]), nitration (nitrotyrosine formation), or nitrosation (S-nitrosoglutathione [GSNO] formation) are of high biological significance and help in generation of more NO donors and deregulate cell signaling. 18 Posttranslational modifications induced after RNS exposure often lead to differential interactions with other cellular targets and induce varied effects depending on their local concentration. 6 RONSs cause modifications such as tyrosine nitration, S-nitrosylation, S-sulfenylation, S-glutathionylation, and cysteine oxidation in residues of regulatory protein that play critical role in carcinogenesis. Furthermore, they also cause alterations in the activities of transcription factors and intracellular effectors of different signaling pathways. 7
Posttranslational Modifications Mediated by NO
Reactive nitrogen species are produced under normal physiological conditions in cells and are monitored and properly controlled by different mechanisms to maintain redox homeostasis, ie, proper regulation of oxidative and nitrosative stress. To maintain normal cellular physiology, redox mechanism plays a lead role in the regulation of signal transduction pathways involved in proper functioning of cellular targets that are prone to malfunction associated with human disease. As mentioned previously, NO mediates some of the major types of protein posttranslational modifications (PTMs) including S-nitrosylation, S-glutathionylation, and tyrosine nitration. 19 Mechanistically, NO regulates protein function by modification of cysteine thiol residues and transition metal centers, by S-nitrosylation of single critical cysteine residue present inside an acid-base or hydrophobic structural motif, or by inducing specific signals after PTM that can be used for self-defense against microbes and cancer cells. 20
S-nitrosylation is one of the most important types of PTM in which an NO group is covalently attached to the thiol side chain of a cysteine residue. This type of PTM serves as a critical mechanism behind redox-based physiological regulation and plays an important role in posttranslational regulation of a wide variety of protein influenced by NO. It is now evident that proteins from almost all functional classes are substrates for S-nitrosylation, and the entire process of S-nitrosylation and de-nitrosylation is highly specific and is tightly regulated by virtue of structural motifs, allosteric regulators, and molecular interactions between target protein and NOS. 21 S-nitrosothiols (SNOs), resulting from S-nitrosylation, may also cause alteration in cellular function of a variety of proteins. 21 , 22 S-nitrosoglutathione, a low-molecular-weight SNO formed after S-nitrosylation of antioxidant GSH, is the major type of SNO in the cell and serves as an NO reservoir in cells. 23 , 24 GSNO also facilitates transnitrosylation reactions and transfers its NO group to a new cysteine thiol group. 21 As S-nitrosylation is a highly regulated, reversible mechanism, generated SNO can be broken down either enzymatically by thioredoxins in a very specific manner 25 or non-enzymatically by antioxidants such as ascorbate or GSH. Indirectly, S-nitrosoglutathione reductase (GSNOR) also controls SNO levels by decomposing GSNO. 26
The other important type of PTM is S-glutathionylation. It is the reversible process in which GSH is covalently added to cysteine residues in target proteins and results in alteration in molecular charge, mass, and structure-associated function and might prevent protein degradation via proteolysis or sulfhydryl overoxidation. Presence of excess S-glutathionylated proteins in serum may also serve as useful oxidative or nitrosative stress biomarkers in diseased individuals. 27 This process has emerged as a candidate mechanism in maintaining intracellular redox state by having a control over the generation of RONS associated with a functional responses and stress signaling. 19 Although the process of tripeptide GSH addition is promoted by oxidative and nitrosative stress, it also occurs in the cell under unstressed conditions. As proper folding of proteins in endoplasmic reticulum is dependent on balanced redox environment, a redox stress condition in the ER can affect rates of S-glutathionylation. 28 Glutathione acts as a biological redox buffer and a balanced GSH (reduced)/GSSG (oxidized) ratio is maintained via GSH peroxidase and GSH reductase enzymes in controlled oxidation/reduction reactions. A decrease in GSH mediated by RONS may cause cell death via apoptosis or necrosis. 29
Protein tyrosine nitration is another type of PTM and is caused by covalent modification by NO-derived oxidants such as peroxynitrite (ONOO−) and nitrogen dioxide (•NO2) and involves the formation of intermediate Tyr• radical from tyrosine. 30 Peroxynitrite is a potent, short-lived, oxidizing, and nitrating agent; by its secondary radicals, it can directly or indirectly promote protein and lipid modifications. 31 Nitration of protein tyrosine residues occurs when a nitro group (-NO2) substitutes hydrogen at the third position in the phenolic ring, leading to the formation of 3-nitrotyrosine (3-NT) as product. 32 The formation of 3-NT in proteins indicates an oxidative PTM that favors pro-oxidant processes. Excess ROS along with NO and derivatives forms peroxynitrite as a nitrating species, which by tyrosine nitration modifies key properties of protein and can cause profound structural and functional effects 33 and, overall, may serve as a marker of nitroxidative stress in diseased condition. 34
Dual Role of NO in Cancer
Nitric oxide depending on its concentration and locations plays dichotomous role in cancer development. Its protumor as well as antitumor effect makes it a very interesting molecule in the tumor microenvironment. At lower concentrations, NO supports carcinogenesis, whereas at higher concentration, it becomes cytotoxic to cancer cells and induces apoptosis by forming peroxynitrite. 35 As reviewed in detail by Choudhary et al, NO plays tumoricidal or tumor-inhibiting role, depending on its concentration and location. Nitric oxide has cytostatic or cytotoxic effect on the growth of cancer cells by shifting cellular metabolism, inhibiting DNA synthesis, activating caspases, and upregulating expression of multiple apoptosis-associated proteins or it plays a tumor-promoting role by different mechanisms, including (a) genotoxic effect—creating toxic and mutagenic species, directly inducing modification in DNA in the form of strand breaks and nucleic acid oxidation and deamination, impaired DNA repair; (b) antiapoptotic effects—by inhibiting caspase activity by S-nitrosylation of the cysteine residue, causing loss of p53 repressor activity GC to AT mutations, increasing B-cell lymphoma 2 (Bcl)-2 expression, activating cyclo-oxygenase (COX), blocking cytochrome C release, and suppressing ceramide generation; (c) angiogeneic effect—via dilating eNOS-mediated arteriolar vessels, increasing hyperpermeability of vascular endothelium, vascular endothelial growth factor (VEGF) release, increase in permeability of tumor vasculature, and production of proangiogenic factors; (d) metastatic effect—via upregulated expression of matrix metalloproteinase 2 (MMP2), MMP-9, and VEGF; (e) dampened immune response against tumor—by suppressed leukocyte proliferation and infiltration. 17
As NOSs are ubiquitously expressed in different malignant cancers, NO derived from cancer cell promotes cancer progression, whereas NO derived from host stromal cell acts differentially and inhibits growth of NO-sensitive cancers but promotes growth of NO-resistant cancers15. There is increasing interest in studying the role of NO and NOSs in cancer growth and progression, and studies conducted in different cancers, including brain, 36 , 37 gastric, 38 colorectal, 39 lung, 40 prostate, 41 bladder, 42 head and neck, 43 pancreatic, 44 and breast cancer,45-50 emphasize its critical role in these various cancers.
Nitric Oxide and S-Nitrosylation Implications in Breast Cancer
Breast cancer is highly heterogeneous, displaying great heterogeneity in different areas of the same tumor in ER, PR, and HER2 expression. 1 It was found that in invasive ductal carcinoma (a breast cancer subtype), NO biosynthesis was significantly higher in grade III tumors with respect to grade II, and it was further reported that NOS expression correlates with tumor grade in breast cancer. 51 After investigations in benign, malignant, and normal breast tissue samples, an inverse correlation was found between metastatic potential of tumors and presence of iNOS protein in tumor cell, and it was suggested that tumor cell–mediated endogenous iNOS expression might play an inhibitory effect on the metastatic potential in breast cancer 52 or tumors with high NOS activity indicated a lower proliferation rate and grade. 53 However, interestingly, it was shown by Vakkala et al 54 that iNOS-positive breast tumor and stromal cells showed increased vascularization and higher apoptotic indices, suggesting iNOS-mediated angiogenesis and apoptosis-promoting role in breast carcinoma (Figure 3).

NO plays a role in both tumor progression as well as tumor regression depending on the concentration of NO in the tumor microenvironment. A lower concentration of NO promotes tumor progression by increasing their proliferation, migration, invasion, and chemoresistance, whereas a higher concentration of NO has cytotoxic effects and causes inhibition of their growth and apoptosis. EMT indicates epithelial-to-mesenchymal transition; NO, nitric oxide.
Studies on the role of eNOS in breast cancer concluded that eNOS is expressed in breast tumors and its expression positively correlates with ER expression and negatively with tumor grade and lymph node status. 55 Expression of NOS has been observed in advanced breast cancers 56 , 57 and breast cancer cell lines. 58 Previous findings have suggested that tumor-derived eNOS promotes the growth and metastasis of tumors by stimulating migration, invasiveness, and angiogenesis in tumor cells. 59 NO-associated nitro-tyrosine levels were also found to be correlated with reduced disease-free and overall survival in breast cancer. 49 In benign breast cancer tissues, eNOS synthase expression was predominantly found localized in apocrine metaplastic cells and entire endothelia, whereas no eNOS activity was found in infiltrating duct carcinoma cells of poorly differentiated type. 60 In breast cancer, the tumor microenvironment 45 and its metabolism 61 also play a significant role in chemoresistance and metastasis in an NOS2-dependent manner.
Steroid hormones, estrogen and progesterone, both control NOS activity; estrogen increases the eNOS activity, whereas progesterone activates iNOS activity. An increased apoptosis rate in response to progesterone is associated with high levels of NO produced by iNOS, whereas low levels of NO produced by eNOS could be behind the proliferative effect of estrogen. All these findings implicate correlation between NOS expression and hormones in breast cancer development. 62 Furthermore, a gene signature analysis performed on estrogen receptor–negative (ER−) breast cancer associated with poor disease outcome revealed that Ets-1, a transcriptional mediator of oncogenic NO signaling, promotes the aggressive phenotype in ER− breast cancer through Ras/MEK/ERK signaling pathway. 63
c-Src acts as an upstream regulator of the estrogen-stimulated phosphatidylinositol 3-kinase (PI3K)/Akt/eNOS signaling pathway. Estrogen results in rapid activation of eNOS by PI3K/Akt-dependent Src kinase. Estrogen causes a complex formation between ER, c-Src, and P85 (PI3K regulatory subunit) and results in the activation of PI3K and Akt. 64 As already discussed above, NO promotes cancer by activating several oncogenic signaling pathways, such as PI3K/Akt and ERK-1/2 pathways. Protein phosphatase 2A (PP2A), a tumor suppressor, negatively regulates the same pathways that are activated by NO in cancers. Activating PP2A in ER− breast cancer would be a novel mechanism to antagonize NO signaling–mediated effects that promote breast cancer. 65
In summary, NOS2-derived NO can be considered as a driver of breast cancer progression by targeting multiple cell signaling pathways, including hypoxia inducible factor-1 alpha stabilization, COX2 activity, phosphoinositide 3-kinase/protein kinase B, mitogen-activated protein kinase (MAPK), epidermal growth factor receptor (EGFR), and Ras pathways. 66 Increased expression of NOS2 in breast cancer is associated with a basal-like gene expression pattern and is a predictor of poor survival in patients with ER− breast cancer. 67 The coexpression of NOS2 and COX2 is reported to promote tumor growth and reduce survival in patients with ER− breast cancer by their cross-talk. 68 A better understanding of steroid hormones and molecular mechanism of their interactions with NO will help in development of novel ways and strategies for effective breast cancer treatment.
The role of Ets family transcription factors has been well studied in different cancers. 69 , 70 Ets-1, a proto-oncogene and member of the same family, is known to promote invasive phenotype by supporting angiogenesis and extracellular matrix remodeling and is associated with poor prognosis in breast cancer. 71 One of the mechanisms to promote invasiveness is by binding to MMP-9 gene, which harbors a binding site for Ets-1. 72 It also acts as a downstream effector of HER2 and also increases MMP-1 expression. 73 A high expression of HER2 is associated with aggressive metastasis in breast cancer cells via MMP-1 and MMP-9 expression. 74
Nitric oxide–mediated S-nitrosylation regulates a wide variety of protein functions. The specificity with which this process targets critical cysteine residues and signals protein-protein interactions is controlled by different acid-base and hydrophobic motifs. 75 It has been recently identified that more than 3000 proteins impacted by NO signaling are largely managed by S-nitrosylation, and a basal level of S-nitrosylation helps in maintaining tissue homeostasis. 76 An increased level of intracellular NO leads to elevated S-nitrosylation in breast cancer and has emerged as an important mechanism promoting breast carcinomas. S-nitrosylation of Ras leads to activation of Ets-1 caused by MAPK-dependent phosphorylation and results in an aggressive breast cancer phenotype. 77 S-nitrosylation of H-Ras also restricts Raf-1 activation and further signals propagation via ERK-1/2. 78 It has also been reported that NO signaling leads to tyrosine phosphorylation of EGFR. Further analysis revealed that S-nitrosylation of EGFR and Src results in activation of oncogenic signaling in human basal-like breast cancer. 79 NO also results in modification of human ER structure by S-nitrosylation, which impairs its DNA-binding activity in turn, leading to obstruction of estrogen-dependent gene transcription. 80
Specifically, S-nitrosylation also plays a critical role in breast cancer angiogenesis and metastasis by targeting different pathways. As recently reviewed in detail by Ehrenfeld et al, 81 S-nitrosylation of target proteins promotes tumor cell epithelial-to-mesenchymal transition (EMT) and helps in their migration and invasion by promoting adhesion to the endothelium and intravasation and extravasation. In basal-like human breast cancers, Src and EGFR S-nitrosylation activates a network of oncogenic signaling and leads to increased EMT and stem cell–like phenotype and contributes to chemoresistance. 79 NO-mediated S-nitrosylation of c-Src kinase at Cys (498) residue stimulates its kinase activity and helps in cancer cell invasion and metastasis. 82 In ER− breast cancers, S-nitrosylation activates different target molecules, such as EGFR, Src, Ras, and CD63 which further initiate associated oncogenic signaling pathways including Ras/ERK, PI3K/Akt, nuclear factor κB, β-catenin, and AP-1.63,83 Ras is reported to be abnormally activated in different types of breast cancers showing overexpression of EGF/ErbB-2 receptors; its activation was further found to be related to MAPK activity. 84 S-nitrosothiol formation by S-nitrosylation and its homeostasis is often impaired in many cancers. In HER2+ breast cancers, alteration in SNO homeostasis gives a survival advantage to the tumors and reduces their trastuzumab sensitivity. 85
S-nitrosoglutathione reductase, an oxidoreductase, helps in denitrosylation and is capable of reducing NO completely 86 ; a mediated targeting could be effective in breast cancer treatment. Flavone also restrains NO production and lessens protein S-nitrosylation in breast cancer cells by inhibiting NOS activity in a dose-dependent manner and, therefore, could be of anticancer use. 87
Nitric Oxide–Mediated Strategies for Cancer Treatment
Accumulating evidence implicates defective levels of NO with different diseases. Lower concentration (picomolar to nanomolar range) of NO is present in normal physiological conditions in a cell, but a sudden increase in the concentration (micromolar range) results in development of pathological conditions. The concentration of not only NO but also other reactive species, generated after reacting with NO, strongly contributes to pathological conditions. 88 Peroxynitrite, generated from NO and superoxide anion in diffusion-controlled reaction, causes a wide range of cellular alterations by interacting with DNA, lipids, and proteins either directly via oxidative reactions or indirectly via radical-mediated mechanisms. Peroxynitrite generation triggers cellular responses causing cell necrosis or apoptosis which have been implicated in disease conditions such as heart failure, diabetes, infectious diseases, neurodegenerative diseases as well as cancer.89-91
Designing novel therapeutics to modulate NO bioavailability by finding ways to increase NO synthase activity, strengthening nitrate-nitrite-NO pathway, designing novel-class drugs for NO-donating function, and limiting NO metabolism and also finding ways to regulate downstream targets such as phosphodiesterases and soluble guanylyl cyclases will be of value. 92 As either an excess or an absence of NO may lead to different pathologies, different strategies of NO regulation show great potential. In conditions of decreased NO production caused by arginine deficiency (reduced arginine production from citrulline), a supplementation of citrulline might be curative. 93 Apart from citrulline, targeting other limiting factors such as tetrahydrobiopterin (BH4) could also be of benefit. Other than arginine or citrulline supplementation, additional approaches such as providing NO donors in the form of inhaled NO and other nitrite sources, NOS3 regulating agents, or targeting endogenous NOS inhibitors such as asymmetric dimethylarginine could have potential therapeutic benefits. 94 , 95 Use of NO-donating nonsteroidal antiinflammatory drugs (NSAIDs), such as NO-aspirin and NO-ibuprofen, have shown to decrease prostate cancer cell growth and induce apoptosis in a dose-dependent manner. 96 A wide variety of other NO donors have been shown to be effective in treating prostate and bladder cancers. 97 Finding similar use of NO donors in breast cancer may be of high benefit (Figure 4).

Strategies to increase NO bioavailability in the tumor microenvironment: Use of prodrugs, NO donors, dietary molecules, photodynamic therapy, arginine and citrulline supplementation, and blocking tumor stroma interaction could be effective in treating cancers. NO indicates nitric oxide; PDT, photodynamic therapy.
Other therapeutic ways are to deliver NO specifically to solid tumors by using polymer NO donors, 98 or nanoparticle-based systems could be useful. 99 Photodynamic therapy (PDT) is a different therapeutic approach used in treatment of certain cancers and involves the use of photosensitizer (PS) and irradiation with light of specific wavelength. PDT treatment causes iNOS/NO induction in the tumor as well as its microenvironment and could play significant role in NO-mediated cytotoxicity and act as a chemosensitizing agent. 100 The use of NO prodrug such as JS-K which is a NO-releasing diazeniumdiolate(s) has also shown to be effective in different cancers. 101 Use of potential dietary chemopreventive agents such as magnolol, 102 cardamom, 103 and curcumin 104 has shown to be effective against NO-induced gene modulation and tumor progression. As reviewed in detail by Vahora et al, 35 use of dietary agents would be of benefit in NO-mediated halting of cancer progression in different cancers, including breast cancer.
Progress made in the last few years in the area of NO and the mechanism behind its action has exponentially increased the overall understanding of NO signaling. Research performed to identify novel strategies via identifying promising new drug candidates, different dietary constituents, or other mechanisms will help in NO-mediated targeting of tumor cells.
Conclusions
Previous research has helped us to gain an insight into the dual role of NO. Biphasic response to NO in cancers is dependent on its levels and may inhibit or promote cancer growth and survival. Expression of several genes involved in tumor biology is regulated by NO and largely by NO-mediated PTM of proteins. Of these, S-nitrosylation has been depicted as a process involved in every phase of cancer progression. It affects a wide variety of proteins important in maintaining cellular functions. Affected proteins lead to cellular dysfunctions contributing to cancer onset, growth, progression, invasion, and metastasis. The exact role of NO in different cancers is determined by the primary organ affected, stage of cancer, and types of cells constituting the tumor microenvironment. Further research into regulation of critical proteins in cancer by RNS may be helpful in developing targeted therapies for cancer and, in particular, breast cancer.
Author Contributions
DM and VP contributed equally in the preparation of manuscript. All authors contributed to discussions and writing of the manuscript.
Footnotes
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by discretionary funds from the School of Graduate Studies, Rutgers University.
Declaration of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.