
Abstract
The glacier ice worm, Mesenchytraeus solifugus, is among a few animals that reside permanently in glacier ice. Their adaptation to cold temperature has been linked to relatively high intracellular adenosine triphosphate (ATP) levels, which compensate for reductions in molecular motion at low physiological temperatures. Here, we show that ATP6—the critical regulatory subunit of the F1Fo-ATP synthase and primary target of mitochondrial disease—acquired an unprecedented histidine-rich, 18-amino acid carboxy-terminal extension, which counters the strong evolutionary trend of mitochondrial genome compaction. Furthermore, sequence analysis suggests that this insertion is not of metazoan origin, but rather is a product of horizontal gene transfer from a microbial dietary source, and may act as a proton shuttle to accelerate the rate of ATP synthesis.
Introduction
Numerous microbial organisms (eg, bacteria, algae, fungi) but relatively few animal taxa (eg, representatives of Annelida, Arthropoda, Rotifera) have adapted to the permanently cold and harsh conditions of glacier ice.1–3 Common to all obligatory glacial species that have been examined are paradoxically high intracellular adenosine triphosphate (ATP) levels in direct comparisons with temperate (ie, mesophilic) species.4–6 This has been interpreted as a compensatory mechanism that offsets reductions in molecular motion at cold temperatures by increasing the probability of molecular collisions with ATP, the universal currency of energy that drives most biochemical reactions. 7
The glacier ice worm, Mesenchytraeus solifugus, was the first organism in which this paradoxical energetic profile was observed 4 and subsequent efforts were aimed at determining the underlying metabolic mechanism. Our collective analyses suggest that enhanced ATP synthesis (with respect to ATP consumption) can explain the observed gains in ATP levels at cold physiological temperatures, and previous investigations targeted adenosine monophosphate (AMP) degradative machinery. Components of this pathway regulate intracellular AMP levels and their abrogation leads to gains in ATP and cold tolerance across disparate cell types8–10; however, genetic analyses found no significant divergence within this pathway in glacier ice worms.
To expand our investigation, we focused here on the F1Fo-ATP synthase complex, the primary source of ATP production under aerobic conditions 11 and a primary target of mitochondrial disease. 12 Gene sequence data, encoding 11 core subunits of ATP synthase, were extracted from the ice worm, 4 closely related congener species and 2 con-familial worms. 13 Comparative analyses revealed an unprecedented ~50 bp (base pairs), 3′-extension of the mitochondrial-encoded atp6 gene in ice worms, corresponding to a polar carboxy-terminal tail of ~18 amino acids. ATP6, a membrane-intrinsic component of the Fo domain, plays a key role in formation of the proton half-channels that provide H+ translocation across the inner mitochondrial membrane driving ATP production.14–16 Considering the functional importance of the ATP6 subunit and strong evolutionary trend of mitochondrial genome compaction,17,18 an insertion of this magnitude is unlikely to be maintained in the genome without providing a significant fitness advantage.
Materials and Methods
Specimens
Mesenchytraeus specimens were collected from field sites in Alaska (M solifugus-Byron), Washington (Mesenchytraeus gelidus, Mesenchytraeus hydrius), California (Mesenchytraeus pedatus), and British Columbia (Mesenchytraeus antaeus, M solifugus-Mariner Mountain) between 2011 and 2015, express-shipped live to Rutgers University, and maintained according to their endogenous habitat/temperature (eg, soil, refrigerated). Enchytraeus crypticus species were a laboratory-maintained stock and M solifugus-Davidson species were ethanol-preserved samples (see Lang et al 13 for details about collection sites).
Nucleic acid processing and polymerase chain reaction
For verification of RNA-Seq data, genomic DNA extractions were performed using E.Z.N.A. Tissue DNA Isolation Kit (Omega Bio-tek, Inc., Norcross, GA, USA) following the tissue protocol. Multiple sequence alignments (MEGA version 6; Tamura et al 19 ) of full-length atp6 transcripts derived from RNA-Seq data were used to design primers. A degenerate forward primer (ATGATACCWGATATTTTYTCMTCATTYGAYCC) was paired with species-specific reverse primers (Figure 1A, highlighted). No sequence data downstream of atp6 were available or retrievable to enable amplification from outside the atp6 coding region. Polymerase chain reaction (PCR) reactions contained 34.5 µL water, 200 µM dNTPs, 600 nM primers, 10 µL of 5× Phusion HF buffer, 1 U Phusion Hot Start II DNA Polymerase (Thermo Scientific, Waltham, MA, USA), and 50 to 200 ng total genomic DNA. Reaction conditions were as follows: denaturation at 98°C for 30°, followed by 35 cycles of 98°C for 10 seconds, 47 to 60°C for 20 seconds, and 72°C for 0.5 to 2 minutes; final extensions were 5 minutes at 72°C. Single-band PCR products were purified using DNA Clean and Concentrator (Zymo Research, Irvine, CA, USA); multiband products were electrophoresed on 0.8% agarose gels and desired bands excised. Gel-excised DNA was purified using a Zymoclean Gel DNA Recovery Kit (Zymo Research) according to the manufacturer’s protocol. Fragments were cloned into pMiniT vectors (New England Biolabs, Ipswich, MA, USA), transformed into NEB 10-beta chemically competent Escherichia coli, and 3 to 5 clones/fragment/species were selected for purification using Zyppy Plasmid Miniprep (Zymo Research). Transcriptomes and RNA-Seq data were generated and processed as described in Lang et al. 13

Verification of Mesenchytraeus solifugus atp6 3′-extension in different ice worm populations. (A) Full-length atp6 sequence was polymerase chain reaction amplified from genomic DNA using an enchytraeid-specific forward primer (red) and the ice worm–specific 3′-extension primer and/or a species specific end primer (highlighted). (B) In addition to the Northern clade (Byron Glacier, MsByr) ice worms used for RNA-Seq analyses, 2 additional ice worm populations, Davidson Glacier (MsDav), considered ancestral, 20 and Mariner Mountain Glacier (MsMMt) were also examined (species color-coordinated to map). The 3′-extension primer amplified only ice worm samples, but the forward primer amplified all when paired with species-specific end primers. The translated protein alignment (C) shows the conserved block of amino acids (highlighted) found in all 3 ice worm populations, but not found in any other species. The 3′-extension is 6 amino acids shorter in the Davidson population. The alignment in (A) is shown with stop codons removed.
Mitochondrial sequence alignment and annotation
DNA sequence chromatograms were evaluated and trimmed using Chromas 2.4.3 (Technelysium, South Brisbane, QLD, Australia). Overlapping sequence fragments were aligned in MEGA version 6 and assembled into consensus sequence contigs using Mesquite v.3.04.19,21 Protein-coding genes were identified using National Center for Biotechnology Information’s (NCBI) BLASTx and BLASTn algorithms. Extracted transcripts were subjected to GenBank searches to verify gene identity, translated to ensure that they were not pseudogenes, and aligned to each other to assess relatedness. Extracts with questionable or incomplete sequence were re-BLAST using genus-specific queries and/or subjected to raw read mapping to rule out assembly error. In effort to determine the origin of the atp6 extension, the translated protein fragment corresponding to the last 18 amino acids was used as a query for local BLAST searches against our RNA-Seq databases as well as NCBI’s nonredundant protein sequence database.
Results
Computational alignments of transcriptome-derived F1Fo-ATP synthase subunits from glacier ice worms and congener species identified a novel, ice worm–specific sequence extending the 3′ end of atp6 by ~50 bp. Raw read mapping showed this sequence was present in >100 raw reads of 2 independent ice worm transcriptomes (and absent in all other transcriptomes) and therefore not an assembly error. To confirm its authenticity, PCR amplifications of full-length atp6 were performed from genomic DNA templates using a forward primer designed to amplify all considered species and an ice worm extension–specific reverse primer (Figure 1A). In addition to the 6 considered species, 2 additional ice worms from widely separated populations (Davidson Glacier, Mariner Mountain Glacier) were evaluated to determine whether the atp6 extension was common to ice worms or an isolated population anomaly (Figure 1B). Robust amplification occurred in all ice worm samples but not in other worms. Mariner Mountain ice worms displayed a 3′-end extension of the same length as the Byron Glacier population (equivalent to 18 amino acids), differing only in a single amino acid (V/P), whereas Davidson worms, members of the putative ancestral clade, 20 displayed a truncated version of the extension—the final 13 amino acids, with a single conserved change (S/A).
The translated amino acid sequence—(V/P)QDRSQAH(S/A)PGHAHSHGF (Figure 1C)—was not represented in any of our transcriptomes, nor was it analogous to any known metazoan sequence deposited in GenBank. Surprisingly, it shared strong sequence identity (⩾68%) to functionally diverse histidine-rich protein motifs from single-celled microbes (Figure 2).
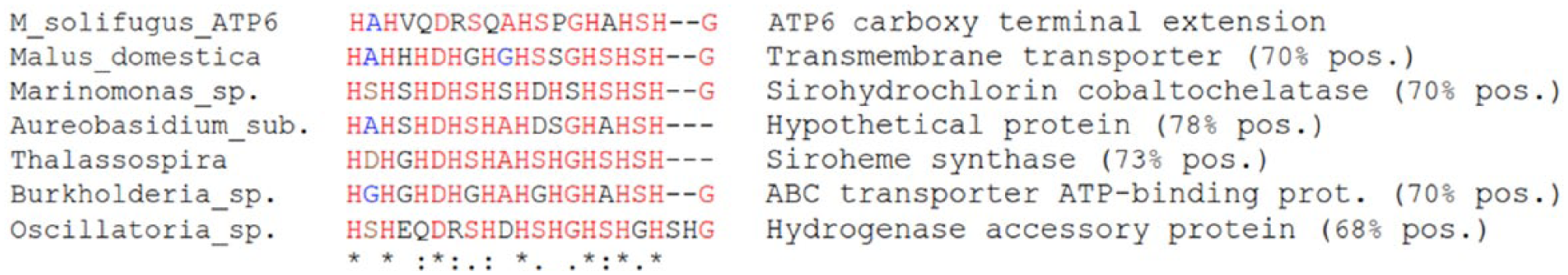
Protein alignment of NCBI nr-pBLAST results using Mesenchytraeus solifugus ATP6 C-extension as query. Histidine-rich domains are involved in numerous cellular processes, but membrane transport is a well-represented function. Malus domestica (apple), Marinomonas (marine bacteria), Aureobasidium subglaciale (glacier fungus), Thalassospira (marine bacterioplankton), Burkholderia (Betaproteobacteria), Oscillatoria (filamentous cyanobacteria).
Discussion
Confirmation of the ice worm atp6 ~50 bp 3′-extension has multiple implications. First, an insertion of this magnitude is unusual due to the strong evolutionary trend in metazoans toward compaction of the mitochondrial genome17,18; thus, sequences are not maintained unless they offer some evolutionary advantage. Davidson and Byron Glacier ice worms diverged from each other during the mid-Miocene (5-10 million years ago 20 ), and the observation that both lineages maintained atp6 extensions indicates strong Darwinian selection.
Second, the structural and functional roles of the ATP6 subunit in the F1Fo-ATP synthase are not only critically important but are also highly sensitive to modification. Specifically, most of the human mitochondrial diseases resulting from ATP synthase dysfunction are a consequence of atp6 single-nucleotide polymorphisms. 22 Indeed, of the 30 currently described atp6 disease-associated mutations, 28 are SNPs and 2 are dinucleotide deletions in atp6 (MITOMAP 12 ). In addition, a single amino acid change in the atp6-equivalent “A” subunit of an extreme alkaliphilic Bacillus bacterium is necessary and sufficient to confer high-pH adaptation. 23 Thus, the comparatively large change of an additional 18 amino acids to ATP6 is likely to have a major impact on its functional activity.
Third, the protein domain encoded by the ice worm ATP6 extension is clearly not random, but rather contains an ordered arrangement of alternating histidine residues, an infrequently used amino acid which can function as both proton donor and acceptor within a physiological pH range. Its origin does not appear to be metazoan, based on GenBank searches, instead it shares striking similarity to microbial sequences (up to 78%), particularly a glacier-dwelling yeast, Aureobasidium subglaciale (see Figure 2). Horizontal gene transfer (HGT), once thought to only occur in bacteria, is now recognized as an ancient and continuing process across all domains of life.24,25 Furthermore, the most common sources of HGT in eukaryotes are endosymbionts and diet.26–28 Notably, glacier-obligate microorganisms are the primary food source for ice worms, 29 and recent work identified a novel bacterial species living as an endosymbiont in the ice worm gut. 30 Note also that organisms occupying extreme environments are enriched in foreign genes, and these play important roles in ecological adaptation.31,32 It follows that the ice worm atp6 extension was likely acquired by HGT from a single-celled dietary source, representing only the second mitochondrial transfer event reported in Metazoa.33,34
The putative function of the ice worm ATP6 extension remains unknown, but a number of microbial genes containing similar His-rich domains function in ion/membrane transport, and histidine residues play a well-described role in proton shuttling. 35 Due to the known structural position of the ATP6 carboxy terminal at the matrix edge of the ATP synthase proton channel, we propose that the ice worm ATP6 extension enhances ATP synthesis via His-dependent shuttling of protons away from the matrix-inner membrane interface so as to increase the local pH, thereby increasing H+ ion flow. Genetic manipulations are inherently challenging in mitochondria; nonetheless, our collective data provide a logical mechanism associated with elevated ATP levels in glacier ice worms, as well as a high potential target for treating energy-related, mitochondrial disease.
Footnotes
Acknowledgements
For assistance with RNA-Seq data generation and assembly, we thank K Halanych and members of his lab (P Brannock, K Kocot, and D Waits) at Auburn University and R Fernández Garcia at the Giribet Lab, Harvard University.
Funding:
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work is supported by ARRA NIH R15GM093685 and Busch grants to D.H.S.
Declaration of conflicting interests:
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
SAL conducted experiments, analyses, and contributed to writing the manuscript. DHS collected field specimens and contributed to writing the manuscript.
Data Accessibility
DNA sequences: GenBank accessions KU728746-KU728800.
Ethical Statement
No vertebrate animals were used in these experiments.