
Abstract
D-amino acid oxidase (DAO) is a flavoenzyme that metabolizes D-amino acids by oxidative deamination, producing hydrogen peroxide (H2O2) as a by-product. The generation of intracellular H2O2 may alter the redox-homeostasis mechanism of cells and increase the oxidative stress levels in tissues, associated with the pathogenesis of age-related diseases and organ decline. This study investigates the effect of DAO knockdown using clustered regularly interspaced short palindromic repeats (CRISPR) through an in silico approach on its protein-protein interactions (PPIs) and their potential roles in the process of aging. The target sequence and guide RNA of DAO were designed using the CCTop database, PPI analysis using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database, Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) analyses, Reactome biological pathway, protein docking using GalaxyTongDock database, and structure analysis. The translated target sequence of DAO lies between amino acids 43 to 50. The 10 proteins that were predicted to interact with DAO are involved in peroxisome pathways such as acyl-coenzyme A oxidase 1 (ACOX1), alanine-glyoxylate and serine-pyruvate aminotransferase (AGXT), catalase (CAT), carnitine O-acetyltransferase (CRAT), glyceronephosphate O-acyltransferase (GNPAT), hydroxyacid oxidase 1 (HAO1), hydroxyacid oxidase 2 (HAO2), trans-L-3-hydroxyproline dehydratase (L3HYPDH), polyamine oxidase (PAOX), and pipecolic acid and sarcosine oxidase (PIPOX). In summary, DAO mutation would most likely reduce activity with its interacting proteins that generate H2O2. However, DAO mutation may result in peroxisomal disorders, and thus, alternative techniques should be considered for an in vivo approach.
Keywords
Introduction
The World Health Organization estimates that the world’s population of those more than 60 years will double from 12% to 22% between 2015 and 2050, as the pace of population aging is faster now compared with the past. According to the National Institute on Aging (NIA), US Department of Health and Human Services, aging involves dynamic changes in a living organism’s biological, physiological, environmental, psychological, behavioral, and social processes. 1 In trying to identify the biological characteristics and factors that contribute to aging, researchers have included 9 hallmarks of aging 2 and the 7 pillars of aging 3 that were aligned to outline the following aging mechanisms: (1) genomic instability/abnormal gene expression, (2) DNA damage, (3) telomere attrition, (4) epigenetic alterations, (5) loss of proteostasis, (6) deregulated nutrient sensing/metabolism, (7) mitochondrial dysfunction, (8) oxidative stress, (9) cellular senescence, (10) stem cell exhaustion, (11) altered intercellular communication, and (12) inflammation.
From the growing body of evidence, there is a large focus on oxidative stress theory that results in the irreversible accumulation of oxidative damage, due to reactive oxygen species (ROS), such as superoxide radical (O2•−), hydroxyl radical (HO•), and hydrogen peroxide (H2O2). 4 Reactive oxygen species contribution to aging is also governed by the cellular metabolism, protective systems, and genotype, where unintended cellular activities result in damage in the form of by-products, errors, and misbalance in cellular components. 5 The overproduction of ROS can disturb the delicate redox balance within cells, resulting in the impairment of essential cellular components to the damage of primary cellular components including DNA, lipids, and proteins. 6 Reactive oxygen species can cause oxidative damage to these cellular structures, leading to various adverse effects on cell function, which eventually linked to aging and the progression of age-related diseases, such as cardiovascular disease, neurodegenerative disease, and diabetes. 7 While mitochondria are the leading player in ROS production, peroxisomes also produce significant amounts of ROS, and their dysfunction may contribute to cell death.8,9 Peroxisomes are highly dynamic organelles consisting of H2O2-producing oxidases and the antioxidant enzyme, namely catalase (CAT), to detoxify the ROS.
Notably, accumulation of D-amino acids has been associated with age-related diseases, including long-term kidney disease, Alzheimer disease, and Parkinson disease. 10 An enzyme known as D-amino acid oxidases (DAO/DAAO, from now on addressed as DAO) catalyzes the oxidative deamination of D-amino acids to produce α-keto acids, ammonia, and H2O2. 11 In mammals, DAO is primarily expressed in the kidney, liver, brain, and to a lesser extent, in the small intestine, and neutrophilic leukocytes. 12 In humans, DAO (1) eliminates D-amino acids in the liver and kidney, (2) modulates D-serine which is required for the activation of N-methyl-D-aspartate (NMDA) receptor in the brain, and (3) expresses in granule fraction of mature human granulocytes in counteracting foreign microorganisms. 13
Recently, a study identified DAO upregulation in cellular senescence, persistent DNA damage, and oxidative stress. 14 D-amino acid oxidase produces ROS by-products of substrate oxidation, and ROS accumulation mediates senescence. By knocking out DAO in mice, previous study has found that there was an enhancement of D-serine exposure in the brain, which improves short-term memory, and suppression of hydrogen peroxide generation in the kidney. 15 However, the long-term effect of DAO knockout and knockdown has been unclear. D-amino acid oxidase is a peroxisomal enzyme, which may interact with other proteins and impact on cellular homeostasis. Considering these, this study aimed to assess the effect of DAO knockdown using CRISPR through an in silico approach and predict its protein-protein interactions (PPIs), as mutations may affect their binding ability and the functional interactions between the target and interacting proteins.
Materials and Methods
Sequence retrieval from National Center for Biotechnology Information
The consensus coding sequences (CCDS) of DAO were retrieved from the National Center for Biotechnology Information (NCBI) (https://www.ncbi.nlm.nih.gov/). Both nucleotide (accession No. NM_001917.5) and protein (accession No. NP_001908.3) sequences of DAO were retrieved for further analysis.
Designing guide RNA and target sequence prediction
The nucleotide sequence of DAO was used to predict the guide RNA (gRNA) sequence and the target sequence (5′ end of the gRNA) using the CCTop (http://crispr.cos.uni-heidelberg.de/). 16 The gRNA of DAO was predicted and chosen according to its efficacy score.
Mapping of target sequence onto DAO protein sequence
The best-predicted target sequence based on the in silico analysis was mapped onto the DAO nucleotide sequence to identify the location. The target sequence was mapped onto the DAO amino acids sequence to obtain the amino acid position of the target sequence in the DAO protein sequence.
Analysis of protein-protein interaction
D-amino acid oxidase protein interaction analysis was carried out using the Search Tool for the Retrieval of Interacting Genes/Proteins (STRING) database version 11.5 (https://string-db.org/). 17 D-amino acid oxidase protein was submitted as the query protein to identify the predicted interacting proteins. The analysis was conducted with the highest confidence level settings (0.9) to avoid false positive or less significant interactions. The interaction data were then exported in tab-separated values (TSV) format, and the network illustration was exported in jpg format.
Analysis of gene ontology and Kyoto Encyclopedia of Genes and Genomes enrichment
The gene list was submitted to Database for Annotation, Visualization and Integrated Discovery (DAVID) database version 6.8 (https://david.ncifcrf.gov/) as a query in the search box to predict the gene ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways involved. 18 Gene Ontology analysis was carried out to specify the cellular locations, functions, and biological processes over-represented in the gene set. 19 Kyoto Encyclopedia of Genes and Genomes analysis was conducted to identify the metabolic pathways involved in the gene set. 20
Analysis of biological pathway using Reactome
Pathway analysis of proteins interacting with DAO was carried out using the Reactome Knowledgebase database version 79 (https://reactome.org/). 21 Reactome analysis was conducted to specify cellular compartments and protein localization participating in reactions, forming a network of biological interactions grouped into pathways involved in the gene set.
Preparation of protein structures
Three-dimensional (3D) protein structures were retrieved from RCSB PDB (Research Collaboratory for Structural Bioinformatics Protein Data Bank) (www.rcsb.org) 22 for the following human DAO (PDB ID: 3G3E), AGXT (PDB ID: 5F9S), CAT (PDB ID: 1DGF), CRAT (PDB ID: 1NDF), and HAO1 (PDB ID: 2NZL). For protein without experimental structures such ACOX1, HAO2, GNPAT, PIPOX, PAOX, and L3HYPDH, homology models were built using SWISS-MODEL (https://swissmodel.expasy.org/). 23 The protein structures were prepared using the The University of California, San Francisco (UCSF) Chimera (https://www.cgl.ucsf.edu/chimera/). 24 The protein structures were dock prepped by removing bound ligands and solvent molecules from the protein structures. The prepared protein structures were then saved in protein data bank (pdb) format for docking analysis.
Analysis of protein docking
Docking analysis of the target protein (DAO) with the interacting proteins was conducted using GalaxyTongDock (http://galaxy.seoklab.org/tongdock). 25 D-amino acid oxidase protein structure was submitted as chain A and each interacting protein structure as chain B in GalaxyTongDock. Each pair of docking structures were downloaded in pdb format for structure analysis.
Analysis of the docked structure
Each docking pairs were analyzed using Solvent accessibility based Protein-Protein InterfaceiDEntification and Recognition (SPPIDER) database to predict the binding residues between the proteins (http://sppider.cchmc.org/). 26 The interface residues for each docking pair were saved for further analysis. The interface residues were compared with the CRISPR target sequence of DAO (gRNA) to predict the effect on the binding of the interacting proteins.
Results
D-amino acid oxidase consensus coding sequences
The CCD represents the protein-coding region of a gene. Nucleotide and protein sequences of DAO were obtained from NCBI (CCDS ID: CCDS9122.1). Nucleotide sequences followed by the protospacer adjacent motif (PAM) site NGG (5′-20 N-NGG-3′) were identified using CCTop. The top 10 gRNAs identified for DAO had an efficacy score of 0.74 to 0.83, as shown in Table 1 and Supplemental Table 1.
Top 10 gRNA sequences retrieved from CCTop.

While the efficacy score for all gRNAs was high, in our past studies, we have shown that the gRNA with the best in silico score also performed the best in subsequent downstream assays. Therefore, for the purpose of this study, the gRNA target sequence (5′-GCCGGCAGCCACGTCGGTGGTGG-3′), with the highest efficacy score (0.83), PAM sequence (TGG), was selected for this study. The predicted nucleotide target sequence lies between positions 108 885 134 to 108 885 156 at chromosome number 12.
Target sequence mapping onto DAO protein sequence
The DNA target sequence was mapped onto the DAO nucleotide sequence. Then, the target sequence was mapped onto the DAO protein sequence. The translated target sequence position found through this mapping method lies from amino acid positions 43 to 50 (tttdvaag), indicated in bold lowercase in the amino acid sequence, as shown in Figure 1.

Amino acid sequences of D-amino acid oxidase.
PPI analysis
Network analyses were conducted to identify the interacting partners of DAO predicted by the STRING database. D-amino acid oxidase is found to have direct or indirect interaction with 10 proteins (ACOX1, AGXT, CAT, CRAT, GNPAT, HAO1, HAO2, L3HYPDH, PAOX, and PIPOX) which were gathered from various sources and evidence, most of which came from curated databases, text mining, and co-expression data as illustrated in Figure 2.

D-amino acid oxidase interaction network extracted from STRING.
All the predicted proteins have an interaction score between 0.944 and 0.975, which is a very high confidence score, suggesting that the interaction of DAO with the predicted protein partners, either physically or functionally, is highly plausible, as shown in Table 2. Since STRING provides information on physical and functional interactions, functional enrichment analysis was conducted to determine each protein’s GO and the significant pathways involved among the predicted proteins and DAO.
Predicted functional partners of DAO from STRING.

GO and KEGG enrichment
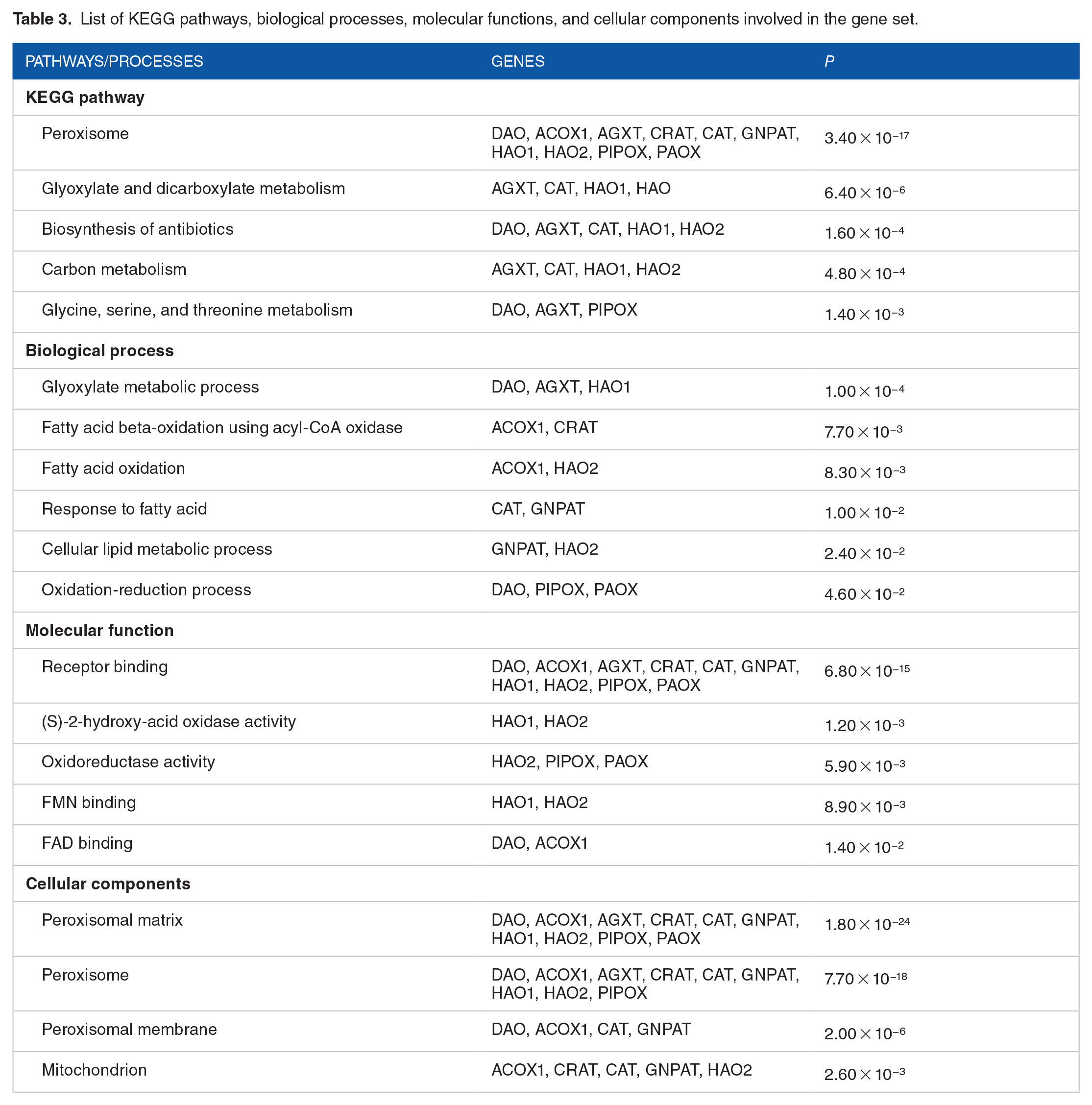
Based on KEGG analysis, 5 pathways are significantly enriched by the gene set, as shown in Table 3. Ten of the interacting genes and DAO are most associated considerably with the metabolic processes in the peroxisome. Five significant processes with a P value less than .05 were involved in the gene set. The glyoxylate metabolic process is the most significantly enriched process by the gene set (P < .05) as the value is the closest to 0 compared with the rest of the results. The most significant molecular function obtained is receptor binding, which is shown by 10 genes. D-amino acid oxidase, ACOX1, AGXT, CAT, CRAT, GNPAT, HAO1, HAO2, PAOX, and PIPOX have receptor binding functions where the proteins can bind with other proteins in functional or physical interactions. The target gene and all interacting genes, except for L3HYPDH, encode peroxisome-localized proteins.
List of KEGG pathways, biological processes, molecular functions, and cellular components involved in the gene set.
Reactome pathway analysis
Based on Reactome analysis, 10 proteins (ACOX1, AGXT, CAT, CRAT, DAO, GNPAT, HAO1, HAO2, PAOX, and PIPOX) were found as participants (members of) cargo proteins containing peroxisomal targeting signal 1 (PTS1) (ID: R-HSA-9033111 and R-HSA-9033148). This protein plays a role in the peroxisomal protein import pathway (ID: R-HSA-9033241), located in the cytosol, peroxisomal matrix, and peroxisomal membrane.
Protein-protein docking
After obtaining docked protein structures from GalaxyTongDock, the structures were analyzed through SPPIDER database to predict the interacting residues of the protein structures. Referring to the gRNA map translated to amino acid sequence, the mutation site should be at T43 T44 T45 D46 V47 A48 A49 G50. As indel mutations in the target gene area would most likely alter the downstream amino acid sequence, the binding of DAO to its interaction partners may be disrupted. This is demonstrated in Table 4, which shows that most DAO binding residues are downstream of T43 and bolded.
Protein docking structures with the respective interacting residues.
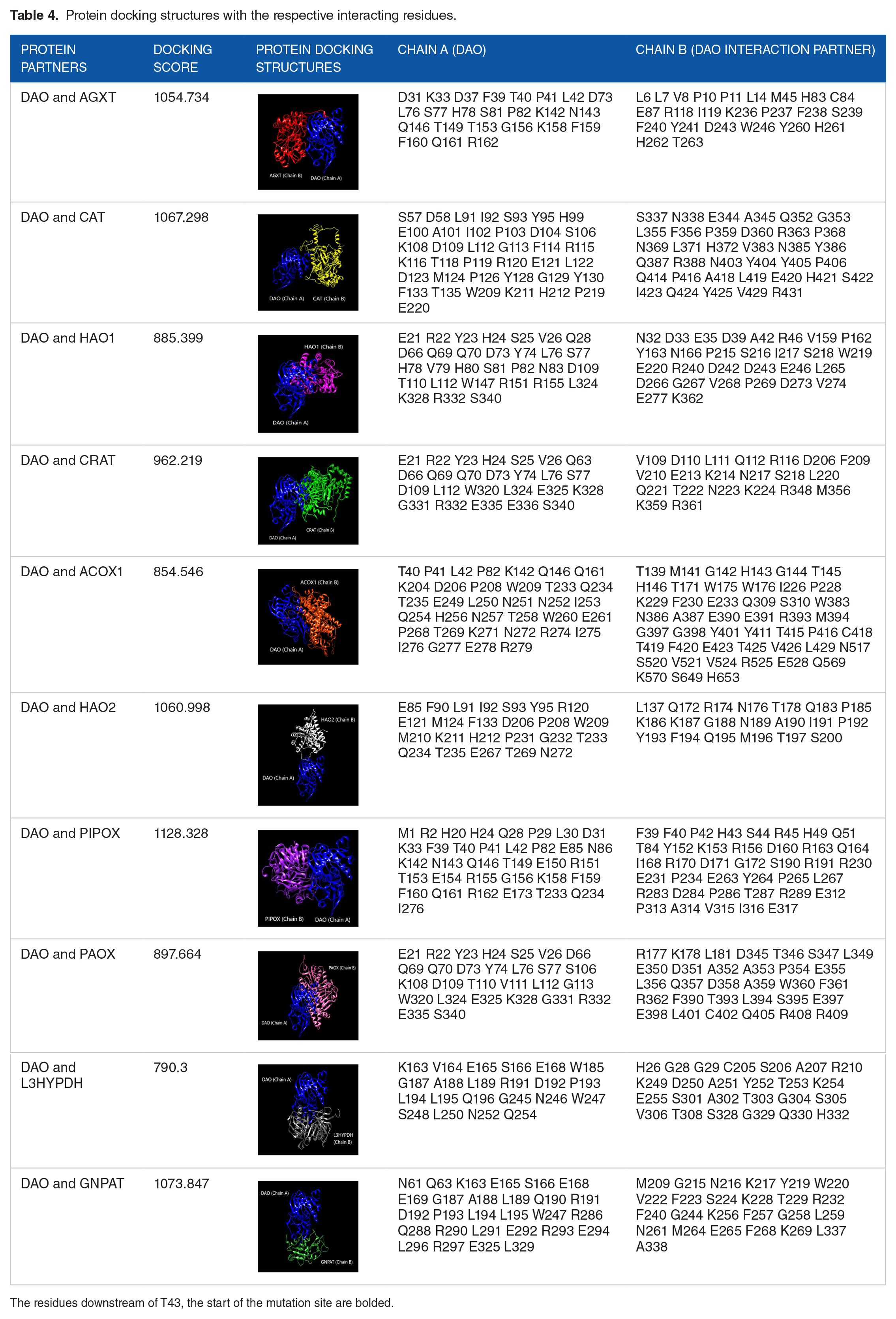
The residues downstream of T43, the start of the mutation site are bolded.
Discussion
DAO catalyzes D-amino acids via three mechanisms.27–29 : (i) dehydrogenation of α-D-amino acids to their imino acid counterparts 270 with concurrent reduction of FAD to FADH2, (ii) the FADH2 is redeoxidised by molecular oxygen 271 to produce H2O2, (iii) hydrolysis of the imino acid to give the corresponding α-keto acid and 272 ammonium. Excess H2O2 increases oxidative stress and is widely recognized as a critical initiator 273 of senescence, and their accumulation is widely observed in senescent cells. 29
We hypothesized that knocking down DAO using CRISPR/Cas9 would inhibit DNA damage-induced senescence. In this study, the best sgRNA target site of DAO was mapped onto the CCDS region to identify the translated target amino acid sequence. The target sequence lies at chromosome 12, between 108 885 134 and 108 885 156 bp, from positions 43 to 50. Mutations in this region might affect its PPI as the interface residues between the proteins might be affected due to the DAO gene knockdown.
Further STRING analysis predicted 10 interacting proteins with DAO based on evidence. GO analysis was carried out to identify the functional interaction between these proteins to better understand the mechanism at cellular levels and their localization. Nine out of 10 of the interacting proteins were found to be peroxisomal proteins (Table 3). Peroxisomes synthesize and degrade various critical cellular compounds. 30 Particularly, H2O2 that is produced is immediately processed by CAT by the peroxisome. However, in certain conditions where the balance of H2O2 production is altered, there is an accumulation of potentially toxic metabolites, putting the cells on a pro-aging perturbation. The following peroxisomal proteins were found to have functional interactions with DAO: (1) AGXT, (2) CRAT, (3) HAO1, (4) HAO2, (5) ACOX1, (6) PIPOX, (7) PAOX, and (8) GNPAT.
AGXT encodes serine-pyruvate aminotransferase, which catalyzes the transamination of glyoxylate to glycine, and deficiency leads to overproduction of oxalate and glycolate in primary hyperoxaluria type 1 (PH1). 31 Upstream of the AGXT reaction, the flavin mononucleotide (FMN)-dependent enzymes hydroxyacid oxidases (HAO1 and HAO2) catalyze the oxidation of glycolate to glyoxylate. 32 Therefore, AGXT and HAO are involved in glyoxylate detoxification in the human hepatocytes. In particular, HAO action releases H2O2 in the peroxisomes. 33
Besides AGXT and HAO, other H2O2-producing oxidases include PIPOX and PAOX, which are involved in amino acid and polyamine catabolism, respectively. PIPOX converts L-pipecolic acid to 1-piperideine-6-carboxylic acid, and deficiency in the liver is associated with patients with human peroxisome biogenesis disorders (PBDs) like Zellweger syndrome. 34 On the contrary, PAOX preferentially oxidizes polyamines such as N 1 -acetyl-spermine or N 1 -acetyl-spermidine to produce spermidine, and elevated polyamines level results in tumor progression. 35 Carnitine o-acetyltransferase encodes carnitine acyltransferases that catalyze acyl exchange between carnitine and coenzyme A (CoA), and reduced activity causes serious neurological and heart problems, similar to that seen in patients with Alzheimer disease. 36
In ether lipid biosynthesis, GNPAT encodes the enzyme GNPAT or dihydroxyacetone phosphate acyltransferase (DHAPAT) and is responsible for the production of plasmalogens. Mutations in GNPAT have been found to cause rhizomelic chondrodysplasia punctata type 2 (RCDP2), marked by shortened long bones and intellectual disability. 37 Meanwhile, in the fatty acid beta-oxidation pathway, ACOX1 encodes the first and rate-limiting enzyme of the very-long-chain fatty acid (VLCFA), and deficiency causes a rapid and severe loss of nervous system function. 38
The Reactome biological pathway analysis reported cargo proteins containing PTS1. Generally, the matrix proteins in the cytosol are imported into the peroxisome via PTS1, a carboxyl-terminal tripeptide motif and cytosolic protein, Pex5p, which is the receptor for PTS1. 39 A study found that aging compromises PTS1 protein import, affecting the antioxidant activity of CAT.40,41 Furthermore, alterations in the number and appearance of peroxisomes were observed, and the organelles accumulated the Pex5p on their membranes. The decline in Pex5p-dependent peroxisomal import of CAT resulted in increased oxidative damage to cellular proteins and lipids, thereby initiating the ”deterioration spiral,” which eventually reduces peroxisomal protein import and exacerbates the effects of aging. 42 As illustrated in Figure 3, the model described 3 potential factors: hypocatalasemia, aging, or chemical inactivation, resulting in reduced cellular CAT expression. 41

Peroxisome deterioration spiral.
Our further protein-protein docking analysis showed that all interacting proteins would most likely be unable to bind to DAO if the target sequence is mutated since all of the interacting or binding residues are located upstream and downstream of the DAO target residues. Changes in the DNA sequence generate inactivating mutations at the target site, creating frameshifts. 43 This might cause changes in the downstream amino acid residues and possibly influence how the protein folds into a 3D structure. Indels adjacent to the PPI interfaces may alter the protein-protein bindings, resulting in the loss or gain of new interactions. 44 Thus, this modification will randomly affect DAO general structure and, more crucially, its binding interface, resulting in a loss of capacity to interact with its partners.
Comparative genome-wide expression analysis revealed novel genes specifically upregulated in senescence, with DAO directly regulated by p53. 45 D-amino acid oxidase cellular senescence activation was observed when DNA damage was induced through anticancer drugs, cancerous genes, or telomere shortening. 14 In response, transcription is induced in DAO and p53-regulated genes, including solute carrier family 52 member 1 (SLC52A1) located at the cell membrane. Increased levels of SLC52A1 allow transport of vitamin B2, which converts to flavin adenine dinucleotide (FAD) abundantly in the cell. Increased FAD concentration increases DAO activity and damages cellular DNA, proteins, and lipids, thereby promoting cellular senescence.
Although most of the peroxisomal proteins (AGXT, ACOX1, HAO1, HAO2, PIPOX, PAOX, GNPAT, and CRAT) have a negative effect on aging due to their production of H2O2, as illustrated in Figure 4, deficiencies or mutations in any of these enzymes result in peroxisomal disorders. 33 In particular, the hypothetical mutation in DAO was related to amyotrophic lateral sclerosis (ALS), a fatal human disease characterized by progressive muscle weakness, atrophy, paralysis, and death from respiratory failure. When DAO activity in the spinal cord was reduced in a mice model, the D-serine levels were consequently increased. 46 The excessive D-serine released from neurons and glia leads to synaptic loss and stimulation of extra-synaptic N-methyl-D-aspartate receptor (NMDAR) currents. 47

The major producer of peroxisomal ROS and their enzymes. ACOX and CRAT in the beta-oxidation of fatty acids, DAO and PIPOX in the amino acid catabolism, GNPAT in the ether lipid biosynthesis, PAOX in the polyamine catabolism, and HAO and AGXT in the glyoxylate detoxification. Meanwhile, CAT is recognized to be involved in the antioxidant system.
In conclusion, the in silico knockdown of DAO at the predicted target site would affect its PPI network, which is involved in H2O2 production. D-amino acid oxidase may serve as a promoter of DNA damage-induced cellular senescence. Overproduction of ROS has a negative effect, and ROS released under stress is thought to cause illness and aging. However, in vivo edits using CRISPR can be done with caution, using alternatives like targeted base edits to avoid unwanted or detrimental effects in future research.
Limitations
This article focuses on the knockdown of DAO using the best gRNA predicted through an in silico analysis, resulting in a slight bias in selection of gRNA. An in-depth study of the top 10 gRNA may produce different results, enabling a broader option for selection of gRNA for subsequent experiments. In addition, the insights gained from the GalaxyTongDock analysis are confined to binding modes and energies, and a more in-depth atomic-level investigation could provide a more profound and complete understanding of the interactions involving DAO and its partner. Furthermore, it should be noted that the proteins and interactions chosen in this study represent only a fraction of very intricate biological landscape, potentially neglecting the involvement of other crucial proteins and pathways. Thus, while the study’s findings offer insightful glimpses, they should be interpreted within the expansive framework of the multifaceted mechanisms that underlie these aging-related phenomena.
Supplemental Material
sj-xls-1-bbi-10.1177_11779322241234772 – Supplemental material for Investigating D-Amino Acid Oxidase Expression and Interaction Network Analyses in Pathways Associated With Cellular Stress: Implications in the Biology of Aging
Supplemental material, sj-xls-1-bbi-10.1177_11779322241234772 for Investigating D-Amino Acid Oxidase Expression and Interaction Network Analyses in Pathways Associated With Cellular Stress: Implications in the Biology of Aging by V Kalidasan, Darshinie Suresh, Nurulisa Zulkifle, Yap Siew Hwei, Leong Kok Hoong, Reena Rajasuriar and Kumitaa Theva Das in Bioinformatics and Biology Insights
Footnotes
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by KT’s grants – Higher Education Malaysia for Fundamental Research Grant Scheme with Project Code: FRGS/1/2020/SKK0/USM/02/9 and Universiti Sains Malaysia for Research University Grant and RR grants—Ministry of Higher Education Malaysia through the Fundamental Research Grant Scheme (FP007-2019A).
Declaration Of Conflicting Interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
DS and YSH carried out the data analysis and wrote the draft article. VK reanalyzed the data and wrote the final article with support from NZ, LKH, RR, and KTD. All authors reviewed the article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.