
Abstract
The effects of salvinorin A (Salvia divinorum principal ingredient), a potent κ-opioid natural hallucinogen, on learning and memory were investigated. Wistar rats were tested in the 8-arm radial maze, for object recognition and passive avoidance tasks for spatial, episodic, and aversive memory. Attention was assessed using a latent inhibition task. Salvinorin A (80-640 μg/kg subcutaneous [sc]) did not affect short-term memory, but it impaired spatial long-term memory. Episodic and aversive memories were impaired by salvinorin A (160-640 μg/kg). Memory impairment was blocked by the selective κ-opioid receptor antagonist, nor-binaltorphimine ([nor-B]; 0.5-1 mg/kg, intraperitoneal [ip]). Salvinorin A (160 μg/kg) disrupted latent inhibition, after LiCl treatment, such as reduced sucrose intake, suggesting an attention would result in an impairment of cognitive behavior. These findings demonstrate for the first time that salvinorin A has deleterious effects on learning and memory, through a κ-opioid receptor mechanism.
Introduction
Salvia divinorum, a member of the Lamiaceae family contains salvinorin A, the main active constituent. 1 –4 Salvinorin A is a κ-opioid receptor agonist noted for its low toxicity 5 and is one of the most potent natural hallucinogens known until now. 6 Salvia divinorum, native to certain areas in the Sierra Mazateca of Oaxaca, Mexico, prefers a hot humid semitropical climate between 750 and 1500 m altitude. It is a perennial herb widespread in Mexico and South America. Furthermore, in many other countries, including the United States, it is legal to purchase S divinorum from Internet suppliers. Awareness of the hallucinogenic effects of S divinorum has grown recently, due to an increase in public representation and concern over its potential harmful effects. 7 The effects of salvinorin A on behavior have been studied under several conditions in rodents and rhesus monkeys. 8 Salvinorin A caused sedation and loss of motor coordination in an inverted screen task in mice 9 at doses between 0.5 and 2 mg/kg intraperitoneally (ip) and in rhesus monkeys treated intravenously (iv). 10 Salvinorin A showed discriminative stimulus effects in rats and rhesus monkeys 2,11 –13 and generalized to the discriminative stimulus effects of structurally diverse, centrally penetrating κ-agonists (bremazocine, U69593, and U50488) using an operant conditioning task where the number of correct choices on the drug appropriate lever was evaluated. 3,14 Salvinorin A increased immobility and reduced rat swimming behavior in a forced swim test 1,3,15 at doses between 0.25 and 2 mg/kg. Such effect was accompanied by a decrease in phasic dopamine release without affecting dopamine reuptake in the nucleus accumbens core and shell. Antinociceptive effects, evaluated in terms of increased threshold latency to radiant heat applied to the tail, using the tail flick test, were also found in mice. 16,17 Additionally, conditioned place aversion was followed by a drop in the dopamine level in the caudate-putamen—the putamen is a structure in the forebrain and with the caudate nucleus forms the dorsal striatum— in mice through the activation of a κ-opioid subtype receptor. 18
Recently, salvinorin A was shown to have rewarding (that is motivational) effects in the conditioned place preference in zebrafish 19 and in the intracerebroventricular self-administration tasks in rats. 20 This effect was accompanied by increased extracellular dopamine content in the shell of the nucleus accumbens (part of the limbic system) which is involved in pleasure as observed for other drugs of abuse, 21 suggesting the potential for addiction in humans. Pretreatment with nor-binaltorphimine (nor-B) and the cannabinoid receptor subtype 1 (CB1) antagonist, rimonabant, reduced the motivational effect, indicating an involvement of both κ-opioid and CB1 cannabinoid subtype receptors. Possible anxiolytic- and antidepressant-like effects in rats, evaluated through the elevated plus maze and forced swim test, respectively, have also been reported. 22 However, depending on the dose, salvinorin A can produce pro-depressant effects in terms of increased immobility time in the forced swim test in rats. 15,23 Binding and functional studies indicate that salvinorin A was more active than the prototypical κ-opioid receptor agonists U50488 and U-69593. 3,24 Recent findings indicate that κ-opioid receptors are rapidly emerging as an important target for the study and treatment of some central nervous system disorders (eg, Alzheimer disease, schizophrenia, bipolar disorder, cocaine abuse) and in the treatment of pain. 25 –28 Salvinorin A has become a popular recreational drug widespread throughout Europe and North America similar to other natural drugs. Salvinorin A is marketed, primarily to adolescents and young adults, as a “safe” hallucinogen, 29 and is sold through smart shops and Internet Web sites. Although information about this drug is limited, recent Salvia-related media reports and Internet traffic suggest the possibility that its abuse is increasing in the United States and Europe. The 2010 NIDA’s monitoring survey publication 30 reported that out of 46 482 students in 8th, 10th, and 12th grades from 396 public and private schools, 5.5% have used S divinorum. This is greater than the percentage of students reported to have used ecstasy (4.5%), heroin (0.9%), cocaine (2.9%), methamphetamines (1.0%), oxycodone (5.1%), LSD (2.6%), GHB (1.4%), and ketamine (1.6%). However, it has been shown 29,31,32 to induce various symptoms of psychiatric disorders, including dissociation, perceptual distortion, depersonalization, and feelings of spatiotemporal dislocation after inhalation of the vaporized active constituent in a range of exposure between 200 and 500 μg. One case has been reported of a Salvia user who committed suicide. 33 κ-opioid receptors are enriched in different brain areas. 28
In line with their localization in the hippocampus, amygdala, hypothalamus, striatum, and spinal cord, κ-opioid receptors are related to learning and memory, emotional control, stress response, and pain. 34 However, the contribution of the κ-opioid system to learning and memory performance has produced contradictory results. 35 Given the well-known activation of κ-opioid receptors by salvinorin A, we hypothesized that rats exposed to this hallucinogen might show cognitive alterations. Accordingly, a significant change in cognition, evaluated through a Hallucinogen Rating Scale–Cognition test, was found in volunteers taking, by inhalation, salvinorin A (15-21 μg/kg). 36 Salvinorin A has also been shown to produce a pattern of disruptive effects in the 5-choice serial reaction time task, a food-motivated test that quantifies attention, similar to ketamine in rats. 37 The learning tasks employed an 8-arm radial maze, passive avoidance, and object recognition measures of spatial, aversive, and episodic memory, respectively. To investigate a possible attention deficit, the latent inhibition task was also used. A second goal was to evaluate the role of κ-opioid receptor for the effects of salvinorin A using the κ-opioid receptor antagonist, nor-B.
Materials and Methods
Animals
Male Wistar rats (Charles River, Calco, Italy) weighing 250 to 300 g on arrival were housed 4 per cage (68 × 38 × 20 cm) in an air-conditioned room with a 12-hour light/12-hour dark cycle (lights on 08:00-20:00 hours) with free access to food and water. The animals were allowed to acclimatize for at least 10 days before the experiments. Then they were randomly divided into groups (10 naive rats per group). All testing took place during the first half of the light period (between 09:00 and 13:00 hours). Animals were used only once and only for 1 test each. All the experimental procedures followed the guidelines of the Italian Council on Animal Care approved by Italian Government Decree No. 28/2010. All efforts were made to minimize the number of animals used and their suffering.
Spontaneous Motor Activity
Spontaneous motor activity was evaluated as previously described by Braida et al 38 in an activity cage (43 × 43 × 32 cm; Ugo Basile, Varese, Italy) placed in a sound-attenuating room. The cage was fitted with 2 parallel horizontal infrared beams 2 cm from the floor. Cumulative horizontal movements were recorded for 15 and 20 minutes after treatment with salvinorin A.
Radial Maze
The effects of salvinorin A on spatial memory were studied in an 8-arm radial maze, as previously described. 39 Briefly, the wooden maze, raised 50 cm above the floor, had an octagonal central platform with 8 arms separated by Plexiglas guillotine-type doors that the experimenter could raise and lower. A food pellet was placed at the end of each arm. Rats were kept at 85% of initial body weight. Performance in the maze was tested by placing each animal on the platform with all the doors closed. After a 1-minute acclimatization period, all the doors were simultaneously opened and each animal was trained to complete the maze until it either successfully visited all 8 arms or 10 minutes had elapsed. After each trial, the maze was cleaned with 2.5% cider vinegar solution and dried thoroughly to eliminate scent trails.
Procedure
During each session, short-term memory was scored on the basis of the number of total errors, the number of correct choices before the first error, and the total time taken to complete the test. An error was defined as re-entry in an arm already visited. Training, at the rate of 1 session/day, continued until the rats had fulfilled the criterion of entering 7 different arms out of the first 8 choices on 5 successive days. They were then given a daily dose of saline (ip injection) or vehicle (subcutaneous [sc] injection) to accustom them to handling and treatment before the actual drug and maze testing. When the baseline response was stable (a week at the most), groups of 10 rats each were randomly assigned to the different treatments. The performance obtained when each rat was given its appropriate vehicle the day before treatment served as control. Simultaneously, the pattern of arm entries, which permits analysis of the angle chosen when a rat enters 2 consecutive arms, was examined. 40 In the 8-arm radial maze, there are 5 possible angles: from 0°, which corresponds to re-entry into the arm just visited, to 180°, which corresponds to entry into the arm directly opposite the one just visited. Angles of 45°, 90°, and 135° are also possible. The first 8-arm entries of each session were used for the analysis. The frequency of each choice was calculated as (number of observations/7) × 100. The response-strategy criterion was defined as the choice of the same angle on more than 44% of all turning responses during a 5-day block of trials. 41 The effect of drug treatment on the pattern strategy was evaluated as the percentage of animals that changed the angle frequency during drug treatment compared to the response-strategy criterion. This was indicative of the flexibility of behavior in situations that required adjustment of the working strategy. 42
Delay procedure
To study long-term memory, we assessed the effect of introducing a 2-hour delay between the fourth and fifth choices on 3 consecutive days in additional groups. This delay was chosen on the basis of a previous study 38 which investigated the worsening of performance in rats during a 5- to 240-minute period. Rats were allowed to randomly enter 4 arms to get the reward (pre-delay). Access to the other 4 arms was prevented by removing each rat from the maze. Then each rat was placed back in its single cage and transported to an adjacent dim observation room, where it was left for 2 hours, after which each rat was transported back to the maze room, put on the center platform, and allowed to complete the remaining 4 arms (post-delay). Treatments were given sc or ip 20 minutes before the pre-delay test. During each post-delay session, performance was scored in terms of the number of total errors, the number of correct choices before the first error, the total time taken to complete the test, and the pattern of arm entry.
Object Recognition
The object recognition task was conducted in an open plastic box. 43 The objects to be discriminated were yellow plastic cylinders, round brown boxes, and green pyramids. There were 3 copies of each object, which were used interchangeably. To rule out the possibility of scent traces left on the objects, they were wiped between each trial with a piece of rag saturated with natural rat odor that had been stored in dirty sawdust from cages housing other rats of approximately the same age. At the end of each day’s experiments, the objects were cleaned with 70% alcohol solution, dried, and again stored in the dirty sawdust. Before testing, rats were allowed to individually explore the apparatus for 5 minutes. Twenty-four hours later, for the first 3-minute sample trial (T1), 3 identical objects were presented in 2 corners of the box. In the second 3-minute choice trial (T2), one of the objects presented in T1 (the “familiar object”) was replaced by a new object. T1 and T2 were separated by a 1-hour interval. The basic measure was the time (in seconds) the rats spent exploring the objects in the 2 trials. Animals received vehicle or salvinorin A 20 minutes before T1. Performance was evaluated as the time spent exploring objects during T2 and calculating a discrimination index (N − F/N + F), where N is the time spent exploring the new object during T2 and F the time spent exploring the familiar object during T2. 44
Passive Avoidance
The step-through type passive avoidance task was used, as previously described, 45 to examine aversive memory. Briefly, the apparatus consisted of 2 compartments, 1 light and 1 dark, connected by a sliding door. During the acquisition trial, each rat was placed in the light compartment and allowed to enter the dark compartment; the time (in seconds) taken to enter was recorded. Once the rat was in the dark compartment, the sliding door was closed and an unavoidable electric shock (0.8 mA for 5 seconds) was delivered to the paws. The retention trial was repeated 24 hours after the acquisition trial, by positioning the rat in the light compartment and recording the time taken to enter the dark one (retention latency). A cutoff time of 300 seconds was used during the retention trial. If the animal did not enter the dark compartment within 300 seconds, it was removed and assigned a maximum score of 300 seconds. The percentage of amnesic animals was also calculated. A rat was considered amnesic when its latency to re-enter was <75 seconds. Treatment was given before the acquisition trial.
Latent Inhibition
The latent inhibition paradigm was carried out, which consisted of 3 days of preexposure, 1 conditioning day, and 1 test day.
46
Twenty-four hours prior to the start of the preexposure phase, the water bottles were removed from the cages. Over the next 3 days, the rats received a sc injection of vehicle or salvinorin A (160 μg/kg). Immediately after this injection, the animals were given free access to a bottle containing a 5% sucrose solution (the preexposed group) or plain water (the non-preexposed group) for 30 minutes. On day 4, the conditioning day, the animals again received an ip injection of salvinorin A or vehicle, but all animals were given free access to a bottle containing a 5% sucrose solution for 30 minutes. Immediately after this period, all animals were given an ip injection of 75 mg/kg LiCl (10 mg/mL; Sigma-Aldrich, St. Louis, Missouri; to induce visceral malaise). On day 5 of the test day, the animals did not receive any injection but were given free access to a bottle containing a 5% sucrose solution and a bottle containing water for 30 minutes. The bottles were weighed prior to the experiment and immediately after the 30-minute drinking period in order to establish the consumption (transformed to milliliter consumption). The degree of conditioned taste aversion was determined by calculating the percentage of sucrose consumption on day 5 relative to the total fluid intake on that day. All experiments were conducted between 9:00 and 11:00
Drugs and Treatment
Salvinorin A (Tocris Bioscience, Cookson Bristol, UK) was dissolved in a vehicle containing ethanol, Tween 80, and saline in a proportion of 1:1:8. Nor-B (17, 17'-(dicyclo-propylmethyl)-6,6',7,7'-6,6'-imino-7,7'-binorphinan-3,4',14,14'-tetrol dihydro chloride; Tocris Bioscience) was dissolved in saline. The solutions were prepared daily. Salvinorin A was injected sc at a volume of 1 mL/kg 20 minutes before each test. Nor-B was injected ip at a volume of 5 mL/kg 2 hours before radial maze, passive avoidance, and object recognition tests. The pretreatment time was chosen on the basis of our previous work with salvinorin A on anxiety-like behavior.
22
Nor-B was given ip 2 hours before salvinorin A. It has been reported that κ-opioid receptor antagonism of U50488-induced analgesia peaked 2 hours after treatment then leveled off up to 96 hours.
47
In the antagonism experiments, the control group received saline ip 100 minutes before the vehicle of salvinorin A sc. The control group received only the vehicle of salvinorin A sc for the evaluation of locomotor activity and latent inhibition tasks. Animals were divided into groups of 10 each. For locomotor activity tests, different groups received salvinorin A (80-160-320-640 μg
Statistics
All statistical analyses were done using Prism 5 software (GraphPad Software Inc, San Diego, CA, USA). Vehicle groups (sc and ip) were pooled for analysis since a preliminary statistical analysis test found no differences between the groups. Multiple group comparisons were performed by either 1- or 2-way analysis of variance (ANOVA) followed by Tukey or Bonferroni procedure for comparisons of means. Fisher exact probability test was used to assess the significance of the percentage of amnesic animals in the passive avoidance task and the number of animals changing angle frequency in the radial maze. The level of significance was taken as P < .05. Data were expressed as mean ± standard error of the mean (SEM).
Results
Spontaneous Motor Activity
Salvinorin A did not affect the mean number of horizontal movements at any of the doses tested, compared with the saline (2024 ± 127) and vehicle (1957 ± 146) groups (Figure 1 ).
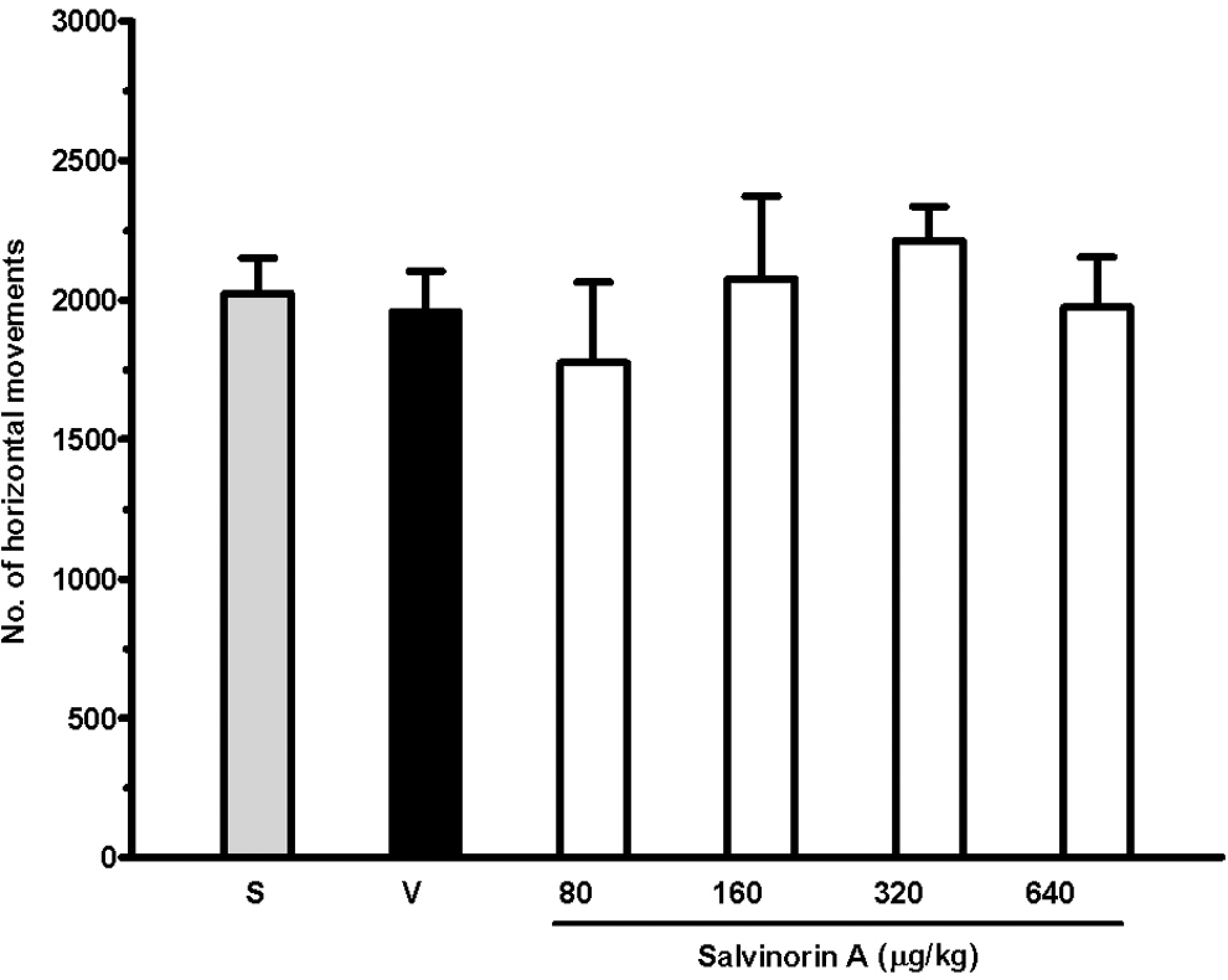
Effects of increasing doses of salvinorin A subcutaneously (sc) on locomotor response in terms of horizontal movements. Rats were treated with saline (S), vehicle (V), or the drug and after 20 minutes were placed in an activity cage for 15 minutes. Their cumulative activity is expressed as the mean ± standard error of the mean ([SEM] 10 rats per group).
Radial Maze
To satisfy the criterion, training lasted 10 to 30 days. Neither vehicle nor salvinorin A (80-640 μg/kg) affected working memory, in terms of mean total number of errors (Figure 2A), mean total time to complete the task (Figure 2B), and mean number of correct choices before the first error (data not shown). Figure 2C shows the percentage of animals changing the pattern of arm entries after treatment. There were not any significant changes in the vehicle group (10%), whereas the number of animals changing the mean angle frequency increased after salvinorin A (160-640 μg/kg; P < .05 or P < .01, Fisher exact probability test). The pretreatment with only nor-B did not alter any radial maze parameter and significantly blocked salvinorin A-induced (640 μg/kg) changes in the percentage of animals changing angle frequency in comparison to the pre-drug value (P < .01, Fisher exact probability test). No significant difference in vehicle and nor-B + salvinorin A groups was shown. Even if it appears that 160 μg/kg induced the highest percentage of animals changing angle frequency, no differences among groups (160-320-640 μg/kg) were statistically significant.
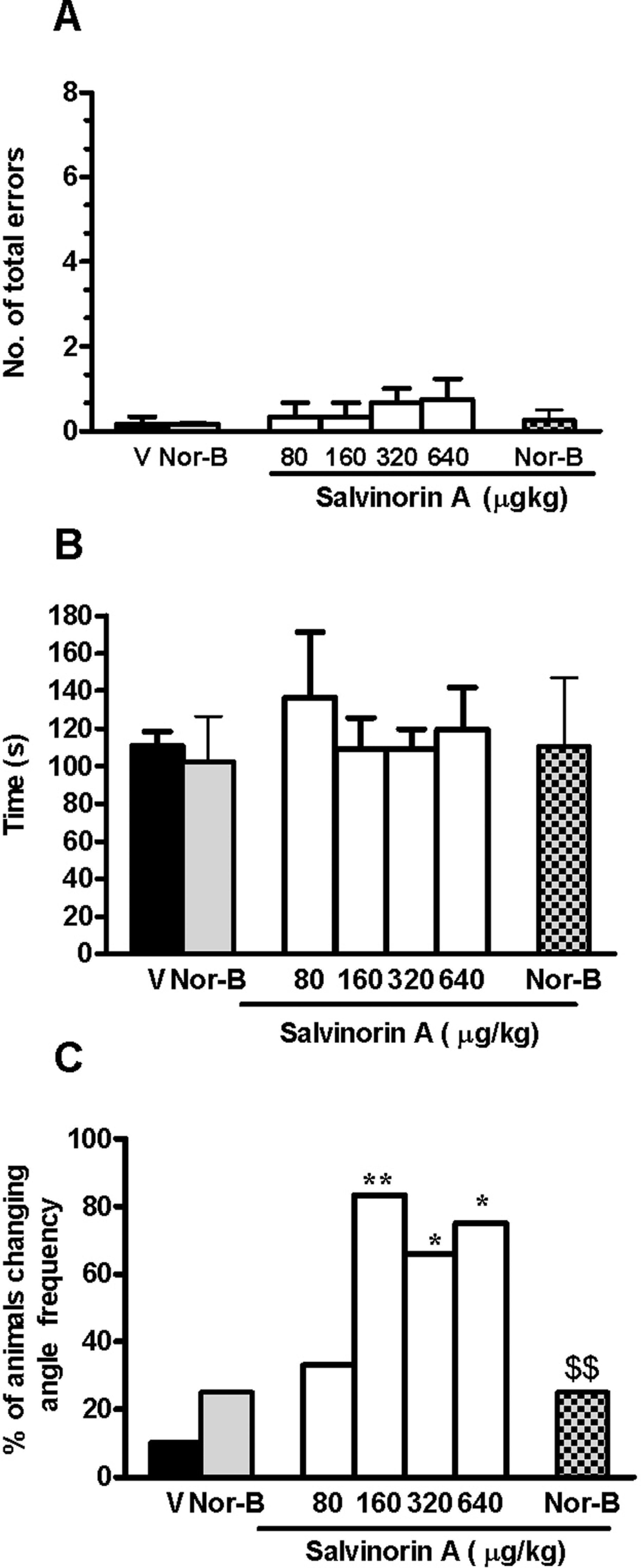
Radial maze performance of salvinorin A alone or in combination with nor-binaltorphimine (nor-B). Salvinorin A was injected subcutaneously (sc), 20 minutes before the test and nor-B (1 mg/kg intraperitoneally [ip]) 100 minutes before salvinorin A (640 μg/kg). Data are expressed in terms of the number of total errors (A), time to complete the maze (B), and the percentage of animals changing the pattern of arm entries in comparison to the predrug value (C). Panels A-B show means ± standard error of the mean ([SEM] 10 rats per group). V indicates vehicle. *P < .05, **P < .01 compared with vehicle and nor-B alone; $$P < .01 compared with salvinorin A 640 μg/kg (Fisher exact probability test).
Radial maze delay
All the rats responded with a similar level of accuracy (≤1 error) in the pre-delay test (data not shown). However, in the post-delay test (choices 5-8), ANOVA revealed a difference between the groups for the total number of errors (P < .0001) and time (P < .0001; Figure 3A-C). In addition, correct choices significantly changed (P < .0001; data not shown). Comparisons indicated that during the post-delay test, all groups treated with salvinorin A performed worse than those given vehicle. Pretreatment with only nor-B did not affect post-delay performance and reversed the increase in the number of total errors and time induced by salvinorin A at the maximal dose (640 μg
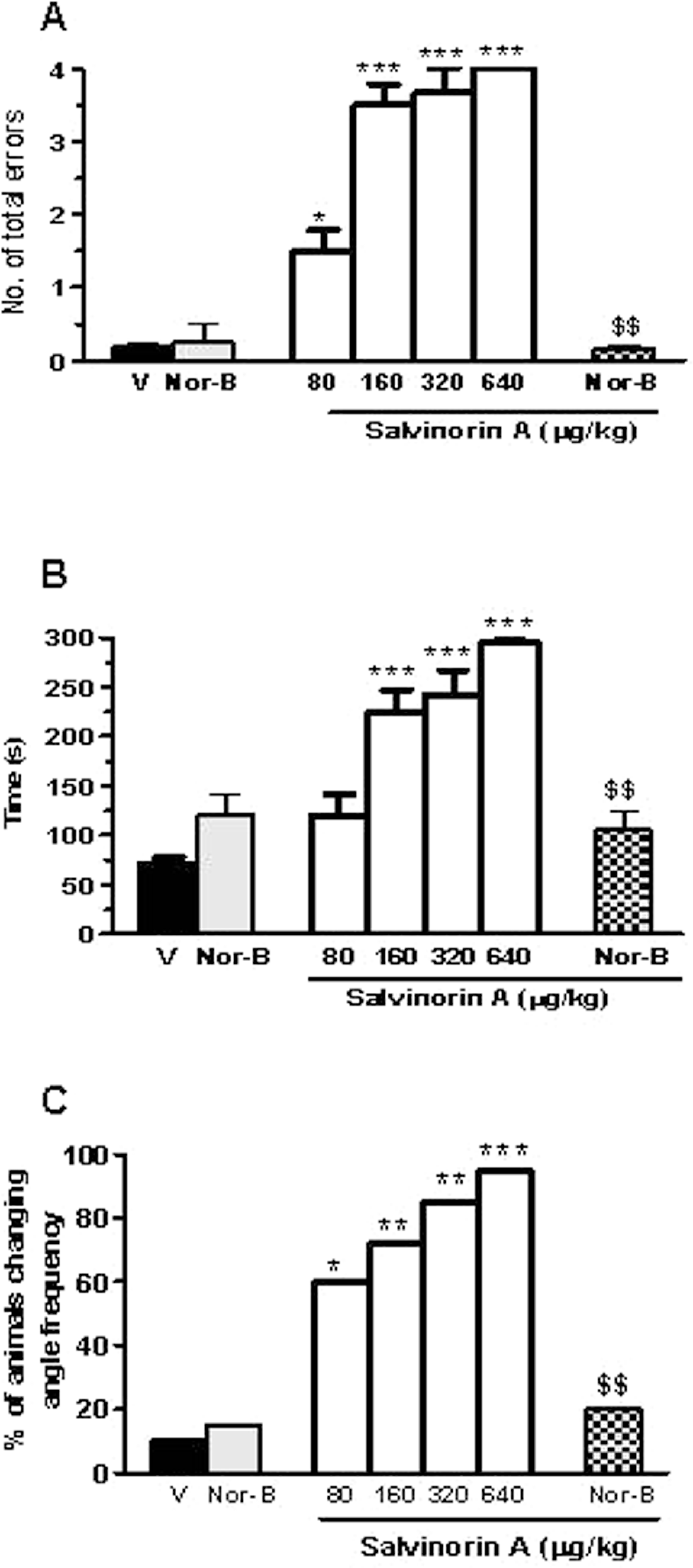
Radial maze performance of salvinorin A alone or in combination with nor-binaltorphimine (nor-B) after a 2-hour delay between the fourth and fifth arm choices in the 8-arm radial maze. Salvinorin A was injected subcutaneously (sc), 20 minutes before the test and nor-B (1 mg/kg intraperitoneally [ip]) 100 minutes before salvinorin A (640 μg/kg). Performance was evaluated in terms of the number of total errors (A), time taken to complete the test (B), and the percentage of animals changing patterns of arm entries in comparison to the predrug value (C). V indicates vehicle. Panels A-B show means ± standard error of the mean ([SEM] 10 rats per group). *P < .05; **P < .01; ***P < .001 compared with vehicle or nor-B alone; $$P < .01 compared with salvinorin A 640 μg/kg; (analysis of variance [ANOVA] followed by Tukey test or Fisher exact probability test).
Object Recognition
During the training session (T1), the groups did not differ significantly in the mean total amount of time spent exploring the 2 objects (data not shown). Nevertheless, there was a significant treatment effect on the mean time spent investigating the novel object during T2 (P < .0001; Figure 4A) and on the discrimination index (P < .01; Figure 4B). Post hoc comparison showed that vehicle- and nor-B-treated rats spent more time exploring a novel object than the object they had been exposed to 1 hour earlier, resulting in a mean discrimination index of 0.79 (± 0.1) and 0.53 (±1.2), respectively. The dose of 640 μg/kg salvinorin A was the maximal dose impairing the time spent exploring the new object. The pretreatment with nor-B in combination with the maximal effective dose of salvinorin A blocked salvinorin A-induced learning and memory impairment. No significant difference was found between vehicle and nor-B + salvinorin A groups.
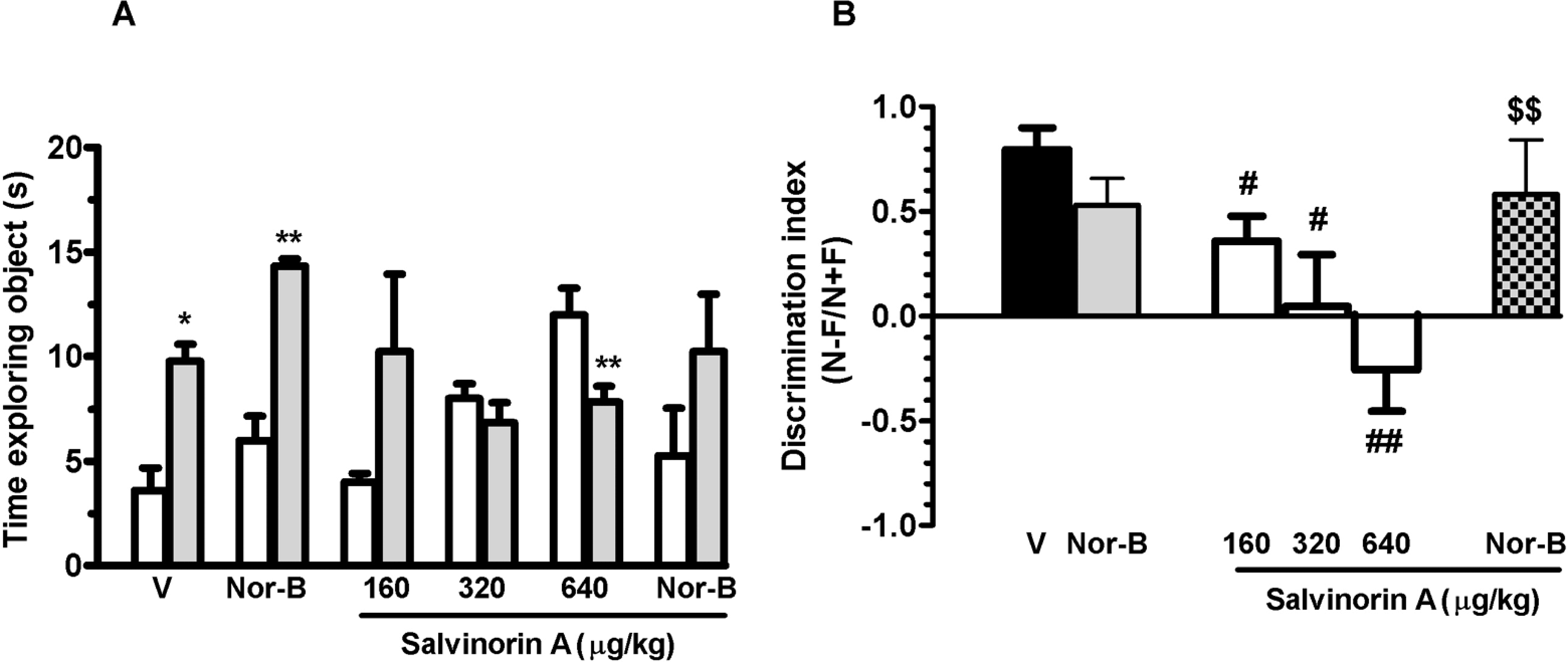
Recognition of novel object evaluated after salvinorin A injection given subcutaneously (sc), 20 minutes before the sample trial (T1), alone (panels A, B) or with nor-binaltorphimine (nor-B). Data are shown as time spent exploring a familiar (white column) and a novel object (gray column) during T2 (1 hour later) (A) and discrimination index (B), where N is the time spent exploring the new object and F is the time spent exploring the familiar object during T2. Each column represents the mean ± standard error of the mean (SEM) of 10 rats. V indicates vehicle; nor-B (1 mg/kg ip) was given 100 minutes before salvinorin A (640 μg/kg). *P < .05, **P < .01 compared to the corresponding familiar object group; #P < .05, ##P < .01 compared with vehicle alone group; $$P < .01 compared with salvinorin A 640 μg/kg; (analysis of variance [ANOVA] followed by Tukey test).
Passive Avoidance
For the antagonism studies, in view of the lack of published data, we ran a pilot study with nor-B to find a dose that did not affect learning and memory (Figure 5
). There was a treatment effect for nor-B (P < .005). Comparison indicated that this drug, given 2 hours before training at doses of 0.1 and 0.5 mg
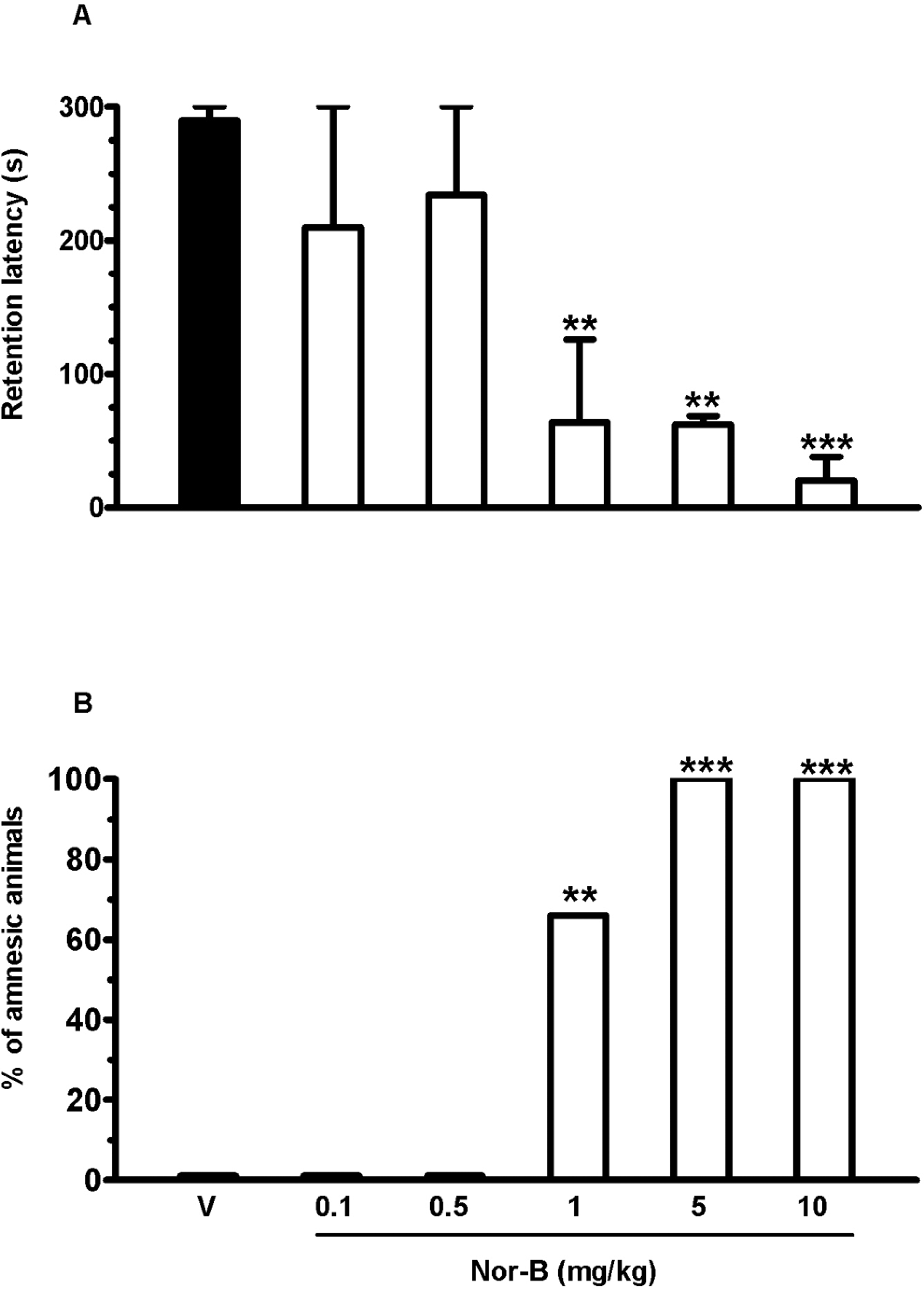
Retention latency in the passive avoidance task after nor-binaltorphimine (nor-B) administration. Nor-B was injected intraperitoneally (ip) 120 minutes before the acquisition trial. Data are expressed as the mean ± standard error of the mean (SEM) of step-through latency 24 hours after the acquisition trial (A) and as the percentage of amnesic animals (B) (10 rats per group). V indicates vehicle. **P < .01, ***P < .001 compared with vehicle group (analysis of variance [ANOVA] followed by Tukey test or Fisher exact probability test).
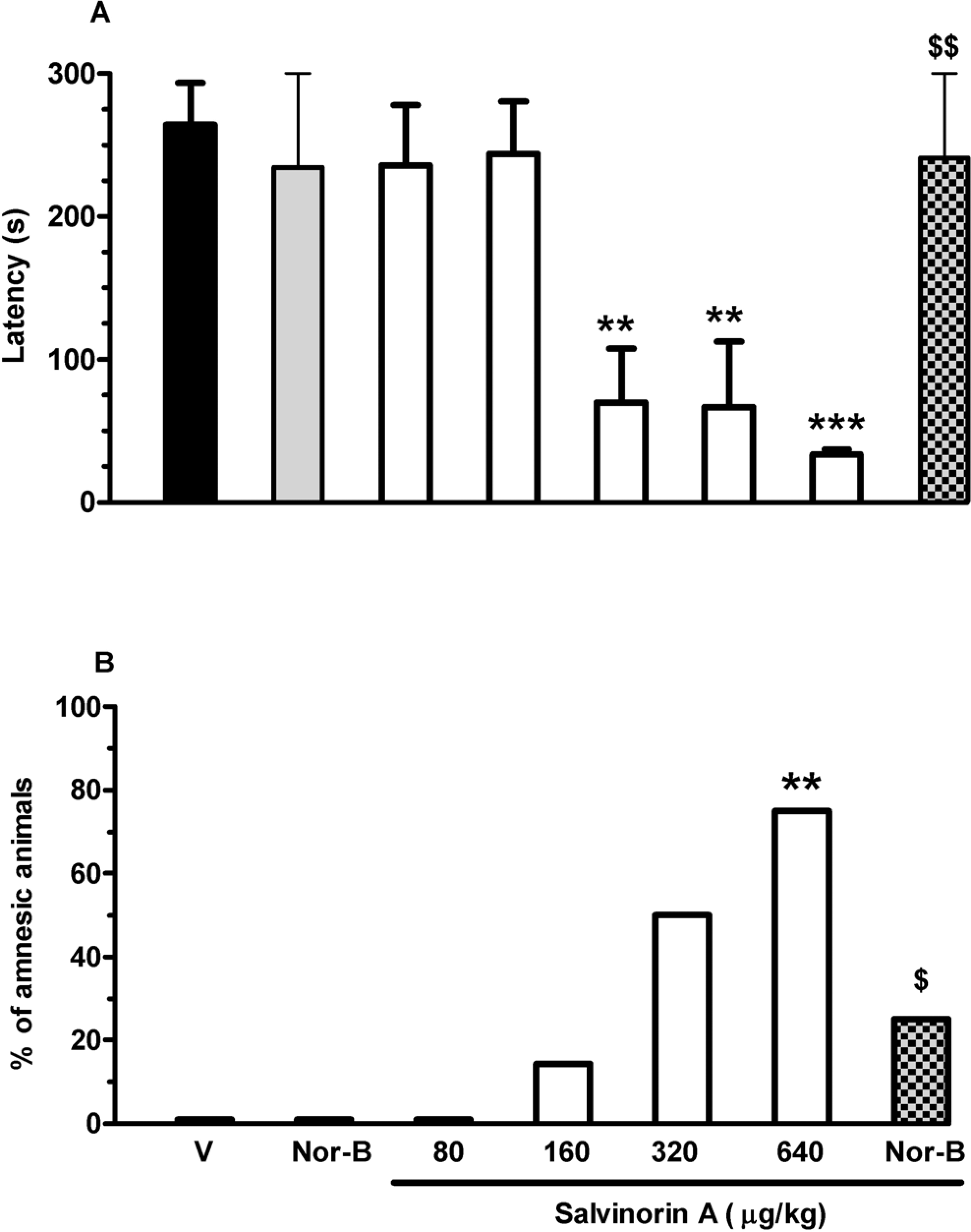
Retention latency in the passive avoidance task. Salvinorin A or vehicle were injected subcutaneously (sc), 20 minutes before the acquisition trial, alone or with nor-binaltorphimine ([nor-B] 0.5 mg/kg). Data are expressed as the mean ± standard error (SE) of step-through latency 24 hours after the acquisition trial (A) and as the percentage of amnesic animals (B) (10 rats per group). V indicates vehicle. **P < .01, ***P < .001 compared with vehicle and nor-B groups; $P < .05, $$P < .01 compared with salvinorin A (640 μg/kg) group (analysis of variance [ANOVA] followed by Tukey test or Fisher exact probability test).
Latent Inhibition
Fluid intake increased during the first 3 days, and the preexposed groups drank slightly more liquid (sucrose: 18.00 ± 2 mL) than the non-preexposed group (water: 16.45 ± 3 mL; data not shown). Importantly, however, there was no effect of salvinorin A on total fluid intake. Figure 7 clearly shows that salvinorin A only affected the preexposed groups and had no effect on the non-preexposed groups. Two-way ANOVA showed a significant main effect of treatment (P = .02) and exposure (P < .01). A Bonferroni test revealed significant differences between the non-preexposed and preexposed groups treated with vehicle, suggesting a latent inhibition. No difference was shown between the non-preexposed and preexposed groups treated with salvinorin A, indicating the lack of latent inhibition.
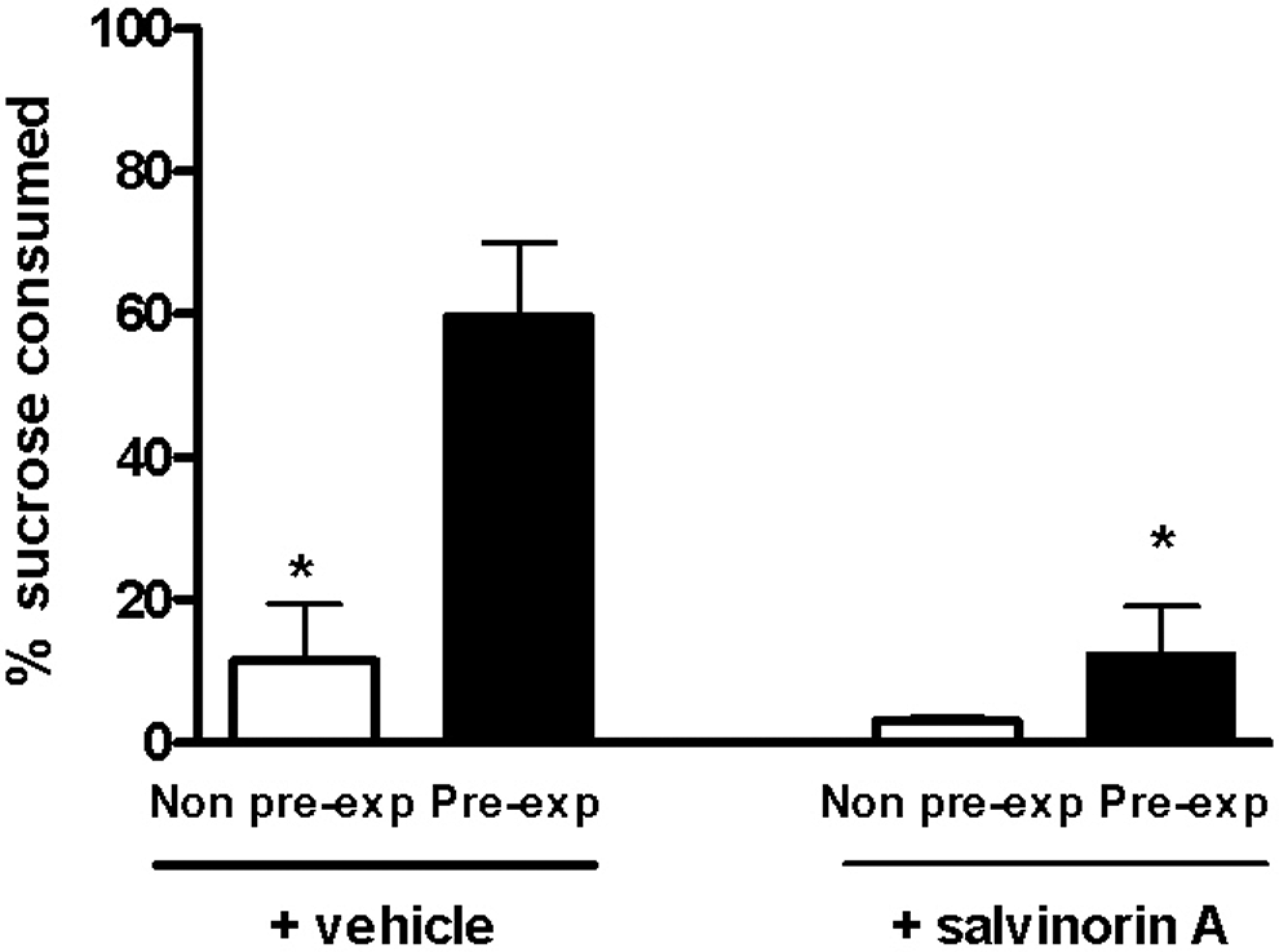
Latent inhibition. Rats were preexposed for 3 days to sucrose solution 5% (preexposed animals) or water (non-preexposed animals) and treated with salvinorin A (160 μg/kg subcutaneously [sc]) or vehicle 20 minutes before each 30-minute daily session. On the fourth day, all rats received LiCl (75 mg/kg intraperitoneally [ip]) immediately after 30 minutes of sucrose session. On the fifth day, all rats were allowed to choose between water and sucrose. Data are expressed as the mean ± standard error of the mean ([SEM] 10 rats per group). *P < .05 compared to preexposed vehicle group (analysis of variance [ANOVA] followed by Tukey test).
Discussion
The principal findings of this study are that (i) salvinorin A disrupted spatial, aversive, and episodic memory and this impairment could be due to an attention deficit; (ii) salvinorin A-induced cognitive impairment was prevented by pretreatment with the κ-opioid receptor antagonist nor-B.
We can rule out that the disruptive effects on memory were due to nonspecific motor impairment since salvinorin A up to the dose of 640 μg/kg did not affect horizontal movements in the activity cage. This lack of effect on motor activity has been reported by others who found no significant changes up to 2 mg/kg after either ip or sc injection in rats
11,48,49
and after iv injection in rhesus monkeys.
2
Inhibition of motor activity in mice, starting from 1 mg/kg ip has also been reported.
18
However, sedation-like effects were observed immediately after iv injection of 0.032 mg
Salvinorin A did not impair short-term spatial working memory. It did not alter the total number of errors, correct choices before the first error, or time taken to complete the maze, in comparison with controls. The compound significantly disrupted these behaviors only when a 2-hour delay was introduced, demonstrating impairment of long-term working memory. Since salvinorin A was given before the pre-delay, the impaired performance during the post-delay period might reflect state-dependent learning rather than specific impairment of long-term memory. However, the pattern of arm entry, a classical long-term working memory parameter, was altered with or without the delay in terms of the larger number of animals changing their patterns of arm entry. A similar disruptive pattern in the radial maze test was described in rats treated with LSD 40 and methylenedioxymethamphetamine, 38 suggesting that the disruption might be related to changes in the perception of spatial relationships, disorientation, and confusion, leading to memory disorders. 29 This is consistent with the hallucinogenic effect that was reported in young people taking S divinorum. 52
The involvement of κ-opioid system in learning and memory in the radial maze has been inadequately investigated. It has been reported that no change occurred in the number of errors and correct choices before the first error appeared in κ-opioid receptor-mutant mice, suggesting that this receptor has no role in working memory.
53
Nonetheless, our findings are consistent with what has been previously reported; whereby mice given the κ-opioid receptor agonist U-50488H directly into the CA3 hippocampal area lost an acquired ability to find a new platform in the Morris water maze task.
54
Moreover, rats trained in the same task and injected with dynorphin B in the same area also showed an impairment in spatial learning.
55
The opioid system is involved in food reward,
56
so the change in performance might have been due to the food deprivation. Therefore, we used other memory tasks that did not involve food reward. The object recognition and passive memory task results confirmed those of the 8-arm radial maze. Salvinorin A reduced step-through latency in the passive avoidance task, suggesting a deficit in aversive memory formation. κ-opioid agonists (Dyn A, B and U50488) did not consistently affect this.
57
–59
Instead, hippocampal infusion of dynorphin A (1-13) ameliorated the mecamylamine-induced learning impairment. Memory impairment detected in our studies with salvinorin A at high doses has been previously reported elsewhere
60
where the 2-day-old chicks were injected intracerebrally with either the endogenous opioid peptide dynorphin (1-13) and the highly κ-selective agonist U-50488. Furthermore, a decrease in synaptic transmission and inhibition of long-term potentiation in the basolateral nucleus of the amygdala of mice treated with U50488 has also been reported, suggesting a key role of κ-opioid receptors in memory formation.
61
Since salvinorin A has been reported to have analgesic activity,
17
the memory impairment might be due to a reduced sensitivity to the paw shock. Nevertheless, an antinociceptive effect was found in mice starting from 1 mg
Salvinorin A impaired memory through κ-opioid receptors. The highly selective κ-opioid receptor antagonist, nor-B, 68,69 fully reversed the cognitive deficit in all the tasks. However, pretreatment with the κ-opioid receptor antagonist, JDTic, blocked all of the effects of salvinorin A in the 5-choice serial reaction time task, in rats. 36
Surprisingly, nor-B, generally used to block κ-opioid receptors, 35,70 had detrimental effects in the passive avoidance test. Similarly, nor-B slightly inhibited (20%-50%) writhing in mice (evaluated in terms of number of writhes induced by an ip injection of acetic acid). 71 This agonistic effect was not blocked by naloxone, suggesting a nonopioid interaction. However, nor-B appeared to enhance the antinociceptive activity of the mu-opioid-selective agonist DAMGO ([D-Ala2, N-MePhe4, Gly-ol]-enkephalin) in the writhing test, leading to the conclusion that it had little effect on mu-opioid receptors. 71 Therefore, the detrimental effect of nor-B alone on memory might be ascribed to mu-opioid receptor activation. Alternatively, the impairment with the κ-opioid receptor antagonist might be due to its anxiogenic properties. Intracerebral infusion of nor-B has also been reported to cause anxiogenesis in the rat (in terms of decreased open-arm entries and time in the elevated plus maze task) and disrupted spontaneous alternation memory in the Y-maze. 72 Conversely, κ-opioid receptor antagonists, nor-B and JDTic, dose-dependently increased open-arm exploration in the elevated plus maze in Sprague Dawley rats 73 and reduced anxiety in mice during nicotine withdrawal. 74 Thus, activation or blockade of κ-opioid receptors can together influence anxiety and memory.
Overall, these findings in different memory tasks indicate that salvinorin A impaired long-term spatial and nonspatial memory. The mechanism by which salvinorin A affected memory remains to be clarified. Dopaminergic and noradrenergic neurotransmission are considered to play important roles in attention processes. κ-opioid receptor activation not only decreases the dopamine release in the nucleus accumbens 75 but also inhibits dopamine neurons that project from the ventral tegmental area (VTA) to the medial prefrontal cortex in rats, 76 suggesting that κ-opioid receptor agonists may disrupt performance during attention tasks by decreasing dopamine neurotransmission.
The present findings are consistent with other reports in the literature that salvinorin A, through κ-opioid receptor activation and by decreasing dopaminergic neurotransmission in the prefrontal cortex, disrupts performance in different memory tasks. Thus, overactivation of κ-opioid receptors could contribute to attention and other cognitive deficits.
Memory impairment is consistent with the reports of space–time disorientation, hallucinations, and psychotic symptoms in some people taking S divinorum 52,77,78 and are important on account of the increasing use of hallucinogenic compounds among young adults and teenagers. 11,79
Footnotes
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: partly supported by grants from Regione Lombardia (2008) to M.S.