
Abstract
Tight junction is a crucial structure in the control of paracellular transport across epithelial/endothelial barriers. This study investigated the protective effect of quercetin against hydrogen peroxide (H2O2)-induced tight junction disruption and hyperpermeability in ECV304 monolayers. Nonlethal concentration of H2O2 (100 µmol/L; 4 hours) decreased expression of the tight junction proteins zonular occudens (ZO)-1 and occludin as well as disrupted the junction structure at the cell border. Concurrently, the increased activities of extracellular signal-regulated kinase (ERK)1/2 and p38 mitogen-activated protein kinase (MAPK) were observed. Pretreatment with quercetin (10 µmol/L; 30 minutes) prior to H2O2 prevented the loss of ZO-1 and occludin. In addition, the level of phosphorylated p38 MAPK, but not that of the phosphorylated ERK1/2, decreased in quercetin-pretreated group. These findings suggested that the protective effect of quercetin involved the inhibition of phosphorylated p38 MAP activity. Furthermore, quercetin could also preserve the functional integrity of ECV304 monolayers from H2O2 exposure.
Introduction
Tight junction structure is essential for the “tight” bonding between adjacent cells of the epithelial/endothelial protective barriers. This structure is composed of integral membrane proteins such as occludin and claudin, connecting to actin cytoskeleton through zonular occudens (ZO) proteins. Its formation at the cell-to-cell contact restricts paracellular fluxes of xenobiotics and blood-borne pathogens into organ systems. 1 –3 Tight junction disruption results in barrier leakage and compromises the defense mechanism of the body. Several pathologic states including edema, jaundice, diarrhea, and blood-borne metastasis have been demonstrated to involve tight junction dysfunction. 4,5
Oxidative assaults can disrupt the integrity of tight junction structure, leading to hyperpermeability of the endothelial monolayers. 6 –9 A number of studies have demonstrated the effects of oxidative stress on the expression and/or organization of tight junction proteins including ZO-1 and occludin. 8,10 –13 For example, hydrogen peroxide (H2O2) decreased the expression of occludin in bovine pulmonary artery endothelial cells, leading to barrier leakage and the loss of transendothelial electrical resistance values. 14,15 The tight junction damage has been linked to certain members of mitogen-activated protein kinases (MAPKs), especially extracellular signal-regulated kinase (ERK or p44/42 MAPK), c-Jun-NH2-terminal kinase (JNK), and p38 kinase. 6,15 –17 It has been shown that the redistribution of occludin in H2O2-induced hyperpermeability of endothelial barriers involved the phosphorylation of ERK1/2. 15,18 The presence of PD98059, a specific ERK1/2 inhibitor, in human umbilical vein endothelial cell (HUVEC) monolayers treated with H2O2 suppressed the hyperpermeability and enhanced the localization of ZO-1 and occludin at the cell surface. 15 Moreover, p38 MAPK was also suggested to be responsible for the H2O2-mediated dysfunction of the microvascular endothelial barriers. 19 Treatment with SB202190, a p38 inhibitor, alleviated an increase in the solute permeability in H2O2-treated HUVEC monolayers. 7
Quercetin is one of the most widely distributed flavonoids and can be found abundantly in fruits, vegetables, and beverages such as apple, onion, tea, and red wine. 20 Similar to other flavonoids, quercetin displays a broad spectrum of pharmacological actions including antioxidant, 21 –23 anti-inflammatory, 22 –26 cardioprotective, 21 and antiproliferative activities. 24,27 –29 Recently, quercetin has been shown to enhance the barrier function of the Caco-2 monolayers through increased expression of tight junction proteins, including occludin and claudin, as well as their assembly. 30,31 The molecular mechanisms of quercetin have been linked to its interaction with a number of cellular targets including protein kinase C (PKC) and MAPK, causing changes in downstream signaling processes. 21,32
In this study, we investigated the protective effect of quercetin on the H2O2-induced disruption of tight junction structure in the ECV304 monolayers. H2O2 at nonlethal concentration was used to disturb junction assembly and its functional integrity. The potential inhibitory effect of quercetin on the MAPK pathways might enable this flavonoid to preserve the expression and localization of tight junction proteins under H2O2-induced stress condition.
Materials and Methods
Chemicals and Reagents
Culture medium was purchased from GIBCO Life Technologies, Inc (Grand Island, New York). Quercetin, H2O2, penicillin G sodium, streptomycin sulfate, dimethyl sulphoxide (DMSO), and 3-[4,5-dimethylthiazol-2-yl]-2,5-diphenltetrazolium bromide (MTT) were purchased from Sigma Chemical Company (St Louis, Missouri). All other chemicals and reagents were reagent grade or the highest grade commercially available.
On the day of experiment, quercetin was freshly dissolved in 100% DMSO and further diluted with culture medium. The final concentration of DMSO in each experiment was less than 0.5%. In addition, the solvent control was also conducted in each experiment. H2O2 was diluted with double-distilled water to the desired concentration at the beginning of each experiment.
Antibodies
Rabbit monoclonal anti-ZO-1 and anti-occludin antibodies were purchased from Zymed (San Francisco, California). Rabbit polyclonal anti-MAP kinase ERK1/ERK2, antiphosphorylated MAP kinase ERK1/ERK2 (p-Thr-202/p-Tyr-204), anti-p38 MAP kinase, and anti-phosphorylated p38 MAP kinase (p-Thr-180/p-Tyr-182) were purchased from Calbiochem (San Diego, California). Mouse monoclonal immunoglobulin G (IgG) anti-β-actin antibody was purchased from Santa Cruz Biotechnology, Inc (Santa Cruz, California). Horseradish peroxidase (HRP)-conjugated anti-(rabbit-IgG) antibody was purchased from Cell Signaling Technology (Beverly, Massachusetts). Dylight 680 labeled-anti-(mouse-IgG) antibody was purchased from KPL, Inc (Gaithersburg, Maryland). Cy-3 conjugated anti-(rabbit-IgG) antibody was from Sigma Chemical Company.
Cell Culture
ECV304 cells, obtained from Cell Line Service (Eppelheim, Germany), were cultured in M199 supplemented with 10% fetal bovine serum and 1% penicillin–streptomycin at 37°C in 5% CO2 air atmosphere. The culture medium was replaced every 48 hours. The cells were subcultured once a week with a split ratio of 1:3.
Cytotoxicity Studies
Cell viability was determined using the MTT reduction assay. 33,34 ECV304 cells (2 × 105 cells/mL) were seeded in 96-well plates and cultured for 24 hours prior to treatment with either quercetin or H2O2. After treatment, the medium in each well was replaced with 100 µL of MTT solution (0.4 mg/mL) and incubated in a 5% CO2 incubator at 37°C for 4 hours. The intracellular formazan crystals were solubilized with 100 µL of DMSO and quantified spectrophotometrically at a measuring wavelength of 570 nm and a reference wavelength of 620 nm (Anthos Labtec HT2 microplate reader, Anthos Labtec Inst, Salzburg, Austria).
Treatment With H2O2 and Quercetin
For the immunofluorescence microscopy and Western blot analysis, the cells were seeded in 24-well plates at 2 × 105 cells/well and grown to confluency (48 hours). For measurement of transepithelial electrical resistance (TEER) and the permeability assay, the cells (2 × 105 cells/well) were grown on polycarbonate membranes in Transwell (0.4 µm pore size; 12 mm diameter; Costar, Cambridge, Massachusetts). Confluent monolayers of ECV304 cells were treated with quercetin 10 µmol/L for 30 minutes and washed once with medium prior to addition of H2O2 (100 µmol/L) for 4 hours.
Immunofluorescence Microscopy
After treatment, the cells were washed twice with ice-cold phosphate-buffered saline (PBS) and fixed with the ethanolic solution containing 5% acetic acid at −20°C for 10 minutes. The cells were permeabilized with 0.05% Triton X-100 in PBS for 10 minutes. After washing 3 times with ice-cold PBS, the cells were treated with 3% bovine serum albumin for 15 minutes, followed by incubation with primary antibodies against occludin (1:200) or ZO-1 (1:200) for 1 hour. The cells were then washed 3 times with PBS and incubated for 1 hour with Cy-3 conjugated secondary antibodies, followed by fluorescence microscopy (BX-FLA, Olympus, Japan).
Western Blot Analysis
Cell lysates were prepared by lysing the cells in the ice-cold lysis buffer (20 mmol/L Tris–HCl [pH 7.4], 150 mmol/L NaCl, 1% Triton X-100, 1 mmol/L Pefabloc, 1% Na deoxycholate, 10 mmol/L NaF, 1 mmol/L Na orthovanadate, 5 µg/mL aprotinin). Protein concentrations were quantified by Bradford assay (Bio-Rad Laboratories, Glattbrugg, Switzerland). Protein samples (30 µg per lane) were loaded onto 8% or 12% sodium dodecyl sulfate (SDS)-polyacrylamide gels and separated by electrophoresis. The proteins were then electrically transferred to polyvinylidene fluoride (PVDF) membranes and blotted for occludin, ZO-1, ERK, p-ERK, p38, and p-p38 with the use of specific primary antibodies at the dilution of 1:1000. Subsequently, the blots were incubated with HRP-conjugated anti-rabbit IgG secondary antibodies (1:2000) and developed using the enhanced chemiluminescence (ECL) detection system (Supersignal, West Pico, Pierce, Illinois). In these experiments, β-actin was also quantified as an internal standard for protein loading. Densitometry measurements on ECL films were performed using Image J program (NIH, Bethesda, Maryland).
Measurement of TEER
The electrical resistance across the monolayer was measured with a chopstick-like electrode connected to a Millicell-ERS (electrical resistance system; Millipore, Bedford, Massachusetts). Transepithelial electrical resistance value was calculated by multiplying the measured electrical resistance (Ω) with the surface area of the monolayer (cm2). Generally, TEER values of the “tight” ECV304 monolayer varied from 85 to 120 Ω cm2. 35 In this study, TEER was monitored in the confluent ECV304 monolayers every 2 days. The apparent “tight” barriers were considered when TEER reached its plateau at maximal values of at least 90 Ω cm2 (usually at 10-12 days). The “tight” monolayers were then ready for H2O2 treatment as indicated above. Changes of TEER values were calculated as a percentage of the baseline values obtained prior to treatment. In this study, duplicate cell monolayers were used for each group, and the experiment was conducted at least 4 times, repetitively.
Permeability Assay
ECV304 cells were cultured in Transwell insert to postconfluency (10-12 days) prior to treatment as indicated above. For permeability assay, the medium in the apical chamber was replaced with 0.5 mL of medium containing 50 µmol/L phenol red and that in the basolateral chamber was replaced with 1.6 mL of the phenol red–free medium. After incubation at 37°C for 1 hour, a 100 µL aliquot of medium in the basolateral chamber was taken to measure the amount of phenol red spectrophotometrically at the wavelength of 430 nm.
Statistical Analysis
Data are reported as means ± SEM of at least 3 separate experiments. The significant differences were tested by either Student t test or analysis of variance (ANOVA) with subsequent Scheffe post hoc analysis, where appropriate. Differences were considered statistically significant at P < .05.
Results
Cell Viability
In this study, both H2O2 (100 µmol/L) and quercetin (10 µmol/L) had no significant effects on the viability of ECV304 cells (data not shown).
Immunofluorescence Microscopy of Tight Junction Proteins ZO-1 and Occludin
Although the treatment of ECV304 with H2O2 (100 µmol/L) for 4 hours had no effect on cell viability, it induced the disturbance of tight junction structure and the loss of the organization of its major protein components, specifically ZO-1 and occludin, as measured by immunofluorescence microscopy. Under the normal circumstance, the ECV304 monolayers expressed the continuous distribution pattern of ZO-1 (Figure 1A) and occludin (Figure 1B) proteins at the cell boundaries. In addition, quercetin had no influence on the localization of these tight junction proteins (Figures 1C and 1D). Treatment of the monolayers with H2O2 (100 µmol/L) apparently changed the pericellular distribution pattern of both proteins (Figures 1E and 1F), suggesting the abnormality of localization and the possible loss of expression of these tight junction proteins. Pretreatment of the monolayers with quercetin (10 µmol/L) for 30 minutes prior to H2O2 was able to restore the complete circumferential expression of both tight junction proteins at the cell border (Figures 1G and 1H). Hence, our results demonstrated the protective effects of quercetin on H2O2-mediated disruption of tight junction proteins at cell-to-cell contact sites.
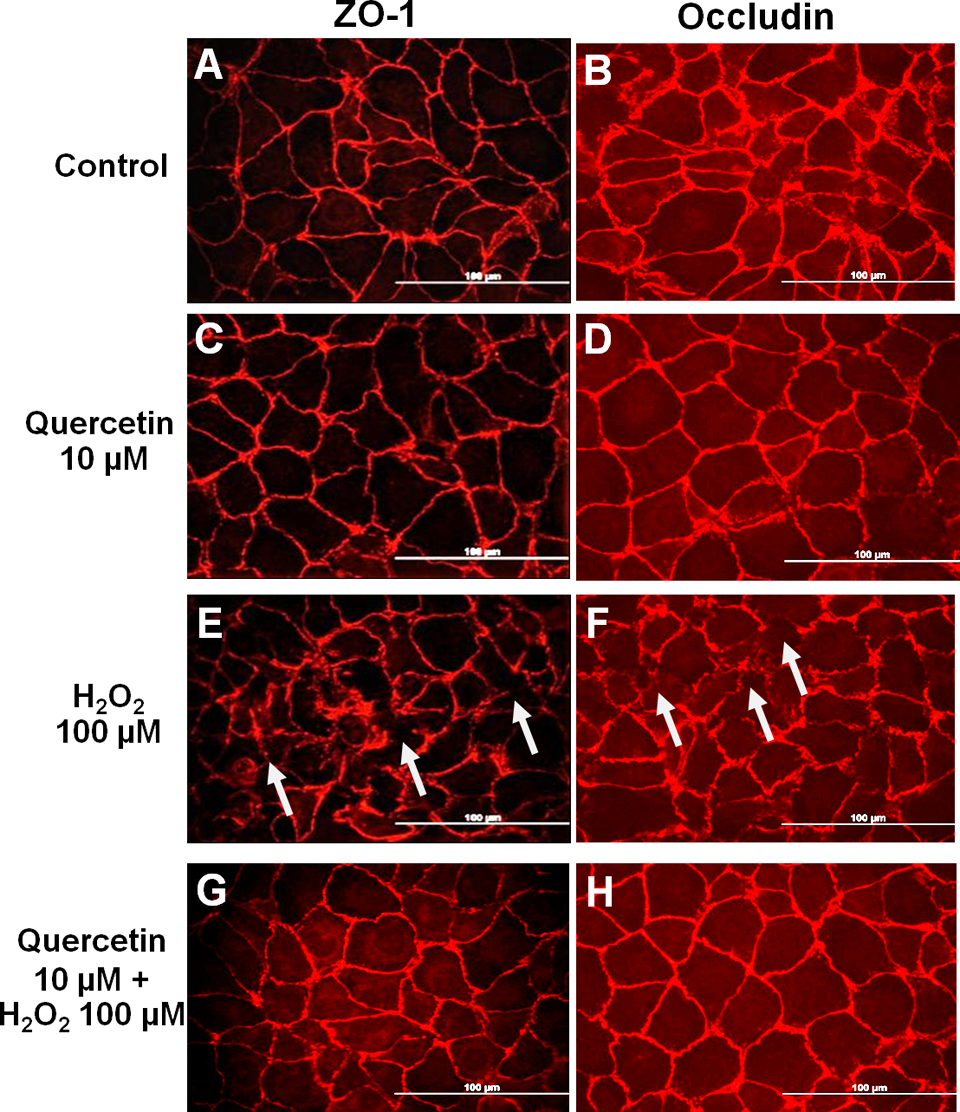
Immunofluorescent staining showing localization of zonular occudens (ZO)-1 and occludin proteins in ECV304 monolayers. A and B, control. C and D, quercetin 10 µmol/L for 30 minutes. E and F, H2O2 100 µmol/L for 4 hours, arrow showed the disruptions in the expression of ZO-1 and occludin. G and H, quercetin 10 µmol/L for 30 minutes followed by H2O2 100 µmol/L for 4 hours. All fields were shot at ×40. Bar = 100 µm.
Expression of ZO-1 and Occludin Proteins
Levels of ZO-1 and occludin expression were then examined with Western blot analyses. As shown in Figure 2, H2O2 (100 µmol/L) significantly suppressed ZO-1 and occludin levels by approximately 30%. Pretreatment of the cells with quercetin for 30 minutes prior to H2O2 was able to significantly preserve at least 90% of the ZO-1 and occludin expression.
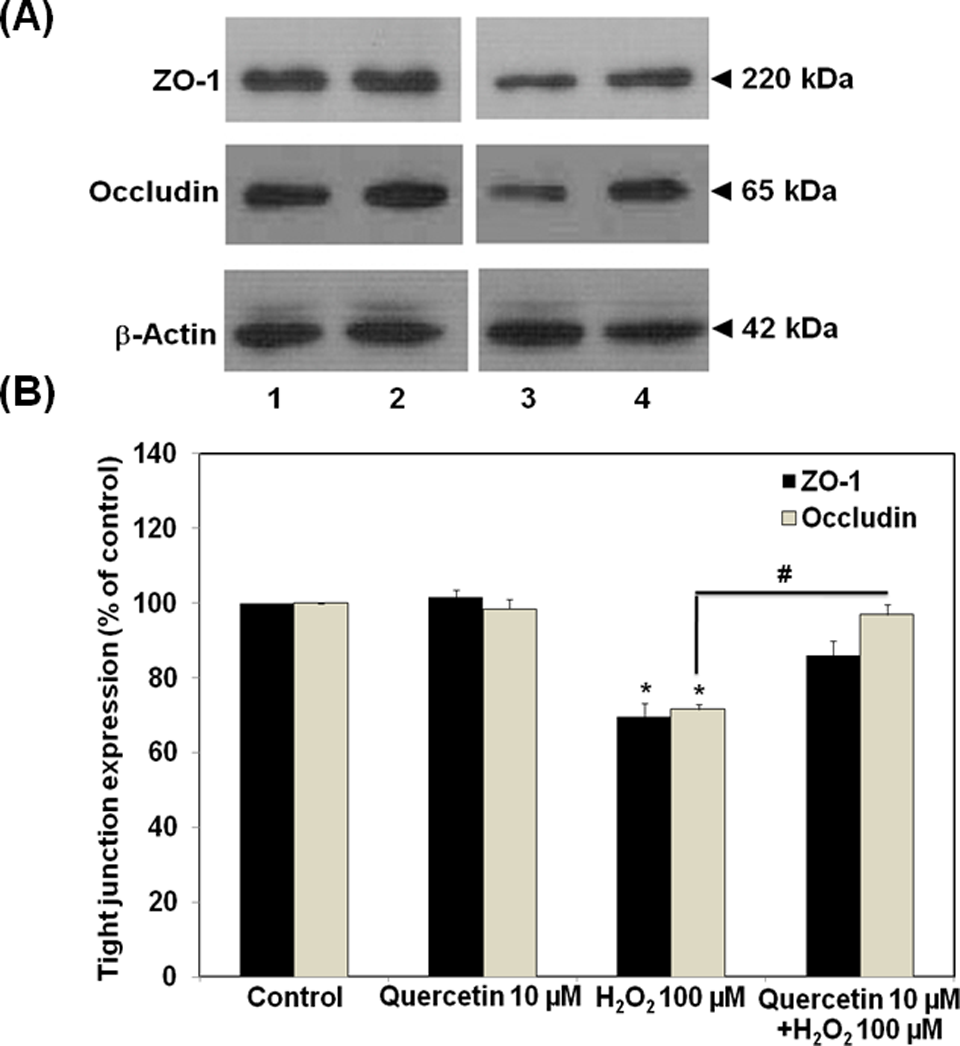
Effects of quercetin on the expression of zonular occudens (ZO)-1 proteins and occludin in ECV304 monolayers. A, Cell lysate were run on 8% (for ZO-1) and 12% (for occludin and β-actin) sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE). B, Densitometrical analysis of ZO-1 proteins and occludin (lane 1, control; 2, quercetin 10 µmol/L; 3, H2O2 100 µmol/L for 4 hours; 4, quercetin 10 µmol/L for 30 minutes followed by H2O2 100 µmol/L for 4 hours). Data are representative of 4 separate experiments. *P < .05, compared with untreated group (control); #P < .05, compared with H2O2 alone group.
Mitogen-Activated Protein Kinase Expression Analysis
We further investigated the involvement of MAPKs in the protective effect of quercetin against tight junction disruption. Several previous reports suggested the activation of ERK1/2 and/or p38 MAP kinase pathways as the underlying mechanism of H2O2-mediated hyperpermeability of endothelial monolayers. 6,7,15 On H2O2 treatment even at low concentration of 100 µmol/L, we detected the increase in ERK1/2 and p38 phosphorylation. These findings agreed with others that H2O2 increased the activities of both ERK1/2 and p38 MAP kinases in the cells. Quercetin (10 µmol/L) had no effects on the levels of phosphorylated forms of ERK1/2 and p38 MAP kinases. However, quercetin given 30 minutes prior to H2O2 affected these 2 MAP kinase activities in an opposite way. As shown in Figures 3 and 4, quercetin significantly enhanced the effect of H2O2 on ERK1/2 phosphorylation, whereas it suppressed the effect of H2O2 on the activation of p38 MAP kinase. The phosphorylated p38 level in quercetin pretreatment group was expressed at the lesser degree than that observed in the H2O2-treated group without quercetin pretreatment (Figure 4).
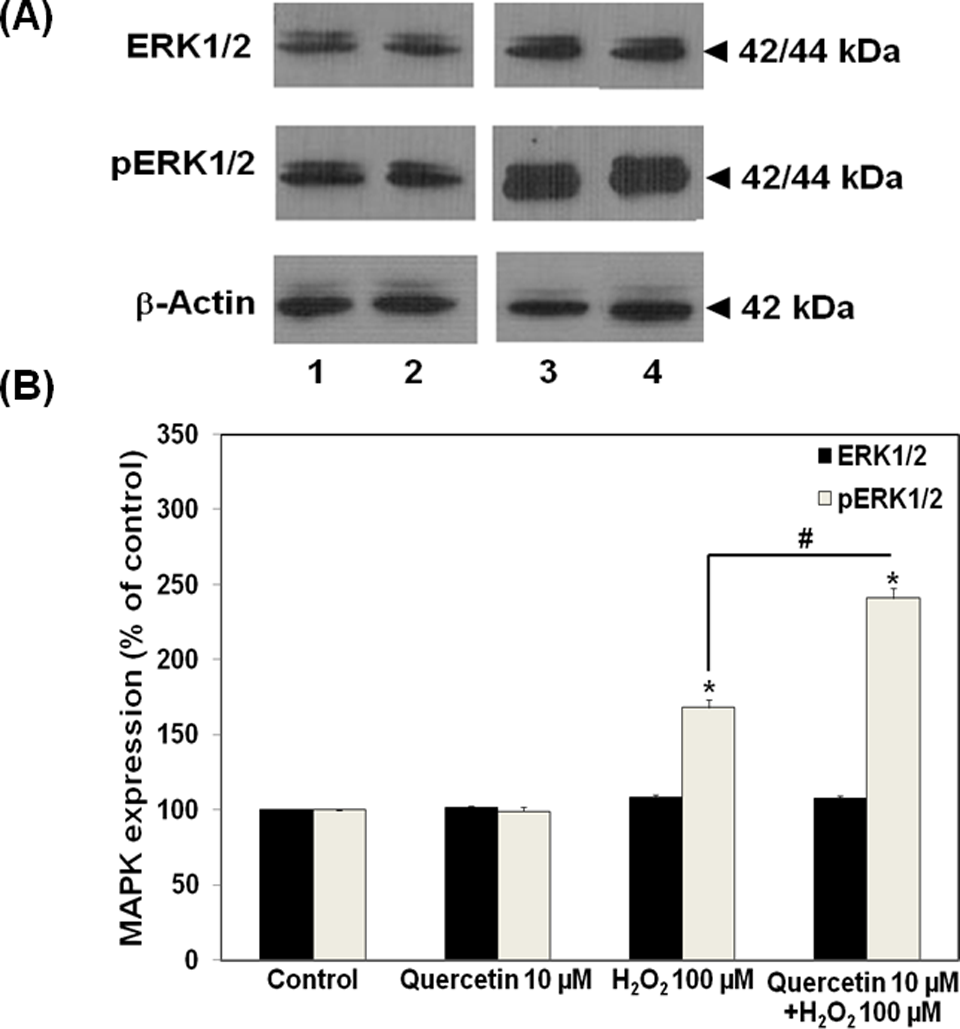
Effects of quercetin on expression of extracellular signal-regulated kinase (ERK)1/2 and its phosphorylated form in ECV304 monolayers. A, Immunoblots of ERK1/2 and phosphorylated ERK1/2. B, Densitometrical analysis of immunoblots of ERK1/2 and phosphorylated ERK1/2 (lane 1, control; 2, quercetin 10 µmol/L; 3, H2O2 100 µmol/L for 4 hours; 4, quercetin 10 µmol/L for 30 minutes followed by H2O2 100 µmol/L for 4 hours). *P < .05, compared with untreated group (control); #P < .05, compared with H2O2 alone group.
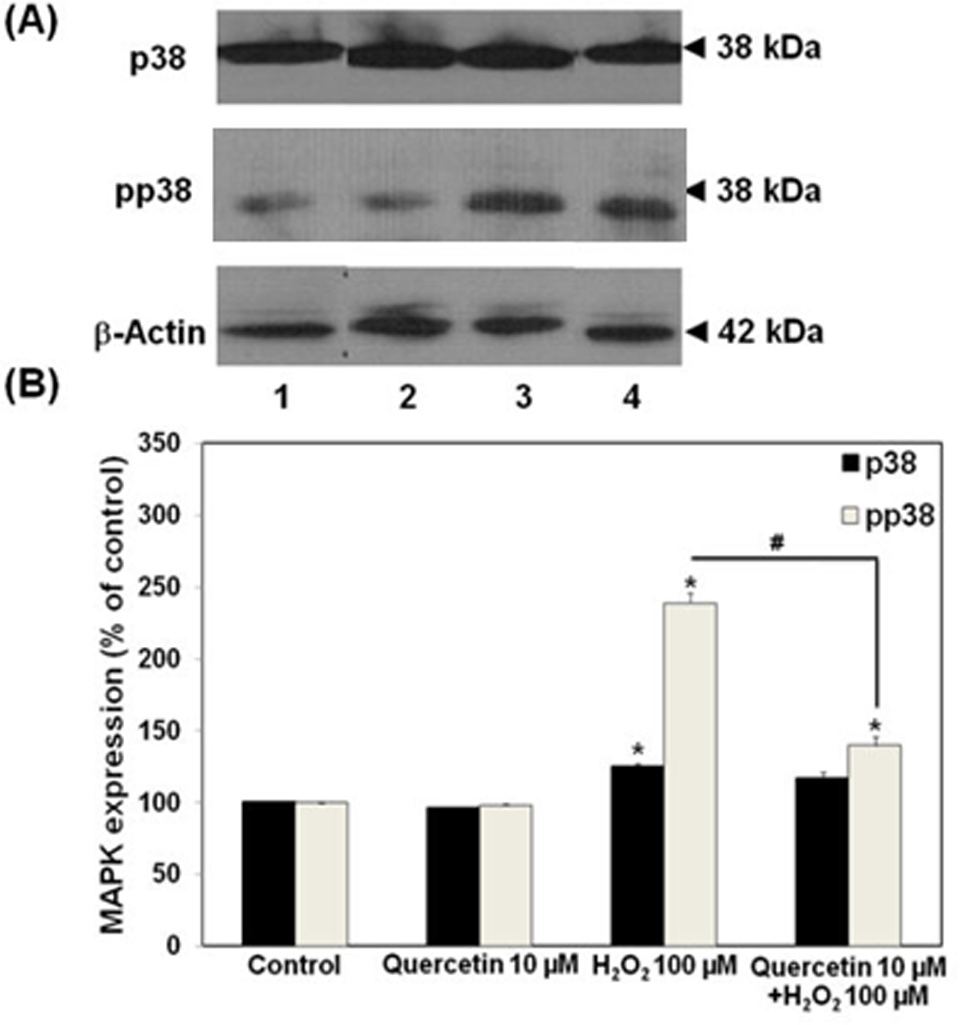
Effects of quercetin on expression of p38 and its phosphorylated form in ECV304 monolayers. A, Immunoblots of p38 and phosphorylated p38. B, Densitometrical analysis of immunoblots of p38 and phosphorylated p38 (lane 1, control; 2, quercetin 10 µmol/L; 3, H2O2 100 µmol/L for 4 hours; 4, quercetin 10 µmol/L for 30 minutes followed by H2O2 100 µmol/L for 4 hours). *P < .05, compared with untreated group (control); #P < .05, compared with H2O2 alone group.
Functional Analysis With TEER Measurement and Permeability Assay
The ability of quercetin to preserve the function of tight junction structure during H2O2 exposure was also tested using TEER measurement and permeability assay. Tightness of the junction would result in high membrane electrical resistance and high restrictiveness to paracellular movement of phenol red across the monolayer. In this study, H2O2 (100 µmol/L) impaired the integrity of tight junction structure of the ECV304 monolayers after 4-hour exposure time, as evidenced by significant reduction in TEER values (Figure 5) and increase in phenol red permeability (Figure 6). In comparison with the control group, H2O2 significantly decreased the TEER values by approximately 20%, whereas it increased the paracellular permeability of phenol red by 2-fold. Pretreatment of the monolayers with quercetin (10 µmol/L) for 30 minutes prior to H2O2 exposure attenuated the effect of H2O2 on TEER reduction (Figure 5), and prevented the H2O2-induced hyperpermeability (Figure 6). These observations suggested the intrinsic protective activity of quercetin against the loss of tight junction integrity upon H2O2 exposure.
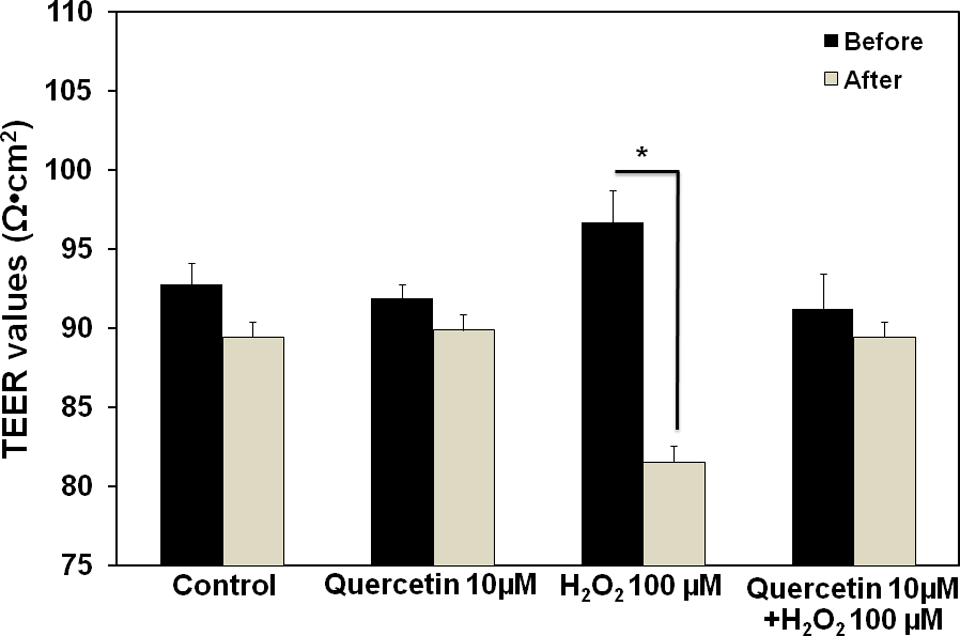
Effects of quercetin and H2O2 on the transepithelial electrical resistance (TEER) values of ECV304 monolayers (12-day postconfluency). The ECV304 monolayers were pretreated with quercetin (10 µmol/L; 30 minutes), followed by H2O2 (100 µmol/L; 4 hours). The TEER values were measured as described in Materials and Methods. Data are presented as means ± SEM obtained from 4 separate experiments. *P < .05, compared with the values before treatment.
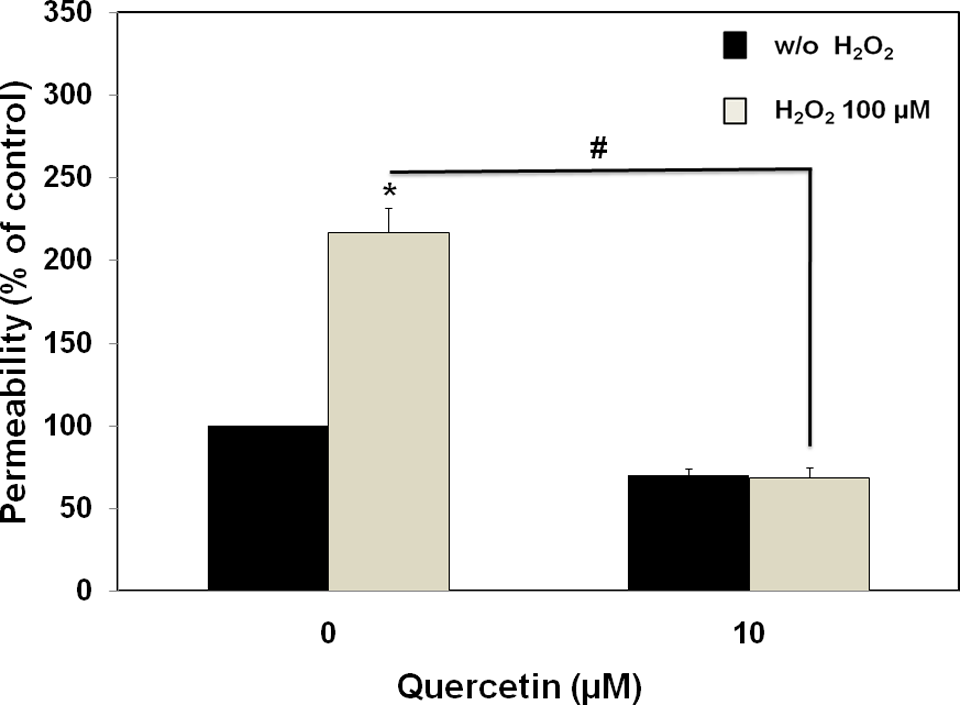
Effect of quercetin on H2O2-induced hyperpermeability of ECV304 monolayers (12-day postconfluency). The ECV304 monolayers were pretreated with quercetin (10 µmol/L; 30 minutes), followed by H2O2 (100 µmol/L; 4 hours). At the end of incubation, the phenol red permeability assay was performed as described in Materials and Methods. Data were obtained from 4 separate experiments and expressed as means ± SEM of the percentage of the untreated group (control). *P < .05, compared with control cultures; #P < .05, compared with H2O2 alone group.
Discussion
Our data demonstrated the protective effect of quercetin against H2O2-induced tight junction disruption and hyperpermeability. To investigate the integrity and function of tight junction complexes upon oxidative assault, the ECV304 cells at 2 different postconfluency periods were used as a model of epithelial barriers. At 2-day postconfluency, the expression and localization of ZO-1 and occludin were observed at pericellular circumference, indicating the presence of tight junction structure. However, it took at least 10 to 12 days postconfluency for these tight junctions to fully develop their “mature” fencing activities, as evidenced by the stable high TEER values and low phenol red permeability. Regardless of the postconfluency period, the ECV304 viability was not affected by 4-hour exposure to H2O2 (100 µmol/L).
The activation of MAP kinase pathways has been well corroborated with the loss of normal expression and localization of junctional proteins, especially ZO-1 and occludin, in H2O2-induced tight junction disruption and hyperpermeability. 6,8,13,15,36,37 For example, Kevil et al reported that 3-hour exposure of HUVEC to H2O2 (500 µmol/L) resulted in hyperpermeability and disorganization of ZO-1 and occludin at the cell-to-cell contact sites, which occurred in concurrent with ERK1/2 activation. 15 We demonstrated that exposure of the ECV304 monolayers (2-day postconfluency) to H2O2 for 4 hours led to a decrease in the expression and localization of ZO-1 and occludin proteins, as evidenced by our findings from the Western blotting analysis and the discontinuous pattern of immunofluorescent staining for these 2 junctional proteins. In addition, the increases in phosphorylated ERK1/2 and p38 were observed concomitantly.
Pretreatment of the ECV304 monolayers with quercetin for 30 minutes was able to prevent a significant loss of ZO-1 and occludin expression upon challenging with H2O2. The antiactive MAP kinase antibodies revealed that an increased p38 activity resulting from H2O2 exposure for 4 hours could be prevented by quercetin. On the contrary, quercetin appeared to potentiate H2O2-induced stimulation of ERK1/2 activity. There were different viewpoints about the involvement of p38 and ERK1/2 activations in H2O2-induced hyperpermeability. Some studies suggested that ERK1/2 phosphorylation was a key regulatory pathway in the maintenance of tight junction integrity against H2O2 treatment. 15,38,39 On the contrary, there have been other reports suggested that p38 MAPK, but not ERK, was involved in the barrier dysfunction caused by H2O2. 19 These differences could involve several factors including cell type, culture condition, and time to exposure. From this study, it was likely that the activation of p38 was responsible for the downregulation of ZO-1 and occludin in the H2O2-treated ECV304 monolayers.
We subsequently investigated the ability of quercetin to preserve the barrier function of the ECV304 monolayers against H2O2 treatment. The tight junction structure was very important in the control of paracellular flux of solute through the endothelial or epithelial monolayers. In general, the functional expression of tight junctions could be assessed by the low-level transport of phenol red across the monolayer, which occurred concurrently with the high TEER values. 40 Hence, the dysfunction of tight junction complexes could be reflected by the reduction in electrical resistance (or TEER) and the increase in paracellular transport. 41 In this study, ECV304 cells grown as monolayer exhibited increasing TEER values over time, with the stable maximal values of 95.3 ± 8.36 Ω cm2 at 12 days postconfluency. At this state, tight junctions of the ECV304 monolayers fully expressed their functional activities. Exposure of ECV304 cells to H2O2 resulted in increased paracellular permeability along with decreased electrical resistance across barriers. These findings were in agreement with other reports on the effects of H2O2 on barrier function. 39,42,43 Transient treatment of the ECV304 monolayers with quercetin for 30 minutes could prevent barrier dysfunction caused by H2O2, suggesting the intracellular actions of this flavonoid in maintenance of tight junction integrity.
In conclusion, pretreatment of the ECV304 monolayers with quercetin prior to H2O2 exposure enabled the cells to preserve the expression and localization of tight junction proteins occludin and ZO-1 at the pericellular circumference. It was possible that quercetin exerted its protective action through the suppression of MAP kinase activities, especially p38 MAPK. In addition, quercetin was able to prevent the H2O2-induced hyperpermeability, presumably through maintaining the integrity of its tight junction structures.
Footnotes
The author(s) declared no conflicts of interest with respect to the authorship and/or publication of this article.
The author(s) disclosed receipt of the following financial support for the research and/or authorship of this article: the 90th Anniversary of Chulalongkorn University Fund (Ratchadaphiseksomphot Endowment Fund), Chulalongkorn University, Bangkok, Thailand.