
Abstract
Immunological memory is a key feature of adaptive immunity. It provides the organism with long-lived and robust protection against infection. The important question is whether cyclophosphamide (CP), as immunosuppressive agent used in cancer therapy and in some autoimmune diseases, may act on the memory T-cell population. We investigated the effect of CP on the percentage of central memory T cells (TCM) and effector memory T cells (TEM) in the mouse model of CP-induced immunosuppression (8-10-week-old male C57BL/6 mice CP treated for 7 days at the daily dose of 50 μg/g body weight [bw], manifested the best immunosuppression status, as compared to lower doses of CP: 10 or 20 μg/g bw). The CP induced a significant decrease in the percentage of CD8+ (TCM), compared to nonimmunosuppressed mice. This effect was not observed in the case of CD4+ TCM population. The percentage of gated TEM with CD4 and CD8 phenotype was significantly decreased in CP-treated mice, as compared to the control ones. Taken together, the above data indicate that CP-induced immunosuppression in mice leads to a reduction in the abundance of central memory cells possessing preferentially CD8+ phenotype as well as to a reduction in the percentage of effector memory cells (splenocytes both CD4+ and CD8+), compared to the cells from nonimmunosuppressed mice. These findings in mice described in this article may contribute to the understanding of the complexity of the immunological responses in humans and extend research on the impact of the CP model of immunosuppression in mice and memory T-cell populations.
Keywords
Introduction
Immunosuppression is defined as a transient or permanent dysfunction/damage of the mechanisms involved in cellular and/or immunity caused by different immunosuppressive agent that leads to reduction of the activation or efficacy of the immune system. Immunosuppression is often associated with the treatment of a broad range of autoimmune diseases, allergic conditions, cancer, infectious diseases (bacterial, viral, and fungal) and with the therapy of transplant patients. 1 –6
Cyclophosphamide (CP) is an alkylating agent and is usually administrated orally or intravenously and much less frequently intramuscularly or intraperitoneally. 7 –14 CP is hydrolyzed in the liver by cytochrome enzymes P450 to the active form 4-hydroxyclophosphamide, which can diffuse out into circulation and affects the target cells. In the cytosol of cell, through spontaneous β-elimination, acrolein and phosphoramide mustard evolve. These are the actual agents responsible for cytotoxic effects and DNA crosslinking. 9,15,16
Due to its cytotoxic and immunosuppressive effects, CP is commonly used against a broad spectrum of cancers, including lymphoma, chronic lymphocytic leukemia, soft tissue and osteogenic sarcoma, several solid tumors, and autoimmune diseases. 17 –20
Besides clinical applications, CP is also widely used in experimental research to induce immunodeficiency in animal models. 11,21,22 CP has been used to study numerous immune processes related to cancer development, transplant tolerance/rejection, and infections. 3,23 –29 Ultimately, the use of CP is critical for splenocyte functions; it affects cell levels in blood and lymphoid tissues and induces changes in the production of nonspecific (myeloperoxidase activity, reactive oxygen radicals, and antioxidant defenses) and specific immunity (antibodies). 30 –33
So far, the effect of CP on T-cell reactivity was described mainly in the context of regulatory T cells (Treg), which are a fundamental component of the immune response in tumors. In a study of patients with breast cancer who received CP, a significant decrease in Treg frequencies within CD4+ T cells up to 42 days following the initiation of treatment was observed. Two weeks later, Treg frequencies were back to pretreatment levels, indicating that upon prolonged exposure to CP, Treg might acquire resistance to depletion. 34 The mechanism of CP-mediated Treg depletion has been intensively studied in the past few years, and it was indicated that susceptibility of Treg cells to CP is multifactoral. 35 –37 Many phenotypic differences between Treg and effector T-cell population were noted. 38 However, there still exists a data gap concerning the effect of CP on the memory T-cell populations.
Immunological memory plays a fundamental role in an adaptive immunity. In the context of the model of immunosuppression with graft rejection, it has been noted that memory T cells are more resistant to immunosuppressive therapies than naive T cells due to the fact that they are independent of secondary lymphoid organs. 39 Moreover, memory T cells have been described to be resistant to regulation by CD4+CD25+FoxP3+ Treg cells. 40 The studies in in vitro and in vivo models have shown that memory CD4+ and CD8+ T cells exhibited lower susceptibility to inhibition of proliferation and cytokine production after exposure to alloantigen. It may have potential implications in the context of cell therapy and transplantation. 41,42 Two subsets of T cells, including TCM and TEM, which differentiate from naive T cells, differ concerning homing potentials and effectors functions and persist for years after immunization. 43 The studies indicated that memory T cells in humans and mice are similar concerning the fundamental mechanisms, thus knowledge acquired in mouse memory studies can be potentially translated to humans. 44,45
The aim of this study was to evaluate the influence of CP on memory T cells, in particular TCM and TEM subsets, in a mouse model of immunosuppression. A better understanding of the effect of CP on memory T-cell biology may provide new insights into the impact of the CP model of immunosuppression in mice and memory T-cell populations.
Materials and Methods
Animals
C57BL/6 mice (males, 5 weeks) were purchased from Charles River Laboratories (Sulzfeld, Germany). Upon arrival, all mice were housed in ventilated cages in environmentally controlled rooms (20°C-25°C, 50% ± 10% humidity, 12-hour/12-hour dark/light). All mice had ad libitum access to pelleted feed and filtered water. After at least a 3-week acclimatization, the mice were randomly allocated into different groups designated to be used for experiments. All procedures were approved by the Local Ethics Commission for Animal Experiments in Lodz (Poland).
CP Administration
The CP (Endoxan; Baxter Oncology GmbH, Halle, Westfalen, Germany) powder was reconstituted to a stock concentration of 10 mg/mL by addition of sterile 0.9% NaCl. The stock was then diluted so that equal injection volumes could be given by intraperitoneal injection to deliver a low, medium, or high dose (CP10, CP20, or CP50 µg/g body weight [bw], respectively) for 7 consecutive days. Control mice received the same volume of saline. Unless indicated below, mice were euthanized by intraperitoneal injection of pentobarbital sodium (Biowet, Puławy, Poland) 24 hours after the last dose of CP. Mice were observed during the course of the experiment and weighed at the start of the experiment and at the daily intervals thereafter till they were sacrificed. Body weight index was calculated from the weight of the body at the date of measurement versus at the start of experiment.
Leukocyte Index and Serum Humoral Immunity Parameters
At designated time points, 2 different blood analyses were performed: First, the leukocyte composition (leukocyte index) was analyzed, second selected humoral parameters (total serum protein, albumins, as well as α-, β-, and γ-globulin levels) were evaluated. In the first case, tail vein blood was recovered from each mouse, 46 collected into anticoagulant-coated tubes, and immediately sent to a veterinary laboratory (Center of Veterinary Analysis, Vetcomplex, Łódź, Poland). Leukocyte index was determined prior to the first CP dosing and on days 2, 4, and 7. In the second case, subsets of mice were euthanized on days 3 and 7 and blood was collected via heart puncture. Serum from each sample was prepared using standard protocols and stored at −20°C until analyses at the veterinary laboratory.
Lymphocyte Proliferation Assay
Mice given 0, 20, or 50 µg/g bw (control, CP20, and CP50, respectively) were euthanized and their spleens were collected at necropsy. Each spleen was aseptically homogenized using a glass tissue grinder (Sigma Aldrich, St Louis, Missouri), and generated cell suspension was centrifuged at 400 × g for 10 minutes at 4°C. The pellet was treated with erythrocyte lysis buffer (0.19 M NH4Cl) for 5 to 10 minutes at 4°C, centrifuged (as above), and washed twice with RPMI 1640 medium (Sigma Aldrich). Splenocytes were then suspended in complete RPMI 1640, that is, medium supplemented with 10% fetal calf serum (Biowest, Nuaillé, France) and antibiotics (100 U/mL penicillin, 100 µg/mL streptomycin; Biowest). The viability using trypan blue (Sigma Aldrich) exclusion was determined. Next, the cells were diluted in complete medium and dispensed into 96-well plates at 5 × 105 cells/well/100 µL. A 100-µL aliquot of phytohemagglutinin (final concentration in well = 2.5 µg/mL; Sigma Aldrich) was added to designated wells, while control wells received only the culture medium. The cells were then cultured at 37°C in a humidified 5% CO2 incubator for 48 hours and 100 μL [3H]-thymidine (2 μCi; Hartmann Analytic GmbH, Braunschweig, Germany) was added 18 hours before the end of the incubation. Next, the cells were harvested onto glass fiber filters and left to dry. The punch filters were then placed in vials and scintillation fluid (OptiPhase “SuperMix,” Wallac, Milton Keynes, UK) was added. The incorporation of [3H]-thymidine, reflecting proliferative activity, was then determined using a scintillation β counter (Wallac 1450 Microbeta Plus Liquid Scintillation Counter, Turku, Finland). Results were expressed in terms of total cpm/sample. All samples were analyzed in triplicate.
Isolation of Bone Marrow, Spleen, and Lymph Nodes
From the mice euthanized to obtain serum on days 3 and 7, the bone marrow from both femurs was isolated using standard protocols. 47 In parallel, the spleen, axillary, and inguinal lymph nodes were isolated and resuspended in RPMI medium, as above. Each cell suspension was centrifuged at 400 × g for 10 minutes at 4°C, the pellet was suspended in 1 mL medium, and total cell counts (and viability) were determined using a hemocytometer.
Identification of CD4+ and CD8+ TCM and TEM Populations by Flow Cytometry
In the first step, the unstimulated splenocytes from CP immunosuppressed and nonimmunosuppressed mice were isolated from the spleen, as described above. Next, the cells were fixed according to manufacturer’s instructions for the expression of CD4 and CD8 receptors with the following mAb: APC-Cy7-anti-CD4 and APC-anti CD8 (BD Biosciences, San Diego, USA) and for memory T-cell markers: APC-Cy7-anti CD4, APC-anti-CD8, V500-anti-CD44 (BD Biosciences), PECy7-anti-CD62L, and FITC-anti-CD127 (eBioscienes, San Diego, USA). An irrelevant isotype-matched mAb were used as control. Stained cells were incubated for 30 minutes at 4°C. The TCM were identified on the basis of the expression of CD62L+CD44+/− CD127+ CD4/CD8+ receptors and TEM had the following phenotype: CD62L−CD44+CD127+/− CD4/CD8+. Data were acquired and analyzed using the FACS LSRII (BD Biosciences) and FlowJo 7.2.2. software (TreeStar Inc. USA). A minimum of 10,000 events was collected. The percentage of positive cells (% gated) was determined.
Statistical Analysis
All data were expressed as means (standard deviation). All intergroup outcomes at fixed time points were compared for statistical significance using a 1-way analysis of variance and a Tukey Honest significant difference post hoc test or a nonparametric Kruskal-Wallis test. Intragroup, that is, as function of time, analyses were not performed. In all cases, significance was accepted at P < 0.05. All analyses were performed using STATISTICA 13.1 PL software (Statistica, Tulsa, Oklahoma).
Results
Effect of CP on the BW, Blood Leukocyte Composition, and Total Number of Cells in Lymphoid Organs
The analysis of bw in mice under CP treatment revealed that only the dose of 50 μg/g bw led to statistically significant progressive body mass loss compared to the control group, starting at day 5 of dosing (Figure 1A). Our study also showed that continuous treatment of mice with 50 μg/g bw of CP caused the decrease in the serum total protein level compared with the control (Table 1). This significant effect was observed after 7 days of the CP administration. Moreover, after 7 days of CP (50 μg/g bw) treatment, a significant decrease in the serum concentration of γ-globulins (33% vs control, P = 0.02) and β-globulins (29% vs control, P = 0.01) was observed. Our study also revealed that both CP20 and CP50 led to reduced numbers of cells in the spleen and inguinal lymph nodes at day 3 (Figure 1B). These changes were 22% from day 0 and 27% versus control at same time (spleen) and 64% from day 0 and 71% versus control at same time in the lymph nodes. Interestingly, in the bone marrow, the same 3-day treatments caused an increase in total cell numbers (21% from day 0 and 9% vs control at same time). However, after 7 days of CP50 μg/g bw regimen, a significant decline of cell numbers was evident (63% from day 3 and 59% vs control at same time). The mid-dose of CP (20 μg/g bw) did not cause any changes in the cell number of bone marrow versus control after 7 days of the treatment. The lowest dose of CP (10 μg/g bw) tested did not impact cell number measures at any time point. In this study, we also evaluated the leukocyte composition in peripheral blood (Figure 1C). The analysis disclosed that none of the tested CP doses induced significant changes in a percentage of monocytes in the peripheral blood. Interestingly, the dose of 50 µg CP/g bw, caused a noticeable increase in the proportion of lymphocytes (as compared to control) after 2 days of treatment; this level was maintained until day 7. In contrast, this CP50 resulted in a significant decrease in the percentages of granulocytes. CP10 and CP20 did not cause changes in the proportion of either lymphocytes or granulocytes.
Selected Indicators of Humoral Immunity in Mouse Serum.a
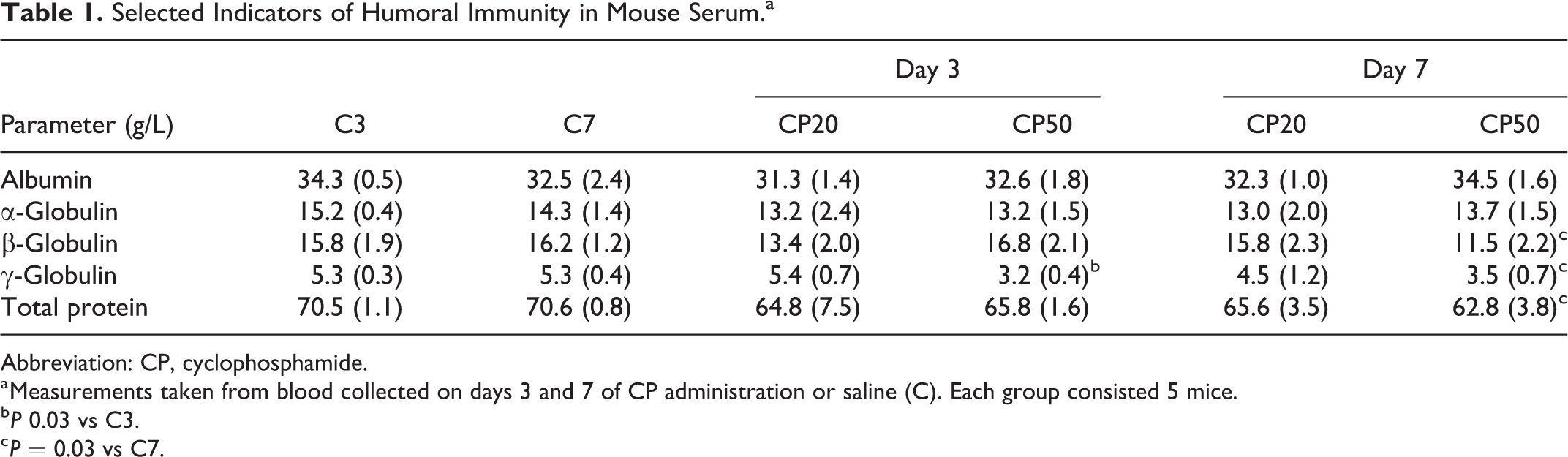
Abbreviation: CP, cyclophosphamide.
a Measurements taken from blood collected on days 3 and 7 of CP administration or saline (C). Each group consisted 5 mice.
b P 0.03 vs C3.
c P = 0.03 vs C7.
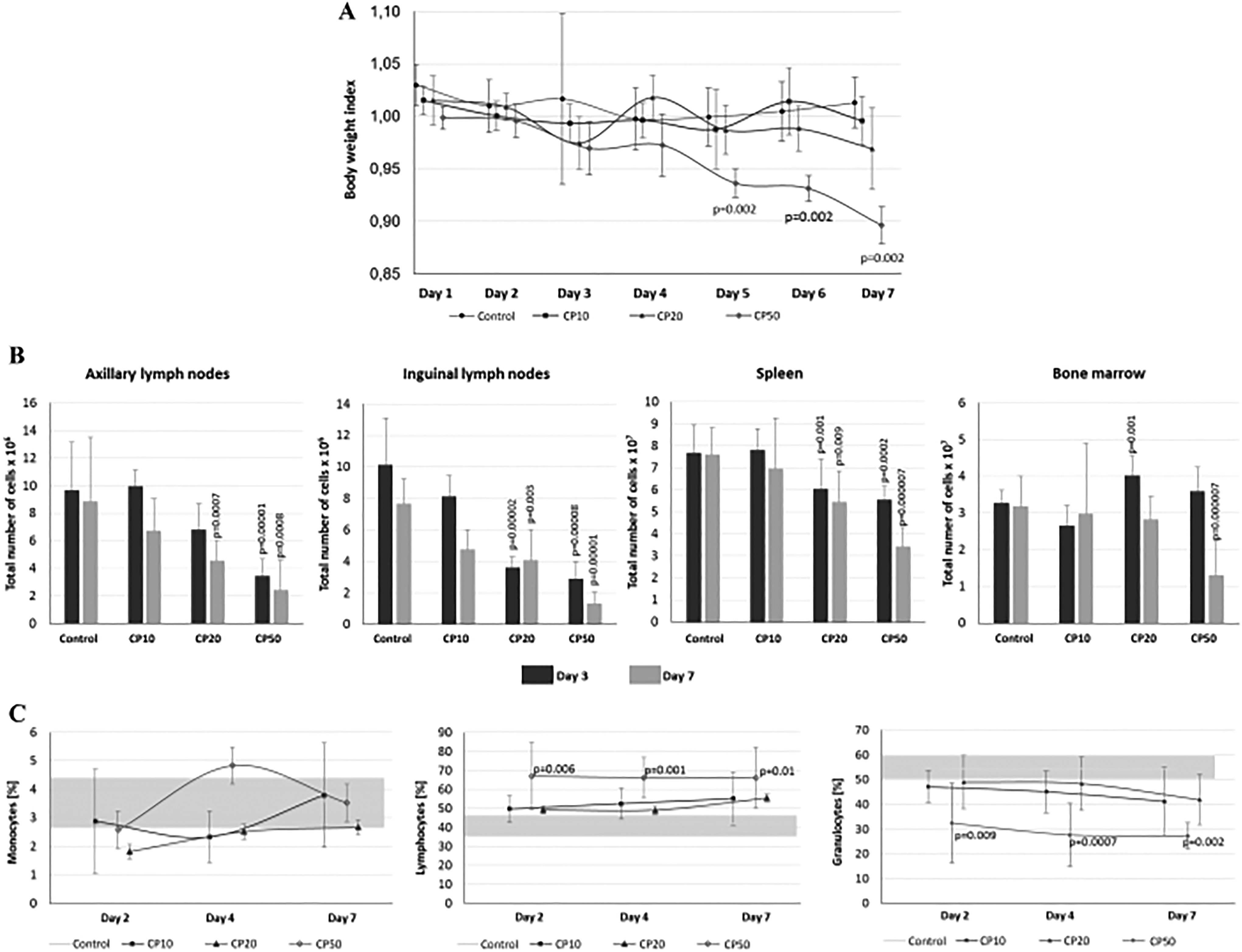
Effect of CP on (A) body weight, (B) total number of cells in the bone marrow, spleen, axillary, and inguinal lymph nodes, and (C) blood leukocyte levels (horizontal gray strip depicts cell number before CP injections). Data are expressed as means (SD; n = 5/group). P value versus control group is indicated. CP indicates cyclophosphamide; SD, standard deviation.
Effect of CP on Ex Vivo Proliferation of Splenocytes
Considering that CP10 was insufficient to cause the effective immunosuppression (as described above), the proliferation of splenocytes was evaluated only in the mice treated with CP20 and CP50. Only CP50 treatment induced a significant decrease in T-cell proliferation index as compared to control and to medium dose of CP (Figure 2).
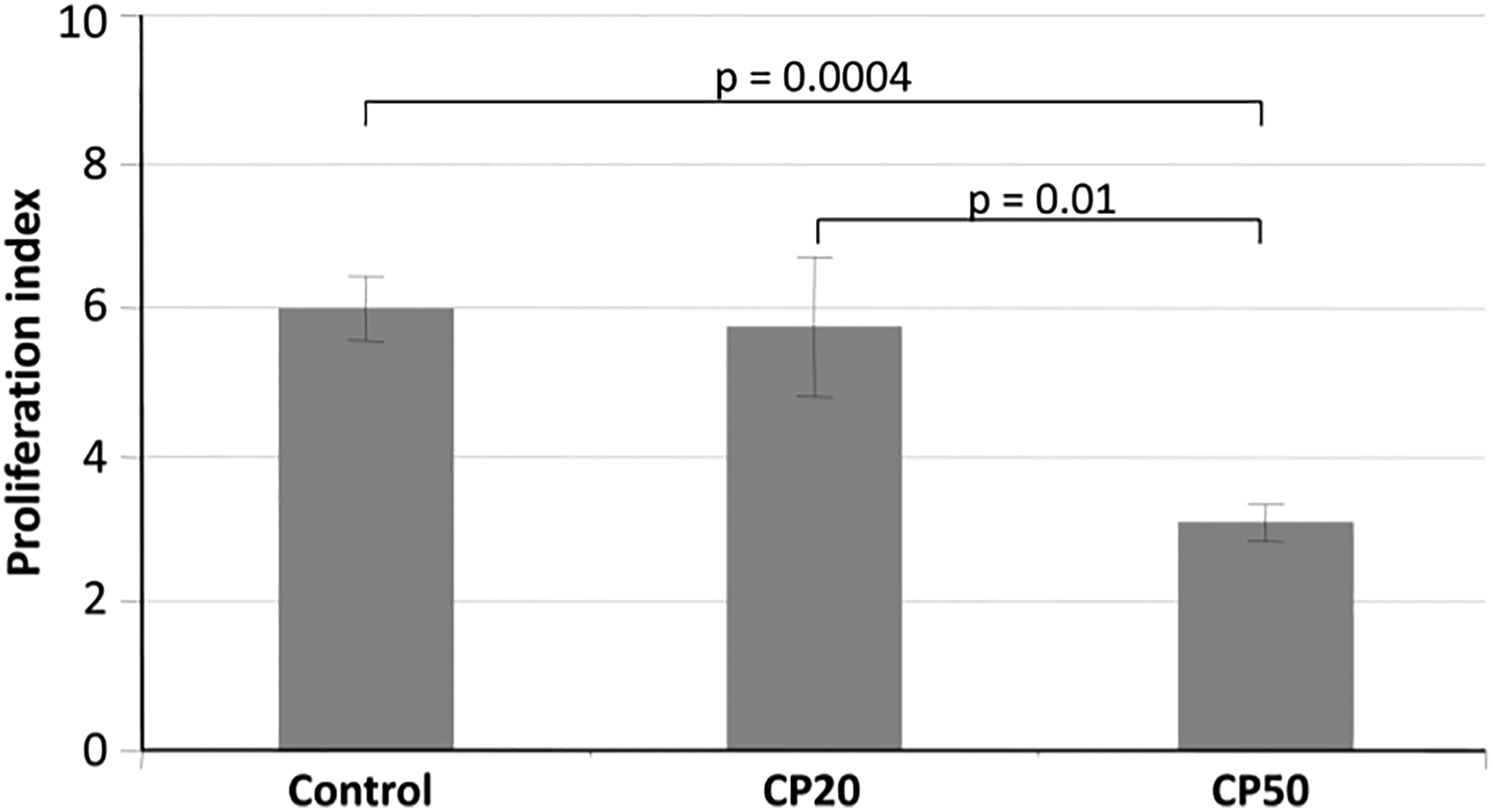
Effect of CP on ex vivo splenocyte proliferation. Data only from control, CP20, and CP50 are shown. Data shown are means (SD; n = 5/group/time point). P value versus control group is indicated. CP indicates cyclophosphamide; SD, standard deviation.
CP Impact on TCM and TEM Response
To investigate the effect of CP50 on memory T cells in mouse, the splenocytes from immunosuppressed mice were isolated and compared to control (cells obtained from nonimmunosuppressed mice). In the first step, splenocytes were gated based on the CD4 and CD8 surface markers expression. As indicated in Figure 3, the percentage of gated CD4+ and CD8+ T cells was similar in both immunosuppressed and control mice. Next, the memory phenotype of CD4+ and CD8+ splenocytes was analyzed. To differentiate between cells with the phenotype of TEM and TCM, we determined the expression of various T-cell surface markers including CD44, CD62L, and CD127 by flow cytometry. 48 In the CD8+ population, CP induced a significant decrease in the percentage of CD8+ TCM, compared to control group (Figure 4). This effect was not observed in the case of CD4+ TCM population. In parallel, a percentage of TEM was determined too. Interestingly, the percentage of gated TEM with CD4 and CD8 phenotype was significantly decreased in CP-treated mice as compared to control mice. Taken together, the above data suggest that CP-induced immunosuppression in mice leads to the generation of lower percentage of central memory cells possessing preferentially CD8+ phenotype and lower effector memory cells (splenocytes both CD4+ and CD8+), compared to the nonimmunosuppressed mice (Figure 3).
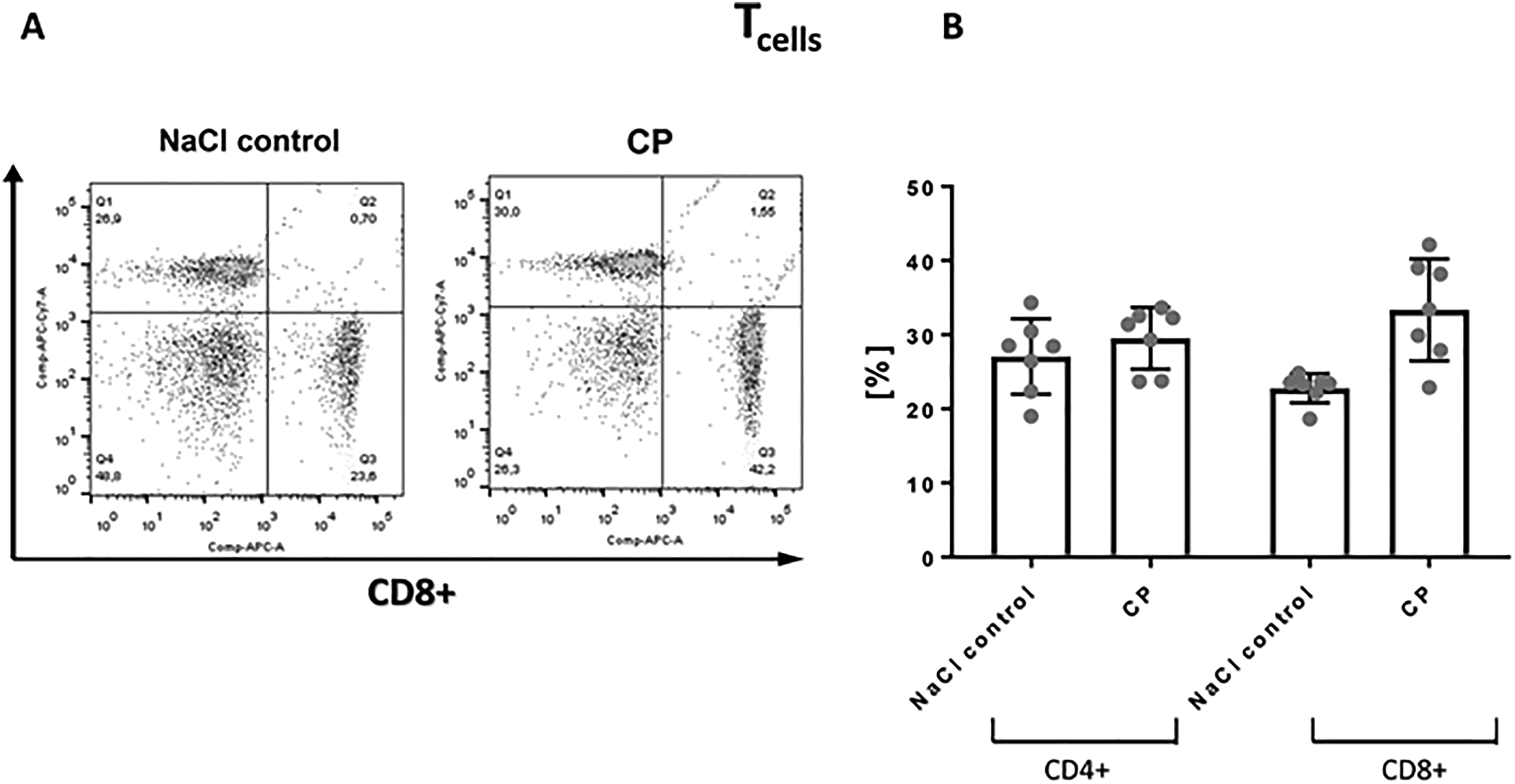
A, The gating strategy used to detect spleen CD4+ and CD8+ T-cell subsets. B, CD4+ and CD8+ surface expression on T cells from immunosupressed (CP) and nonimmunosupressed mice (NaCl control). One representative experiment of the 7 independent ones is shown. CP indicates cyclophosphamide.
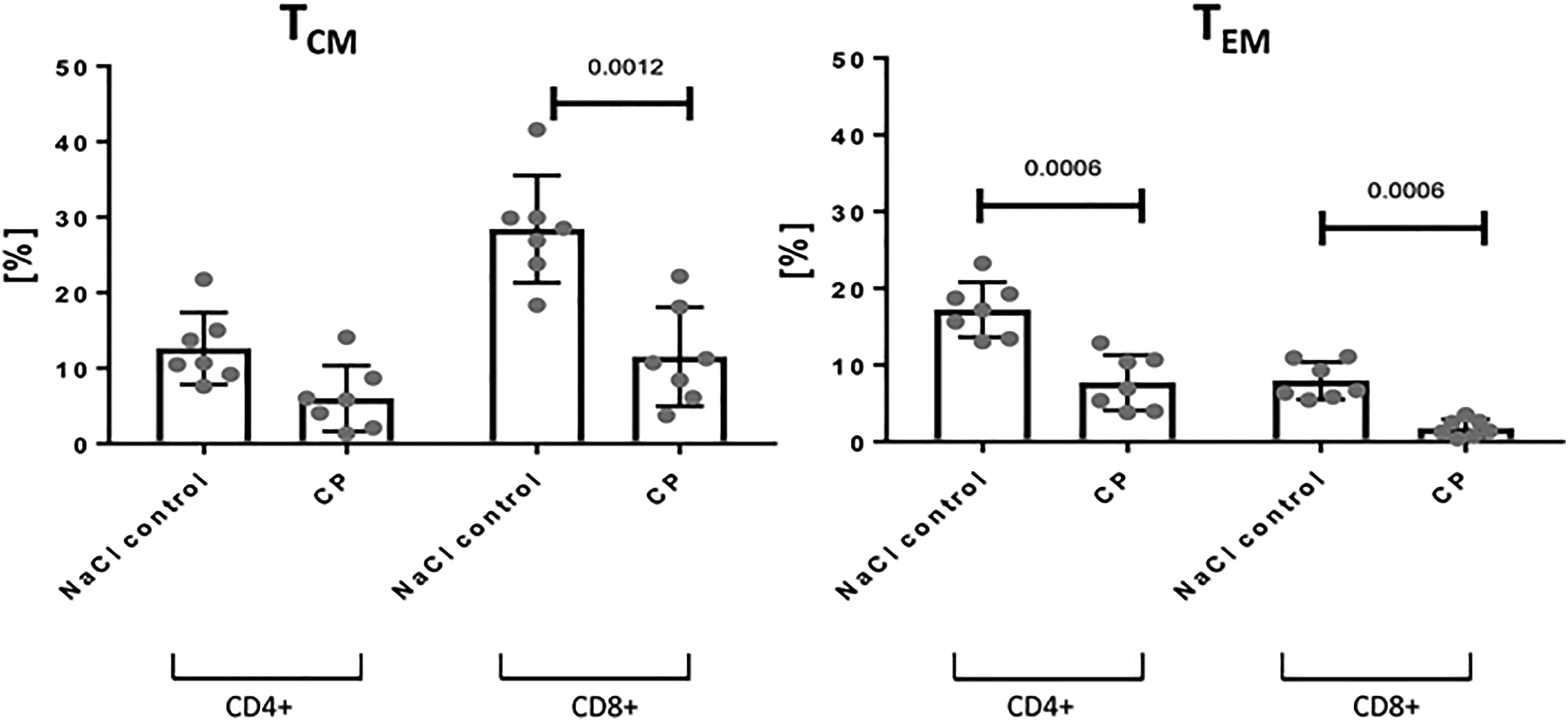
Effect of CP50 on central (TCM) and effector memory T cells (TEM) from immunosupressed (CP) and nonimmunosupressed mice (NaCl control). Data are expressed as means (SD; n = 7/group) of the percentage of positive cells with the (TCM) or (TEM) phenotype. P value versus control group is indicated. CP indicates cyclophosphamide; SD, standard deviation.
Discussion
Cyclophosphamide is widely used to treat various cancers. 49 –51 Since CP inhibits the immune system, it is commonly used to develop immunodeficiency in different animal models. The literature is replete with papers showing that CP induces marked effects on immune cells, depending on route of administration, 18,52 –55 dosage, timing of dose, and even the general type of animal model, 18,53,54,56 –59 or even animal strain. 60 To study whether CP impacts memory T cells in mice, in the first step, we established the appropriate animal model of CP-induced immunosuppression. Among 3 tested doses of CP (10, 20, and 50 μg/g bw), the most distinct effect was observed when 50 μg/g bw of CP was given to mice. The dose of 50 µg/g bw was also used in humans when the hematopoietic stem cell transplantation was necessary. According to Luznik et al, the high-dose CP regimen involving 2 doses before bone marrow infusion and the next 2 doses after transplantation was well tolerated and sufficient to prevent graft-versus-host disease (GVHD) development. 2 The effect of the same high dose of CP, given once or twice as a sole GVHD prophylaxis, was reported in the studies by Majzner et al. 61 We observed a significant decrease in total cell number in axillary and inguinal lymph nodes, spleen, and bone marrow (only day 7 for bone marrow), a decline in the serum levels of β- and γ-globulins, a reduction in the numbers of cellular components in blood and in splenocyte ex vivo proliferation activity. Furthermore, only the 50 μg/g bw dose for 7 days resulted in a decrease of bw of mice as compared to the mice in the control group. Feng et al noted similar effects when animals received intraperitoneally 50 μg CP/g bw once a day for 10 days. 52 Moreover, those researchers noticed that after stopping CP treatment, bws recovered quickly. This effect was also observed in our studies (data not shown). Kanno et al analyzed effects of CP on reproductive parameters in mice treated once a week (intraperitoneally) for 6 weeks with CP doses of 100, 150, 200, or 250 μg/g bw. They observed significant body mass variations among the mice. This bw alternation could be attributed to reduced appetite as a result of the CP toxicity. 54
The current study also showed that CP treatments led to changes in the composition of leukocytes in peripheral blood of the mice. These results were in accordance with Schiavoni et al. 62 They demonstrated that CP induced secretion of type I interferon (IFN) in vivo and then enhanced the expansion and long-term survival of T lymphocytes with a memory phenotype CD44highCD4(+) and CD44highCD8(+). Type I IFN is involved in the polarization toward Th1 immune response what augmented the generation and activity of CTL. According to Mattar et al, low dose of CP (10 μg/g bw) induced a considerable decrease in Th2 cytokines and significant enhancement in the production of Th1 cytokine simultaneously. 63
The present study also demonstrated a decrease in total cell number present in lymph nodes, spleen, and bone marrow of the CP-treated mice. This outcome supports findings by Barcew et al who demonstrated morphological destructive changes in the bone marrow and spleen of CP-treated mice. 64 Specifically, they observed significant decreases in cells on day 2 after intraperitoneal injection of 200 µg/g bw. A significant impact of CP on bone marrow, the primary site for hematopoiesis, could explain the time-related decreases in granulocyte levels in the other two sites as a result of the administration of 200 µg/g bw of CP.
In our study, we also showed that repeated treatments of mice with CP (50 μg/g bw) had an inhibitory effect on serum levels of β- and γ-globulins as well as total protein. The β-fraction is composed mostly of transferrin and β2-microglobulin, 65,66 and CP exerts toxic effect on hepatocytes/hepatic enzymes responsible for transferrin synthesis. 67,68 Both γ-globulins and β2-microglobulin are produced by lymphocytes whose activities are inhibited by CP. 22 As such, it is not surprising that levels of β- and γ-globulins, as well as total protein, were impacted by the high-dose CP regimen. The current results were thus in accordance with those of Salinas-Carmona et al who treated mice with CP and reported the significant decrease in serum β-globulins levels. 33
The results presented in this work showed that CP50 also led to a significant decrease in ex vivo T-cell proliferation, an outcome consistent with other studies in mice or in vitro in human lymphocytes. 69 –72 In contrast, Xu at al noticed proliferative activity of splenocytes (assessed using MTT [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide] colorimetric method) increased in CP-treated mice (150 μg/g bw, once week, 4 weeks). 73 Interestingly, in that study, serum levels of tumor necrosis factor-α, IFN-γ, and interleukin (IL)-2, IL-6, and IL-12 were significantly reduced in the CP-treated mice. While many of these cytokines are involved in upregulating inflammation, IL-2 helps to regulate T-cell proliferation. 63 Thus, a reduction in IL-2 due to CP should have resulted in less inhibition of T-cell proliferation in the hosts (or ex vivo).
The results of our work showed that the immune functions of CP-treated mice have been efficiently suppressed when CP was used at the concentration of 50 μg/g bw. Thus, this animal model was used for further evaluating the effect of CP on memory T cells.
Memory is the hallmark of the acquired immune system. In mouse models, it has been shown that in response to infections with pathogens and reinfections, the memory T cells are generated and the protective response is efficiently induced. Memory CD4+ T cells were found to mediate protective immune responses to infections caused by parasites, influenza, virus, and Mycobacterium tuberculosis, while mouse memory CD8+ T cell-mediated protection was shown in the lymphocytic choriomeningitidis virus model. 74 –77 The heterogeneity of the memory T-cell population was firstly described approximately 2 decades ago by Sallusto and colleagues indicating the existence of 2 subsets with distinct effector functions—TCM and TEM. 43 TCM subset is a group of cells circulating in the blood and in lymphoid tissues, such as the spleen, lymph nodes, or bone marrow, while TEM can circulate between lymphoid tissues and nonlymphoid tissues during the steady-state conditions. It has been noted that in vitro stimulation of naive T cells differentiates first to TCM and then to TEM depending on the strength and duration of T-cell receptor stimulation and the presence or not polarizing cytokines. 78 In mice, 2 populations of memory CD4 T cells were found to survive for months after immunization with antigen: One found in the lymph nodes producing IL-2; the other was found in nonlymphoid tissues and produced IFN-γ. 79 Moreover, following bacterial or viral infections, 2 populations of CD8 memory T cells were identified in nonlymphoid tissue. 80 Both CD4 and CD8 memory T cells persist as TCM and TEM in mice, and these 2 subsets differ in the expression of chemokine receptors, costimulatory molecules, and adhesion. 43
Although the presence of multifunctional memory T cells was established to be correlated with protective memory responses, there is a gap in understanding the protective response of memory T cells in immunosuppressed conditions. 48,81 The study by Schiavoni et al provided the first evidence that CP as an inducer of type I IFN-γ in vivo enhanced the number of T cells exhibiting the CD44hi memory phenotype. 62 However, the knowledge, how CP treatment affects different subtypes, especially TCM and TEM, remains unknown.
Our results indicate that short-termed (7 days) CP treatment in mice did not affect significantly the percentage of CD4+ and CD8+ T-cell populations. In contrast, when memory T-cell subpopulations possessing CD4 or CD8 receptor were analyzed, the significant decrease in TCM and then to TEM was observed compared to control mice. It has been shown that CP affects both CD4+ and CD8+ memory T-cell subpopulations located in lymphoid and nonlymphoid tissues. 82,83
As the quality of T-cell responses is essential for determining the disease outcome to various infections, better understanding of the functional capacity of TCM and TEM responses may contribute to the development of preventative and therapeutic strategies for infections, including the latent infections where the risk of the reactivation is dramatically increased in the immunocompromised patients.
Footnotes
Author Contribution
Włodarczyk, M. contributed to conception and design; contributed to acquisition, analysis, and interpretation of data; drafted manuscript; and critically revised manuscript; Ograczyk, E. contributed to acquisition of data and drafted manuscript; Kowalewicz-Kulbat, M. contributed to conception and design; contributed to acquisition, analysis, and interpretation of data; drafted manuscript; and critically revised manuscript; Druszczyńska, M. contributed to interpretation of data and critically revised manuscript; Rudnicka, W. contributed to conception and design and critically revised manuscript; Fol, M. contributed to conception and design; contributed to acquisition, analysis, and interpretation of data; drafted manuscript; and critically revised manuscript. All authors gave final approval and agree to be accountable for all aspects of work ensuring integrity and accuracy.
Authors’ Notes
At the time this study was conducted, MW, EO, MD, MK-K, and MF were employees of the University of Lodz, and WR was the retired employee of the University of Lodz.
Acknowledgments
This work was supported by the National Science Centre (Poland) under grant number 2013/11/B/NZ6/01304.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.