
Abstract
Aflatoxin B1 (AFB1), one of the most common mycotoxins found in human foods and animal feed, is principally hepatotoxic and hepatocarcinogenic. The aim of the present study was to explore the effect of AFB1 on messenger RNA (mRNA) expression of pregnane X receptor (PXR), constitutive androstane receptor (CAR), and aryl hydrocarbon receptor (AhR) and some of their target cytochromes using primary cultures of human hepatocytes. Our results showed that AFB1, at noncytotoxic increasing concentrations, caused a significant upregulation of cytochrome P 2B6 (CYP2B6), CYP3A5, and to a lesser extent CYP3A4 and CYP2C9. Pregnane X receptor and CAR mRNA expression increased in the 3 treated livers. Aflatoxin B1 was found also to induce an overexpression of CYP1A1 and CYP1A2 genes accompanied by an increase in AhR mRNA expression. These findings suggest that AFB1 could activate PXR, CAR, and AhR; however, further investigations are needed to confirm nuclear receptor activation by AFB1.
Introduction
Cytochrome P450s (CYP450s) are a large superfamily of heme-binding enzymes, which play an important role in the synthesis and metabolism of endogenous substrates and in the biotransformation of xenobiotics. 1,2 Besides inactivation of the foreign compounds, CYP-mediated biotransformation may result in the formation of toxic metabolites such as epoxides, quinines, and quinone imines. 3 Large differences frequently exist between humans and laboratory animals, 4 and interindividual variations are observed in the human response owing to genetic, physiopathological, and/or environmental factors. A number of xenobiotics are known to be inducers or inhibitors of CYPs. 5 ,6 Induction of CYPs was a receptor-mediated mechanism leading to an increase in gene transcription. 7 –9 Important nuclear receptors involved in the induction of CYP3A, CYP2B, and CYP1A subfamily forms comprise, respectively, the pregnane X receptor (PXR), the constitutive androstane receptor (CAR), and the aryl hydrocarbon receptor (AhR). Pregnane X receptor and CAR constitute 2 important members of the NR1I nuclear receptor family. They were originally defined as xenobiotic receptors and are master regulators of phase I and II drug-metabolizing enzymes as well as of drug uptake and export systems preventing the accumulation of toxic chemicals within the body. 10 –12
Aryl hydrocarbon receptor is a member of the basic helix-loop-helix/PER-aryl hydrocarbon nuclear translocator (ARNT)-SIM family of DNA-binding proteins. The AhR activation has been suggested to control the expression of xenobiotic-metabolizing enzymes, particularly, CYP1A1 and CYP1A2 enzymes. 13,14 In addition, AhR plays a significant role in carcinogenic effects of complex environmental mixtures of contaminants containing high levels of PAHs, such as extracts of airborne particulate matter. 15 Activation of AhR by various toxicants has been shown to induce perturbations of cell-signaling pathways regulating cell proliferation, differentiation, apoptosis immune response, and carcinogenesis. 16 –19 Given the diverse and important regulatory roles played by the AhR in basic biology and in toxicology, it is of great importance to investigate the effect of natural contaminants on AhR expression and activity, in order to predict likely drug interactions or chemically induced carcinogenesis.
Aflatoxin B1 (AFB1), the most common mycotoxin found in human food and animal feed, is the most prevalent and toxin of the aflatoxins. 20 Aflatoxin B1 plays a major role in the etiology of human hepatic and extrahepatic carcinogenesis. 21 –23 An increased incidence of hepatocellular carcinoma has been associated with dietary exposure to AFB1, particularly in populations that are commonly exposed to hepatitis B virus. 24,25 Aflatoxin B1 requires metabolic activation in order to elicit its carcinogenic properties. The CYP450 isoenzymes involved in AFB1 bioactivation in human liver are mainly CYP3A4 and CYP1A2. Cytochrome P3A4 (CYP3A4) metabolizes AFB1 to AFB1-exo-8,9-epoxide, while the major product of its enzymatic action is aflatoxin Q1 (AFQ1), a less toxic metabolite. 26 In contrast, CYP1A2 metabolism produces aflatoxin M1 (AFM1) and the AFB1 epoxide. However, the epoxide produced by CYP1A2 is a mixture of exoisomer and endoisomer. 26 It has been reported that only the exoisomer of the epoxide is genotoxic. 27 Detoxification of the AFB1-8,9-epoxide occurs through the glutathione S-transferase (GST)-mediated conjugation with glutathione (GSH). 21
In contrast to these inductive effects on CYP450, no data are available concerning the effect of AFB1 on CYP expression and nuclear receptor activation in humans. In this regard, the aim of the present study was to investigate the effect of AFB1 on nuclear receptors (PXR, CAR, and AhR) and some of their target human CYP450 messenger RNA (mRNA) expression. For this end, we monitored our study on primary cultured human hepatocytes which are the most recommended tools to study CYP inductibility and nuclear receptor activation in humans. 28,29
Materials and Methods
Primary Culture of Human Hepatocytes
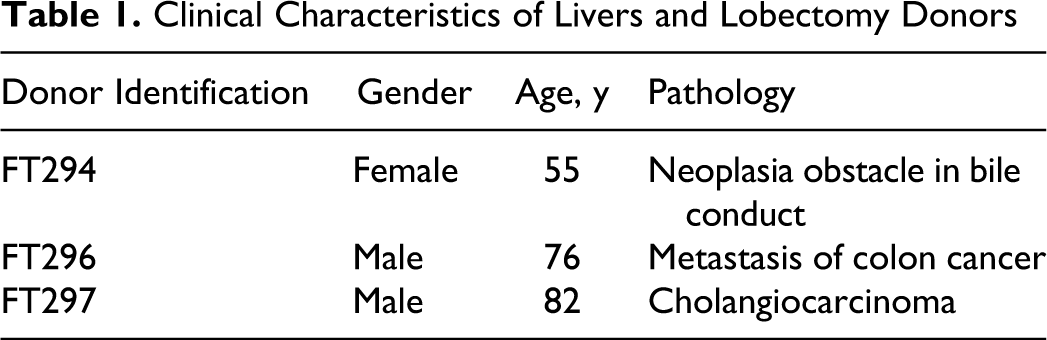
Hepatocytes were prepared from lobectomy segments, resected from adult patients for medical reasons unrelated with our research program. Human hepatocytes from 3 donors, FT294, FT296, and FT297 (Table 1), were isolated as described previously. 30 The entire primary human hepatocytes isolation procedure is approved by the French National Ethics Committee.
Clinical Characteristics of Livers and Lobectomy Donors
Following isolation, cell viability was assessed by the trypan exclusion blue test. Then, cells (90%-95% of viability) were plated on collagen-coated culture dishes at a density of 1.4 × 105 cells/cm2. Culture medium was enriched as described previously 31 for plating with 2% fetal calf serum (v/v). The medium was exchanged for a serum-free medium the day after, and the culture was allowed to stabilize for an additional 48 to 72 hours prior to the treatments. Cultures were maintained at 37°C and 5% CO2 in a humidified incubator.
Cytotoxicity Assay
To evaluate the cytotoxic effect of AFB1 in hepatocytes, we used the adenosine triphosphate (ATP) assay. This assay is based on the detection of a chemiluminescent signal proportional to cellular ATP levels caused by a luciferin–luciferase reaction. The amount of ATP is directly proportional to the number of viable cells in culture. The assay was performed as specified by the manufacturer (CellTiter-Glo Luminescent Cell Viability Assay; Promega, Madison, Wisconsin, USA).
Hepatocytes were seeded in 96-well plates at 5 × 104 cells per well. After an incubation period of 24 hours with increasing concentration of AFB1 ranging from 0.001 to 50 µmol/L or with dimethyl sulfoxide (DMSO) at 0.1% as vehicle, hepatocytes were lysed and total ATP content per well was determined using a luminometer (Packard Instrument Company, Meriden, Connecticut).
Total RNA Purification and Real-Time RT-PCR
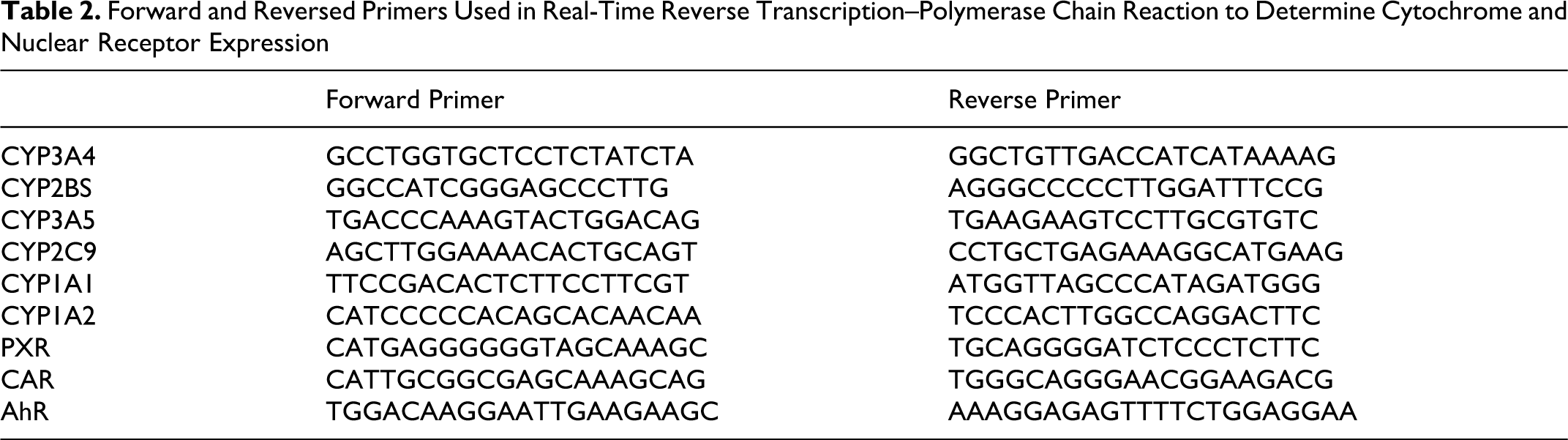
Hepatocytes from FT294, FT296, and FT297 were used for real-time reverse transcription–polymerase chain reaction (RT-PCR) analysis. After an incubation period of 24 hours with increasing concentration of AFB1 ranging from 0.001 to 50 µmol/L, vehicle (DMSO, 0.1%) or positive controls rifampicin (Rif) at 10 µmol/L, phenobarbital (PB) at 500 µmol/L, and 3-methylcholanthrene (3-MC) at 0.1 µmol/L total RNA was isolated from 5 × 105 human hepatocytes using TRIZOL reagent (Invitrogen, Leek, The Netherlands). Complementary DNA was synthesized from 100 ng of total RNA using M-MLV System for RT-PCR at 37°C for 60 minutes in the presence of random hexamers (Amersham Biosciences, Uppsala, Sweden). Real-time PCR was carried out in 96-well plates and monitored by Stratagene MxPro (Agilent Technologies, Santa Clara, USA) under the following general conditions: denaturation program of 95°C for 10 minutes, amplification with 3 segments, and the quantification program repeated for 40 cycles (denaturation at 95°C for 30 seconds, annealing at 65°C for 30 seconds, and elongation and single fluorescence acquisition at 72°C for 40 seconds per 100 base pairs [bp]) followed by 1 cycle (1 minute at 95°C, 30 seconds at 55°C, and 30 seconds at 95°C). No reverse transcriptase sample and water controls were included in each run. The 18s mRNA served as the housekeeping transcript for target mRNA normalization. Messenger RNA quantification was achieved using software provided with the instrument. The 5′ and 3′ primers are indicated in Table 2.
Forward and Reversed Primers Used in Real-Time Reverse Transcription–Polymerase Chain Reaction to Determine Cytochrome and Nuclear Receptor Expression
Statistics
The results were expressed as means ± standard deviations. Statistical differences between control and treated livers were done using 1-way analysis of variance (ANOVA) followed by Dunnett t test. Value of P < .05 was considered significant.
Results
Cytotoxicity Assay
The effect of AFB1 on the acute liver cytotoxicity was assessed by the ATP assay. Treatment of 3 different human livers (FT294, FT296, and FT297) with increasing AFB1 concentrations led to the mortality of hepatocytes as illustrated in Figure 1. It is of note that considerable variation in the response to AFB1 cytotoxicity was found among the different hepatocyte preparations. Hepatocytes from FT296 were less sensitive to AFB1 than hepatocytes from FT294 and FT297. The estimated IC50 concentration leading to 50% of cytolethality for FT294 was approximately 0.5 µmol/L and for FT297 was approximately 50 µmol/L, while in FT296, at the highest tested concentration of AFB1 (50 µmol/L), the percentage of viability was about 67.4% ± 2.3%.
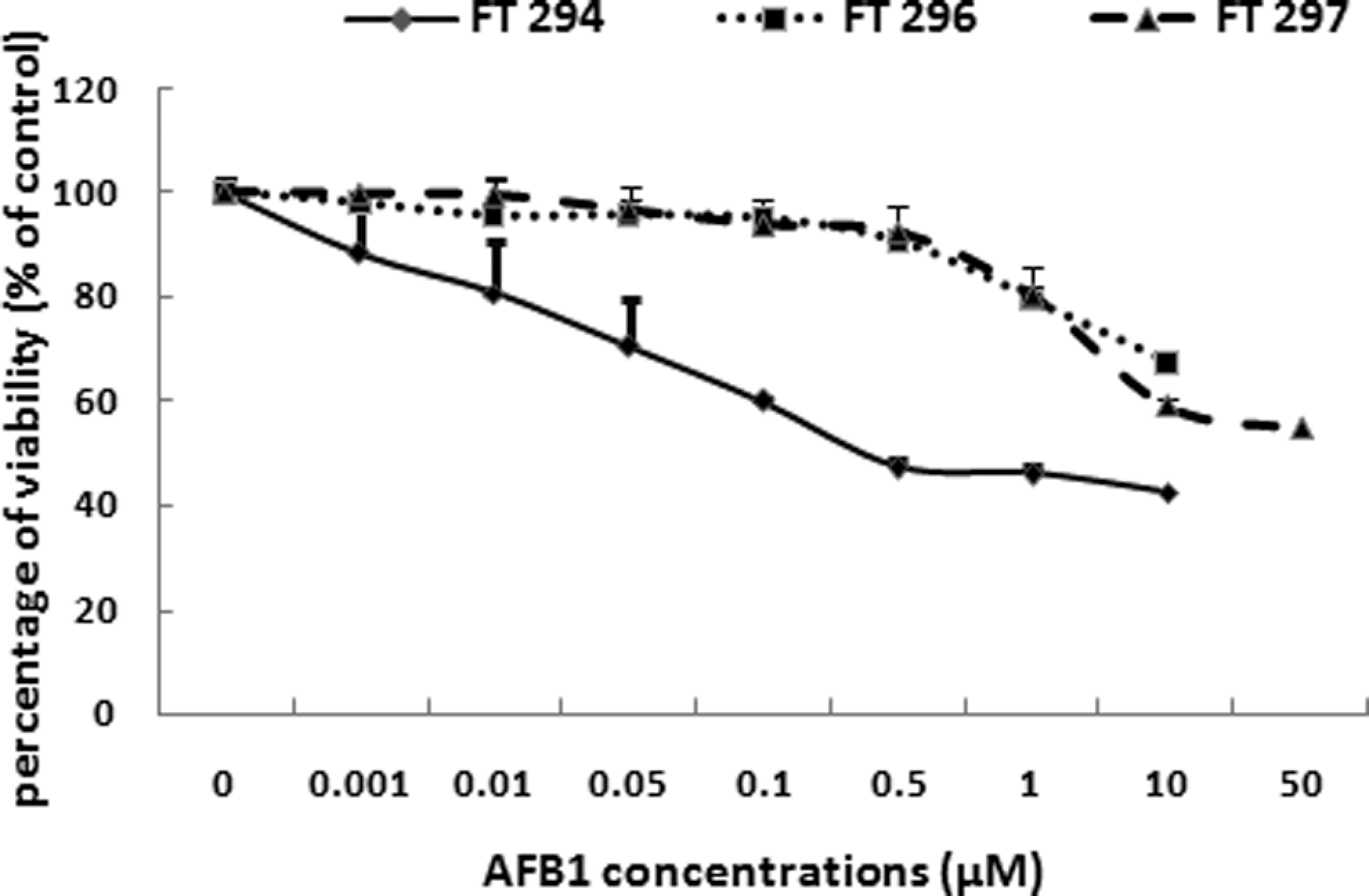
Effect of AFB1 on primary cultured human hepatocytes viability. Cells were treated with AFB1 at the indicated concentrations for 24 hours. Cell viability was determined using the ATP assay. ATP content is directly proportional to the number of metabolically active cells. Data are expressed as the mean ± standard deviation. Analyses were performed in 3 different primary cultures of human hepatocytes (FT294, FT 296, and FT297). AFB1, aflatoxin B1; ATP, adenosine triphosphate.
Messenger RNA Induction of PXR and/or CAR Regulated Cytochromes by AFB1
In the present study, we focused on the effect of AFB1 in mRNA expression of PXR and/or CAR regulated cytochromes. For this purpose, we analyzed the levels of some CYP mRNAs particularly, CYP3A4, CYP2B6, CYP3A5, and CYP2C9. Our results showed that CYP3A4 mRNA was significantly increased by AFB1 treatment already at 0.001 µmol/L (3.41- ± 1.03-fold induction in FT294, 1.87- ± 0.35-fold induction in FT296, and 3.21- ± 1.09-fold induction in FT297; Figure 2A). Cytochrome P2B6 (CYP2B6) mRNA expression was also upregulated with increasing concentrations of AFB1. It was about 14.0- ± 2.7-fold induction in FT294, 3.87- ± 0.6-fold induction in FT296, and 3.97- ± 1.36-fold induction in FT297 at AFB1 1 µmol/L (Figure 2B).
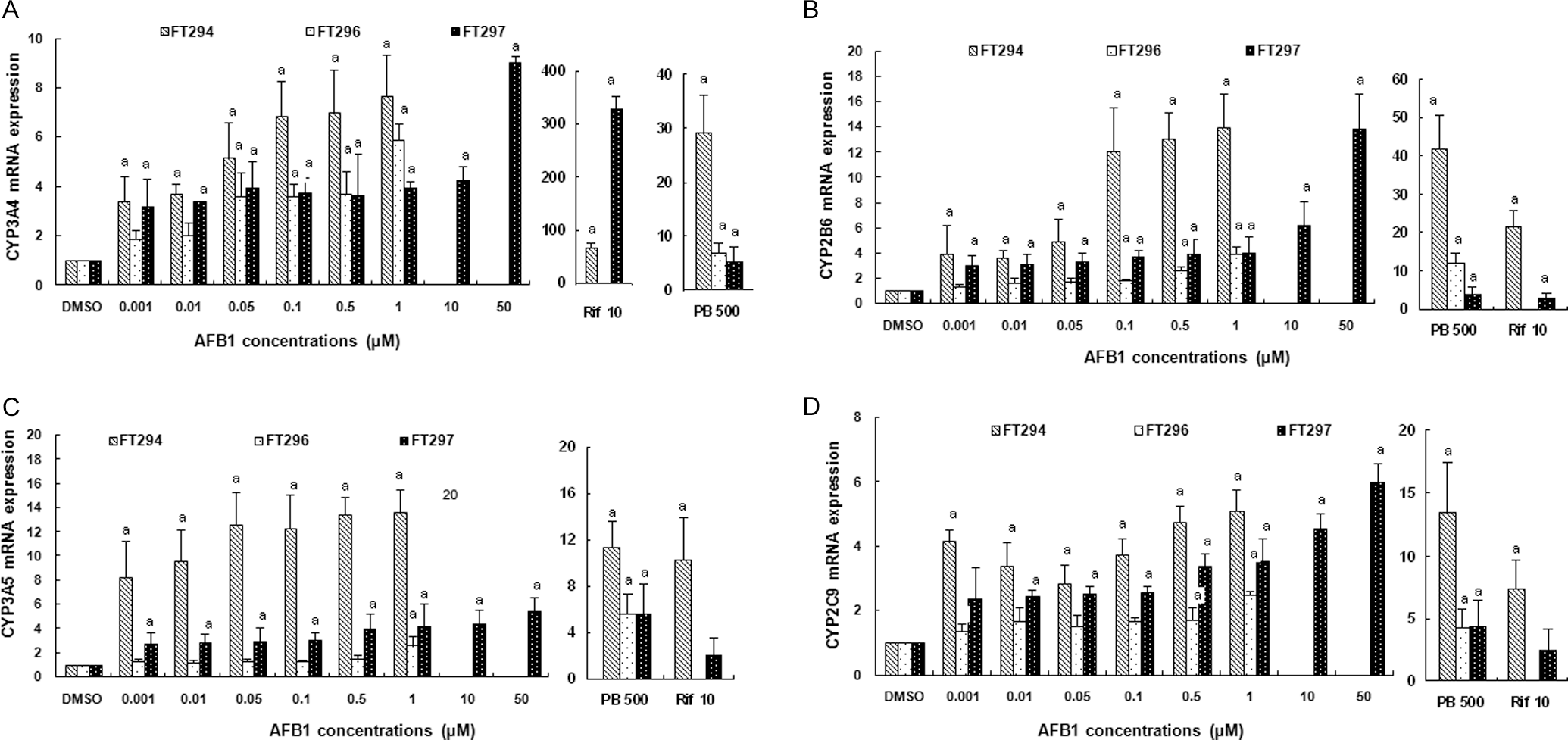
Effect of AFB1 on (A) CYP3A4, (B) CYP2B6, (C) CYP3A5, and (D) CYP2C9 mRNA expression in primary cultured human hepatocytes. Hepatocytes were treated for 24 hours with increasing concentrations of AFB1 ranging from 0.001 to 50 µmol/L. In parallel, cells were treated with DMSO for negative control and with rifampicin at 10 µmol/L and with phenobarbital at 500 µmol/L for positive controls. Values are normalized to 18s mRNA levels. Data are means from duplicates and are expressed as mean ± standard deviation. Analyses were performed in 3 different primary cultures of human hepatocytes (FT294, FT 296, and FT297). “a” indicates the value significantly different from the control value (0 µmol/L) at P < .05. Nonrepresented histogram is a not determined value. AFB1, aflatoxin B1; DMSO, dimethyl sulfoxide; mRNA, messenger RNA.
We also assessed CYP3A5 mRNA expression, it was also induced by AFB1. The maximum of its induction was about 13.67- ± 1.87-fold in FT294 at AFB1 1 µmol/L (Figure 2C). Cytochrome P2C9 (CYP2C9) was also induced by AFB1 treatment already at 0.01 µmol/L of AFB1 (3.41- ± 0.74-fold induction in FT294, 1.68- ± 0.43-fold induction in FT296, and 2.45- ± 0.21-fold induction in FT297; Figure 2D).
We treated hepatocytes with Rif at 10 µmol/L and PB at 500 µmol/L as positive controls. In all treated livers, we found a strong CYP3A4 and CYP2B6 induction by, respectively, Rif and PB. These findings confirm the results described in literature. However, Rif was found to induce an increase in CYP2B6, CYP3A5, and CYP2C9 mRNA levels but lesser than PB did (Figure 2 ).
Messenger RNA Induction of AhR-Regulated Cytochromes by AFB1
We also investigated the effect of AFB1 on AhR-dependent expression of CYP1A1 and CYP1A2 mRNAs. The potency of AFB1 to induce CYP1A1 and CYP1A2 varied between individual human hepatocyte cultures. Aflatoxin B1 caused statistically significant (P < .05) induction of CYP1A1 and CYP1A2 mRNAs in FT294 and FT297 (Figure 3A, B). However, this induction was less observed on FT296 in all tested AFB1 concentrations. Maximal inductions were about 10 ± 1.04 in FT294 at 1 µmol/L AFB1 for CYP1A1 and about 38 ± 5.7 in FT294 at 1 µmol/L AFB1 for CYP1A2 mRNA level.
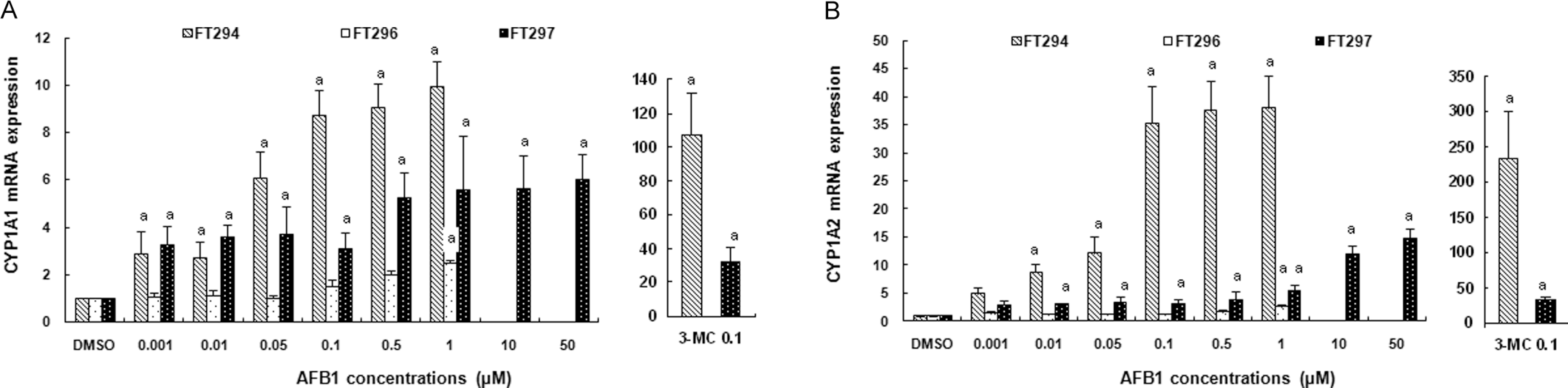
Effect of AFB1 on (A) CYP1A1 and (B) CYP1A2 mRNA expressions in primary cultured human hepatocytes. Hepatocytes were treated for 24 hours with increasing concentrations of AFB1 ranging from 0.001 to 50 µmol/L. In parallel, cells were treated with DMSO and 3-methylcholanthrene at 0.1 µmol/L for negative or positive controls, respectively. Values are normalized to 18s mRNA levels. Data are means from duplicates and are expressed as mean ± standard deviation. Analyses were performed in 3 different primary cultures of human hepatocytes (FT294, FT 296, and FT297). “a” indicates the value significantly different from the control value (0 µmol/L) at P < .05. Nonrepresented histogram is a not determined value. AFB1, aflatoxin B1; DMSO, dimethyl sulfoxide; mRNA, messenger RNA.
3-Methylcholanthrene is known to be the prototypical inducer of AhR and its corresponding target genes. 3-Methylcholanthrene of 10 µmol/L was used as positive control. In agreement with previous reports, 3-MC induced a significant upregulation of CYP1A1 and CYP1A2 expression in 2 primary cultures of hepatocytes. The extent of induction appeared highly variable between the individual livers. Indeed, for CYP1A1, induction by 3-MC was about 107.4 ± 24.5 in FT294; while in FT297, it was about 32.3 ± 8.2. For CYP1A2, induction was about 234.8 ± 66.24 in FT294 and 66.2 ± 4.0 in FT297 (Figure 3 ).
Receptor mRNA Regulation by AFB1
The effect of AFB1 on receptor expression was examined in the 3 primary cultured human hepatocytes. Using real-time RT-PCR, the analysis of receptor mRNA levels clearly showed remarkable interindividual differences among different hepatocyte preparations (FT294, FT296, and FT297). For PXR gene, we observed, at 1 µmol/L of AFB1, 11- ± 1.8-fold, 2.78- ± 0.3-fold, and 4.63- ± 1.04-fold inductions, respectively, in FT294, FT296, and FT297 (Figure 4A). Constitutive androstane receptor was also induced by AFB1 but lesser than PXR. The maximum of its induction was 5.01- ± 0.9-fold induction in FT297 at 50 µmol/L of AFB1 (Figure 4B). The AhR gene was significantly (P < .05) induced in all tested livers (maximal inductions were 4.7- ± 1.6-fold in FT294, 2.67- ± 0.5-fold in FT296, and 3.22- ± 0.74-fold in FT297 at 1 µmol/L of AFB1; Figure 4C).
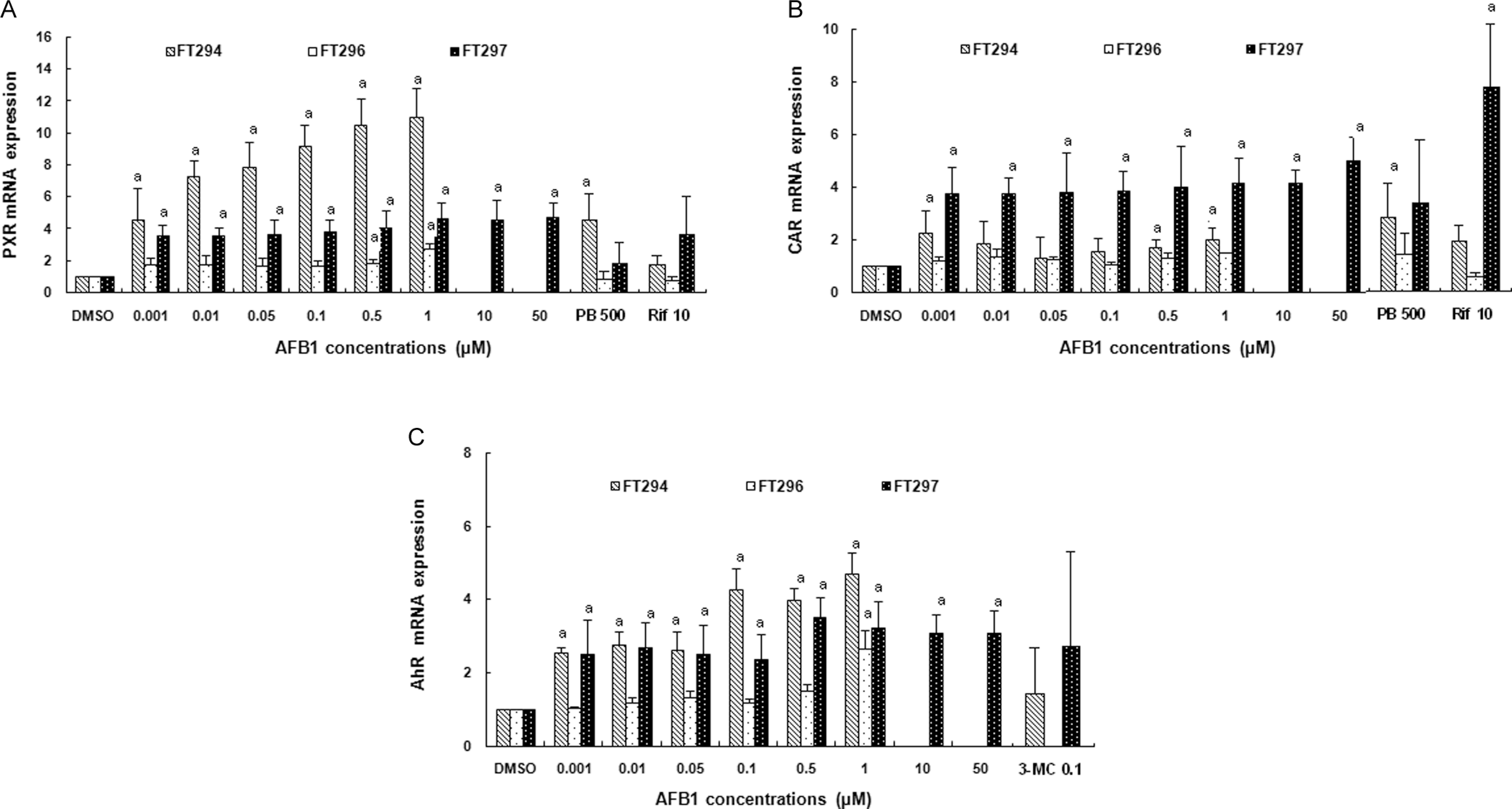
Effect of AFB1 on (A) PXR, (B) CAR, and (C) AhR mRNA expressions in primary cultured human hepatocytes. Hepatocytes were treated 24 hours with increasing concentrations of AFB1 ranging from 0.001 to 50 µmol/L. In parallel, cells were treated with DMSO as vehicle for negative control and/or with rifampicin at 10 µmol/L, phenobarbital at 500 µmol/L, and 3-methylcholanthrene at 0.1 µmol/L for positive controls known to activate PXR, CAR, and AhR receptors, respectively. Values are normalized to 18s mRNA levels. Data are means from duplicates and are expressed as mean ± SD. Analyses were performed in 3 different primary cultures of human hepatocytes (FT294, FT 296, and FT297). “a” indicates the value significantly different from the control value (0 µmol/L) at P < .05. Nonrepresented histogram is a not determined value. AFB1, aflatoxin B1; DMSO, dimethyl sulfoxide; SD, standard deviation; PXR, pregnane X receptor; CAR, constitutive androstane receptor; AhR, aryl hydrocarbon receptor; mRNA, messenger RNA.
Discussion
Aflatoxin B1 is a “procarcinogen” in that enzymatic bioactivation is a prerequisite for carcinogenic and toxic activity. 32 Accordingly, elucidation of the mechanisms of AFB1 metabolism has been the focus of intense research over the years. Aflatoxin B1 is metabolized by hepatic microsomal CYP450s to the reactive, electrophilic exo-AFB1-8,9-epoxide (AFBO) which binds to DNA and other critical cellular macromolecules. 33 Although both CYP1A and CYP3A isoforms oxidize AFB1, there are conflicting reports on their relative importance. 34,35 Aflatoxin B1 metabolism studies in human liver microsomal preparations indicate a predominant role for CYP3A4 and that its expression level was an important determinant of the AFB1 disposition in human liver. 36
Although the effect of AFB1 on the CYP450 activation, especially CYP3A4 and CYP1A1, is well studied, its effect on the expression of CYP and nuclear receptor genes has not been emphasized. Thus, the aim of the present study was to investigate the effect of AFB1 on the mRNA expression of some human CYP450 and nuclear receptors involved in their transcriptional regulation.
First, we evaluated the effect of increasing AFB1 concentrations on the viability of primary cultured hepatocytes. Our results showed that AFB1 reduced cell viability with an estimated IC50 of 5 µmol/L in FT294 and approximately 50 µmol/L in FT297. It is of note that considerable interindividual variations were noticed among the different treated cultures. This is likely to be due to the differences in interindividual variations in both levels and inducibility of hepatic CYP enzymes. Several investigations reported that these interindividual differences can be due to environmental factors (eg, diet, cigarette smoking, and exposure to enzyme-inducing pollutants), physiological factors (eg, age, disease, and endocrine homeostasis), and polymorphisms in CYP genes, nuclear receptors, regulatory proteins, and transporters. 37 –39
Second, we examined the effect of increasing concentrations of AFB1 on some CYP450 and nuclear receptor (PXR, CAR, and AhR) expressions in primary cultures of human hepatocytes. Our results clearly showed that AFB1 induced CYP2B6 and CYP3A5 mRNA overexpression in a concentration-dependent manner. Cytochrome P3A4 (CYP3A4) and CYP2C9 were also induced by AFB1 but in a lesser extent than CYP3A5 and CYP2B6 (Figure 2). Induction of CYP3A, CYP2B, and CYP2C predominantly takes place via the PXR and/or CAR nuclear receptor. 40,41 Thus, we tested whether AFB1 is capable of inducing PXR and CAR mRNA. In the 3 treated livers, AFB1 increased PXR and CAR gene levels (Figure 4 ). Induction of CYP3A, CYP2B, and CYP2C overexpression accompanied with enhancement in PXR and CAR mRNA levels suggested that AFB1 could activate these nuclear receptors. Aflatoxin B1 could activate PXR, CAR, or both at the same time. In fact, PXR overlaps functionally with CAR in terms of ligand binding and gene activation. Both receptors can be activated by the same xenobiotic and endobiotic compounds. 42 Pregnane X receptor and CAR were originally shown to regulate CYP3A and CYP2B genes, respectively. 43,44 It was then demonstrated that PXR could upregulate CYP2B gene using the same response element used by CAR and, vice versa, for upregulation of CYP3A gene of CAR, 45 suggesting a functional redundancy in the CAR and PXR signaling pathways.
Aflatoxin B1 could activate PXR and/or CAR directly if this mycotoxin acts as a ligand to nuclear receptors. In this case, we can assume that following AFB1 binding, PXR as well as CAR, translocate from the cytosol to the nucleus and form a complex with retinoid X receptor α ([RXRα] NR2B1). Subsequently, the heterodimer binds response elements in the promoters of target genes. 46 –48 Further investigations such as the in vitro reporter gene assay are needed to confirm PXR and/or CAR activation by AFB1 and to recognize the molecular mechanism of AFB1 interaction with these receptors.
In the present work, we have demonstrated that AFB1 was able to upregulate AhR expression in human cultured primary hepatocytes already at AFB1 0.001 µmol/L (Figure 4). Aflatoxin B1 increased both CYP1A1 and CYP1A2 gene expression in the 3 treated livers (Figure 3). These findings suggested that AFB1 could activate AhR. Aflatoxin B1 presumably activated AhR via the classical pathway in which after AFB1 binding to receptor, it translocates to the nucleus and heterodimerizes with ARNT and then this heterodimer binds to the consensus regulatory sequences (xenobiotic response elements [XREs]) located upstream in the promoter of target genes (eg, CYP450 such as CYP1A1).
It is noteworthy that, in contrast to PXR whose activation requires a direct ligand binding, CAR and AhR activations may occur in a ligand-independent pathway. 49 –53 Thus, further experiments are needed to confirm nuclear receptor activation by AFB1 and precisely define the molecular mechanism of interaction between this mycotoxin and receptors.
Activation of nuclear receptors by AFB1 may be a useful tool in the treatment of tumors induced by AFB1. In fact, butylated hydroxytoluene was found to protect against AFB1 toxicity in turkeys through inhibition of CYP1A1 activity. 54,55 Sulforaphane inhibits AFB1-DNA adduct formation via transcriptional repression of CYP3A4 and CYP1A2 in primary cultures of human hepatocytes. 56 Furthermore, specific CYP3A4 inhibitors like troleandomycin have been shown to inhibit AFBO production, 35 while inducers of CYP3A4 activity such as 3-MC and Rif increase AFB1 metabolism in cultured human hepatocytes. 57 Aforementioned, the expression of CYPs, particularly CYP3A and CYP1A, is under the transcriptional control of PXR and/or CAR and AhR. So that, treatment with nuclear receptor inhibitors can lead to inhibition of CYP transcription and activity and subsequently, inhibition of P450-mediated AFB1 bioactivation. Until now, many AhR antagonists that have been evaluated for their ability to directly inhibit the CYP1 family were found to be efficient in chemopreventive strategy. 58 It would be interesting to evaluate the effect of nuclear receptor antagonists on AFB1 toxicity.
Conclusion
According to our knowledge, this is the first report trying to investigate AFB1 effect on nuclear receptors and CYP450 expression in human hepatocytes. Primary human hepatocytes are a popular in vitro system for drug biotransformation research because of their strong resemblance of in vivo human liver. Our results showed that AFB1 induced an upregulation of PXR, CAR, and CYP2B6, CYP3A5, CYP2C9, and CYP3A4 genes. Aflatoxin B1 also increased AhR and CYP1A1 and CYP1A2 mRNA expressions. Further investigations are needed to confirm PXR and/or CAR and AhR activation by AFB1 and to recognize the molecular mechanism of the interaction between AFB1 and nuclear receptors. It would be interesting to evaluate whether nuclear receptor activation by AFB1 could modulate its carcinogenicity.
Footnotes
Acknowledgment
This research was supported by the Ministère Tunisien de l’Enseignement Supérieur et de la Recherche Scientifique et de la Technologie (Laboratoire de Recherche sur les Substances Biologiquement Compatibles: LRSBC).
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) received no financial support for the research, authorship, and/or publication of this article.