
Abstract
Objectives
Feline infectious peritonitis (FIP) is a high mortality infectious disease. Single nucleotide polymorphisms (SNPs) in the genes encoding interferon gamma (IFNG), tumour necrosis factor alpha (TNFA) and dendritic cell-specific intercellular adhesion molecule-grabbing non-integrin (DC-SIGN; CD209) have been associated with increased and decreased risk of developing FIP. This study was designed to determine whether these associations were present in a UK population of pedigree cats using samples from cats euthanased with a confirmed diagnosis (FIP, n = 22; non-FIP, n = 10) or clinically healthy cats over 11 years of age (n = 3).
Methods
DNA was extracted from tissue (n = 32) or blood (n = 3) and PCR performed for regions of IFNG, TNFA and CD209. PCR amplicons were sequenced, each SNP genotype was determined, and genotype/allele frequency for each SNP and FIP status were compared.
Results
No significant association was found between the genotype and FIP status for any SNP analysed. There was a trend for the heterozygous CT genotype at both IFNG g.401 and IFNG g.408 to be associated with FIP (P = 0.13), but this genotype was also found in a substantial proportion of non-FIP cats. There was also a trend for the heterozygous CT genotype at IFNG g.428 to be associated with FIP (P = 0.06), although most cats with FIP had the CC genotype at this locus. No associations were found between any allele at TNFA g.-421, CD209 g.1900, CD209 g.2276, CD209 g.2392 and CD209 g.2713 and FIP.
Conclusions and relevance
The use of the IFNG, TNFA and CD209 SNPs described to predict the risk of FIP cannot currently be recommended.
Introduction
Feline infectious peritonitis (FIP) is a high mortality infectious disease of cats caused by feline coronavirus (FCoV). The prevalence of infection with FCoV can reach 90% in multi-cat households, 1 but FIP develops in <5% of infected cats. 2 Feline genetic factors are suspected to play a role in the development of FIP, with studies from North America and Australia showing an increased risk of FIP in pedigree over non-pedigree cats.3–6 In addition, some breeds,3,4,6 and even bloodlines within a breed, 7 appear to have a higher risk of developing FIP than others. There also appears to be some variation in breed predisposition between continents.3,4,6 Abyssinians and Rexes were found to be at high risk and Persians at low risk in both the USA and Australia. However, Burmese were found to be at high risk in Australia and at low risk in the USA, while Himalayans (colour-point Persians) were found to be at high risk in the USA and at low risk in Australia. A German study found no association between breed and predisposition to FIP, but the number of pedigree cats included was small and mixed-breed and domestic shorthair cats were amalgamated in one group. 8
During the pathogenesis of FIP, following infection of enterocytes, FCoVs infect monocytes and macrophages, where they replicate and are systemically spread. 9 Entry of FCoVs into some feline cells is mediated by interactions between the viral spike glycoprotein and host receptor proteins, some of which have yet to be elucidated. 10 Dendritic cell-specific intercellular adhesion molecule-grabbing non-integrin (DC-SIGN), a mannose-binding lectin, has been shown to be a key co-receptor during monocyte entry of both serotype I and II FCoVs. 11 An aberrant immune response to the presence of FCoV in macrophages and monocytes results in the development of vasculitis and tissue granulomas, which, in turn, leads to the body cavity effusions and mass lesions that characterise this disease. 12
Studies have suggested that a decreased risk of developing FIP is associated with a strong T-helper 1 cell-mediated immunity, with higher concentrations of interferon-gamma (IFN-γ) mRNA or protein in the blood of healthy FCoV-infected cats than cats with FIP.13,14 Interestingly, one of these studies also showed high levels of IFN-γ in the abdominal effusion of cats with FIP, despite having low levels in the blood, suggesting that cell-mediated immunity may also play a role in mediating pathogenesis. 14 A vaccination study indicated that susceptibility to FIP is also likely to involve dysregulation of the IFN-γ/tumour necrosis factor-alpha (TNF-α) response, with production of high IFN-γ and low TNF-α levels associated with a decreased risk, and production of low IFN-γ and high TNF-α levels associated with an increased risk. 15 Another study also showed that transcription of inflammatory cytokines and chemokines, including TNF-α and IFN-γ, was upregulated in the mesenteric lymph nodes of cats with FIP, consistent with inflammatory pathway activation. 16
Feline single nucleotide polymorphisms (SNPs) have been associated with the risk of developing FIP. One study that looked at the IFN-γ gene (IFNG) found the heterozygous genotype (CT) at both IFNG g.401 and IFNG g.408 to be associated with an increased risk of FIP. 17 These two SNPs appeared to be in complete linkage disequilibrium; that is, the genotypes at these two loci were identical in all cats examined. In addition, the presence of a T allele at IFNG g.428 was associated with a decreased risk of FIP. Another study that looked at the TNF-α gene (TNFA) found the presence of a T allele at TNFA g.-421 to be associated with a decreased risk of FIP. 18 In addition, when the DC-SIGN gene (CD209) was examined, the presence of an A allele at CD209 g.1900 or a T allele at CD209 g.2713 were reported to be associated with an increased risk of FIP, while the presence of a T allele at CD209 g.2276 or an A allele at CD209 g.2392 were found to be associated with a decreased risk of FIP. 18
The aim of the present study was to evaluate the utility of these SNPs in predicting the risk of FIP in a cohort of pedigree cats using samples from the Bristol FIP Biobank.
Materials and methods
Animals and samples
The cats in the study comprised those of known pedigree within the Bristol FIP Biobank (n = 32), for which a definitive diagnosis of either FIP (n = 22) or a disease other than FIP (n = 10) were available, and elderly (>11 years), staff-owned pedigree cats (n = 3) that were considered to be healthy based on history, clinical examination and blood testing. The FIP cats comprised British Shorthair (n = 6), Ragdoll (n = 6), Birman (n = 3), Maine Coon (n = 2), Siamese (n = 2) and one each of Abyssinian, British Blue and Burmese. The non-FIP cats comprised Bengal (n = 3), Ragdoll (n = 2) and one each of Birman, British Shorthair, Devon Rex, Havana, Maine Coon, Persian-cross, Siamese and Tiffanie.
For the cats from the Bristol FIP Biobank, tissue samples were collected at post-mortem examination. Some of these cats had been included in earlier studies.16,19–21 Formalin-fixed tissue samples were subjected to histological examination, including immunohistochemistry for FCoV antigen to confirm FIP, and categorised as FIP or non-FIP, as previously described. 19 Tissue samples collected into RNAlater (Life Technologies, ThermoFisher Scientific), faeces and body cavity fluid were stored at −80°C prior to processing. For the staff-owned cats, excess blood from samples collected for routine health screening was stored at −80°C prior to processing.
DNA extraction, amplification and sequencing
Feline genomic DNA was extracted from stored samples as previously described for total nucleic acids. 19 Feline genomic DNA was extracted from blood (100 µl) using a Chemagic 360 automated platform (Perkin-Elmer) in combination with the Chemagic body fluids nucleic acid kit (Perkin-Elmer) and eluted in elution buffer (100 µl).
Tissue, faeces and body cavity fluid samples from the Bristol FIP Biobank were subjected to FCoV quantitative RT-PCR, with results previously reported elsewhere. 19
PCR to amplify gene fragments containing the target SNPs was performed using 2 × GoTaq Master Mix (Promega), 200 nM forward and reverse amplification primers (see Table 1) and 5 μl of DNA in a total volume of 25 μl. Thermal cycling was performed in a PTC-200 DNA Engine (MJ Research) with the following thermal profile for IFNG and TNFA: 95°C for 2 mins followed by 40 cycles of 95°C for 20 s, 60°C for 20 s and 72°C for 20 s; and the following thermal profile for CD209: 95°C for 2 mins followed by 35 cycles of 95°C for 20 s, 64°C for 20 s and 72°C for 60 s. Amplicons were visualised by agarose gel (1% w/v) electrophoresis, using ethidium bromide stain and viewed using a GelDoc-It Imaging System (UVP).
Primers used in PCR amplification (A) and sequencing (S) for each gene

Amplicons (IFNG 559 base pairs [bp; primer set 1] or 799 bp [primer set 2], TNFA 498 bp, CD209 1095 bp) were purified (NucleoSpin Extract II; Macherey-Nagel) according to the manufacturer’s instructions and sequenced (DNA Sequencing & Services, University of Dundee, Dundee, UK; www.dnaseq.co.uk) (see Table 1 for primer sequences). The IFNG amplicon was sequenced using the forward and reverse amplification primers. The TNFA amplicon was sequenced using a separate forward sequencing primer and the reverse amplification primer. The CD209 amplicon was sequenced using separate forward sequencing primers and the reverse amplification primer. The derived sequences were assembled using MacVector v15.5.4. As allele dropout was seen in some cats for IFNG using primer set 1, IFNG primer set 2 was used in these cats.
Data analysis
SNPs were described relative to their genomic position from the ‘A’ of the start codon of the relevant gene. The association between each genotype at each position and FIP status (FIP vs non-FIP; non-FIP comprised cats that were euthanased for diseases other than FIP, and the healthy, alive cats) was analysed using Fisher’s exact test with the Freeman–Halton extension. The association between each allele frequency at each position and FIP status was analysed using Fisher’s exact test. Analyses were performed in XLSTAT v2017.6 (Addinsoft). A P value of ⩽0.05 was considered statistically significant.
Results
Data for each sample (including FIP status and genotype for each SNP) are presented in the table in the supplementary material. All cats with FIP had FCoV detectible in one or more tissue samples by PCR, and, by definition, at least one tissue was positive by immunohistochemistry for FCoV antigen within inflammatory lesions. Three of the deceased non-FIP cats were shedding FCoV in faeces at the time of death and two had FCoV detected by PCR in tissue samples collected post mortem (immunohistochemistry was negative for FCoV antigen in these tissues); the remaining non-FIP cats were negative for FCoV by PCR in all samples available for testing. FCoV serology was not available for cats in the Bristol FIP Biobank. One of the living cats was reported to be seronegative for FCoV at 12 years of age.
IFNG SNPs
The IFNG g.401 and IFNG g.408 SNPs were in complete linkage disequilibrium, with identical genotypes at these loci in all 35 cats. Table 2 shows the number and percentage of cats with each genotype at these loci. No genotype was found to be associated with FIP at either locus (P = 0.13), although the percentage of cats with the heterozygous CT genotype at both loci was 2.5 times greater in FIP cats (n = 13/22 [59.1%]) than in non-FIP cats (n = 3/13 [23.1%]). Neither the C nor T alleles at either locus were associated with FIP status.
Interferon-gamma gene (IFNG) genotype and allele frequency results for non-feline infectious peritonitis (non-FIP) and FIP cats included in the study

P value indicates the likelihood that a genotype or allele is associated with FIP status as determined by the Fisher’s exact test (with the Freeman–Halton extension as required)
SNP = single nucleotide polymorphism
All 13 non-FIP cats had genotype CC at IFNG g.428, whereas only 16/22 (72.7%) FIP cats had this genotype. No genotype was found to be associated with FIP status at this locus (P = 0.06), although no cats in the non-FIP group had the heterozygous CT genotype and no cats in either group had the TT genotype. Neither the C nor T alleles at either locus were associated with FIP status.
SNPs were also found (see the table in the supplementary material) at IFNG g.230, IFNG g.253, IFNG g.308, IFNG g.333, IFNG g.468, IFNG g.523 and IFNG g.524, but none were associated with FIP status.
TNFA SNPs
No genotype at TNFA g.-421 was associated with FIP status, and neither were the C nor T alleles (Table 3).
Tumour necrosis factor-alpha gene (TNFA) genotype and allele frequency results for non-feline infectious peritonitis (non-FIP) and FIP cats included in the study

P value indicates the likelihood that a genotype or allele is associated with FIP status as determined by the Fisher’s exact test (with the Freeman–Halton extension as required)
SNP = single nucleotide polymorphism
CD209 SNPs
All 35 cats had genotype GG at locus CD209 g.1900. At locus CD209 g.2276, 34 cats had genotype CC and one FIP cat had genotype CT. At locus CD209 g.2392 (Table 4), the majority of FIP and non-FIP cats had genotype GG, and there was no association between genotype, or between G or A alleles, and FIP status. At locus CD209 g.2713 (Table 4), the majority of FIP and non-FIP cats had genotype CC, and there was no association between genotype, or between C or T alleles, and FIP status. A further SNP was found at CD209 g.2718 (see the table in the supplementary material) but was not associated with FIP status.
Dendritic cell-specific intercellular adhesion molecule-grabbing non-integrin gene (CD209) genotype and allele frequency results for non-feline infectious peritonitis (non-FIP) and FIP cats included in the study
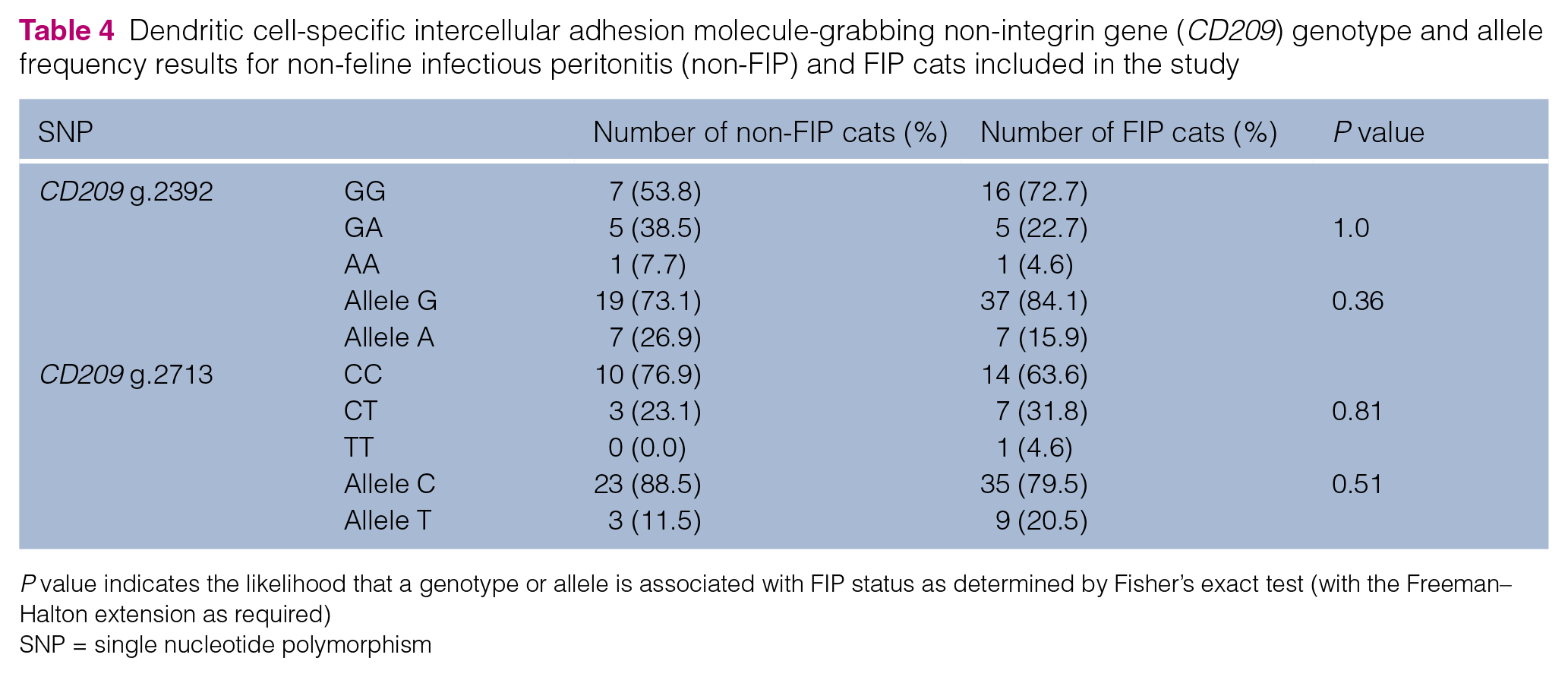
P value indicates the likelihood that a genotype or allele is associated with FIP status as determined by Fisher’s exact test (with the Freeman–Halton extension as required)
SNP = single nucleotide polymorphism
Discussion
An SNP describes nucleotide variation at a single point within the genome. SNPs within protein coding regions of a gene (ie, exons) may change the translated amino acid sequence or result in premature termination of translation and protein truncation. SNPs in non-coding regions (eg, introns) may increase or decrease transcription factor binding or change gene splicing and result in altered protein levels or alternative isoforms.
SNPs in genes such as IFNG, TNFA and CD209 can significantly change the amount and function of these important immunomodulatory proteins and potentially influence disease risk.22–25 A number of SNPs in these genes have been reported to be associated with an increased or decreased likelihood of FIP.17,18 These comprise: three SNPs in non-coding regions of IFNG (intron 1 IFNG g.401, g.408 and IFNG g.428); one SNP within the non-coding gene promotor region of TNFA (TNFA g.-421); one SNP within the extracellular domain of the protein (CD209 g.1900); and three SNPs in non-coding regions (intron 6 CD209 g.2276 and CD209 g.2392; intron 7 CD209 g.2713). Of these nine SNPs, only one (CD209 g.1900) is predicted to affect protein structure/function by substituting Trp128 for a stop codon. The previously reported intronic SNPs found associated with disease status may be in linkage disequilibrium with other loci that are yet to be discovered, and it is these loci that confer disease susceptibility or resistance. Our study aimed to characterise these previously reported SNPs in a pedigree population of 35 UK cats, and compare genotype and SNP allele frequency to FIP status.
Our results supported the previously reported finding of complete linkage disequilibrium between IFNG g.401 and IFNG g.408, 17 as this was present in all cats tested. However, a significant association between the heterozygous CT genotypes at IFNG g.401 and IFNG g.408 and FIP status was not found in this study, although a previous study found significance, with 62.1% (n = 18/29) of FIP cats having the CT genotypes vs 31.7% (n = 26/82) of non-FIP cats (Fisher’s exact test, P = 0.004). 17 In our study there was, nonetheless, a trend for the CT genotypes to be more common in FIP cats (59.1%) than in non-FIP cats (23.1%). Our study was limited by the number of pedigree cats that had been euthanased for a non-FIP cause, and it is possible that, with the greater power of a larger study, a significant association of the CT genotypes with FIP could have been found. However, it should be noted that in both this and the previously published study, 17 a significant number of non-FIP cats had the CT genotypes and, therefore, absence of the CT genotypes was not a good predictor that cats will not develop FIP. Hence, at this time, the selection of breeding cats based on the presence or absence of the CT genotypes at IFNG g.401 and IFNG g.408 cannot be recommended.
There was also a trend for the heterozygous CT genotype at IFNG g.428 to be associated with FIP (P = 0.06), although most cats with FIP and all of the non-FIP cats had the CC genotype at this locus. This is in contrast to previously reported findings, which showed the T allele at IFNG g.428 to be more prevalent in the non-FIP group. 17 Indeed, in our study the T allele at IFNG g.428 was only found in FIP-affected cats (n = 6/22). The difference in association between the two studies might be explained by different genetic populations.
Our results found no evidence of any association between FIP risk and SNPs TNFA g.-421, CD209 g.1900, CD209 g.2276, CD209 g.2392 and CD209 g.2713. These SNPs were previously reported to confer either an increased (CD209 g.1900 G>A, odds ratio [OR] 3.7; CD209 g.2713 C>T, OR 1.75) or a decreased (TNFA g.-421 C>T OR 3.9; CD209 g.2276 C>T, OR reported as not available; CD209 g.2392 G>A, OR 2.6) risk of developing FIP. 18 In addition, these authors described a G to A substitution at CD209 g.1900 that was not seen in our study population. 18
We are aware that the current study has several limitations; these comprise the number of cats included, the ‘potential’ for the non-FIP cats to have developed FIP at a later stage and the possibility for genetic variation between breeds. However, all three of these limitations were also present in the other studies.17,18
We had a total of 35 cats, of which 13 were classified as non-FIP and 22 as FIP. Using the genotype frequency determined for the population described in our study, at least 28 cats would be required in each group to achieve statistical significance between the groups. However, we were limited by the number of pedigree cat samples that were available in the Bristol FIP Biobank.
The possibility that the deceased cats in the non-FIP group could have gone on to develop FIP, were they not to have developed the alternative disease for which they were euthanased, cannot be ruled out. The non-FIP cats used in previous studies were all <3 years of age at the time of enrolment with evidence of FCoV infection (positive by FCoV PCR on one or more of blood, nasal/oral/conjunctival/rectal swabs and/or faeces),17,18 and were alive at least 2 years after recruitment in one of these studies. 17 However, in the current study, the non-FIP cats had either been euthanased for reasons other than FIP, with a median age of 5.8 years (range 4 months to 13.8 years), or are alive at the time of writing (⩾12.4 years). Data regarding prior exposure, infection and shedding status were available for a proportion of the non-FIP cats (see the table in the supplementary material), but not for all, so did not provide any meaningful results. One of the living cats with CT genotype at both IFNG g.401 and IFNG g.408 had a negative FCoV serology result; however, it had resided in a single-cat household for nearly its entire life after being obtained from a multi-cat household as a kitten, such that early prior exposure could not be ruled out. FCoV serology was not available for the other two living cats.
It cannot be ruled out that some pedigree breeds may be fixed for a single allele for a specific SNP, potentially creating bias; however, for the most frequently represented breeds (British Shorthair, Ragdoll, Birman), different genotypes were present at the target loci. It was not possible to statistically compare results within individual breeds owing to the small numbers in each group. In the study by Hsieh and Chueh, 17 approximately half of the cats were reported to be of known pedigree, or pedigree cross; however, pedigree breed and SNP status data, including any relationship between the two, were not reported. Future studies could focus on individual breeds for which the risk of developing FIP appears to be greater (eg, Abyssinians and Rexes), to exclude interbreed variation.
Conclusions
While not achieving statistical significance, the results of our study support a possible weak positive association between the CT genotypes at IFNG g.401 and IFNG g.408 and the risk of developing FIP. However, as the CT genotypes at these positions were present in a significant proportion of non-FIP cats (n = 3/13) and absent in a significant proportion of FIP cats (n = 9/22), a genetic test based on these two SNPs alone to predict the risk of developing FIP, or to guide breeding programmes to reduce the risk of FIP, cannot be recommended at this time.
Supplemental Material
Table 1
For each sample, including FIP status, breed, age at sampling, cause of death (or alive at time of writing), and genotype for each single nucleotide polymorphism tested.
Footnotes
Acknowledgements
The authors thank the veterinary practices, cat breeders and rescue centres that helped in the acquisition of samples used in this study. We thank our colleagues, current and past, at the Feline Centre and Veterinary Pathology Unit, Langford Vets, University of Bristol, who assisted in obtaining post-mortem samples. We thank members of the Histology Laboratory, Veterinary Laboratory Services, Institute of Veterinary Science, University of Liverpool and Institute of Veterinary Pathology, Vetsuisse Faculty, University of Zurich, for their technical assistance.
Supplementary material
The following file is available online: Table: for each sample, including FIP status, breed, age at sampling, cause of death (or alive at time of writing), and genotype for each single nucleotide polymorphism tested.
Conflict of interest
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This study was supported by a Langford Vets Clinical Research Grant to HK-D, ST and CRH, for which the authors are very grateful.
Ethical approval
The project has been approved under ethical review by the University of Bristol Animal Welfare and Ethical Review Board (VIN/14/013; VIN/16/020).
Informed consent
This was a retrospective study of samples previously collected with full informed consent from owners who agreed that samples could be used for research purposes.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.