
Abstract
Collectively, retinal diseases, including age-related macular degeneration, retinitis pigmentosa, and diabetic retinopathy, result in severe vision impairment worldwide. The absence and/or limited availability of successful drug therapies for these blinding disorders necessitates further understanding their pathobiology and identifying new targetable signaling pathways. Nuclear receptors are transcription regulators of many key aspects of human physiology, as well as pathophysiology, with reported roles in development, aging, and disease. Some of the pathways regulated by nuclear receptors include, but are not limited to, angiogenesis, inflammation, and lipid metabolic dysregulation, mechanisms also important in the initiation and development of several retinal diseases. Herein, we present an overview of the biology of three diseases affecting the posterior eye, summarize a growing body of evidence that suggests direct or indirect involvement of nuclear receptors in disease progression, and discuss the therapeutic potential of targeting nuclear receptors for treatment.
Introduction
The human nuclear receptor (NR) superfamily is composed of 48 evolutionarily related transcription factors, which respond to endogenous ligands, including steroid hormones, fatty acids, bile acids, lipophilic vitamins, and cholesterol metabolites, and exogenous ligands, such as drugs and toxins.1-4 Functionally, NRs are critical regulators of a wide range of physiologic and developmental pathways. The myriad molecular pathways modulated by NRs include, but are not limited to, inflammation, lipid metabolism, apoptosis, extracellular matrix regulation, energy metabolism, and angiogenesis.5-7 These pathways are of particular interest to vision scientists, as these functions are also compromised in several retinal diseases, such as diabetic retinopathy (DR), age-related macular degeneration (AMD), and retinitis pigmentosa (RP)8-11 ( Fig. 1 ). Treatment options currently available for the above-listed vision-impairing retinal diseases are either quite limited or unavailable. This unmet need classifies identification of new signaling pathways that contribute to initiation or progression of retinal diseases and potential therapeutic targets as a top priority. Of relevance is that many members of the NR superfamily have well-characterized ligands and serve as therapeutic targets for the development of drugs to treat diseases including cancer, diabetes, atherosclerosis, inflammation, and endocrine disorders.5,6,12,13 Accordingly, identification of NR-driven pathways altered in retinal diseases, for which there are drugs already available, would be highly advantageous to potentially decreasing the timeline to treatment in man for ocular indications. In light of the overlap between pathogenic pathways of several retinal disorders with other NR-regulated diseases, herein we provide an overview of a growing body of evidence that suggests NRs may play an overlooked role in the development of pathologies associated with retinal diseases. We begin with a brief overview of the NR superfamily. We follow with describing the known pathobiologies associated with several diseases affecting the posterior eye, including RP, DR, and AMD. Finally, we summarize recent progress in identifying the contribution of various NRs, either directly or indirectly, to the biology of these diseases. Collectively, these studies provide promising support for consideration of therapeutic targeting of this class of receptors for treatment of various retinal disorders.
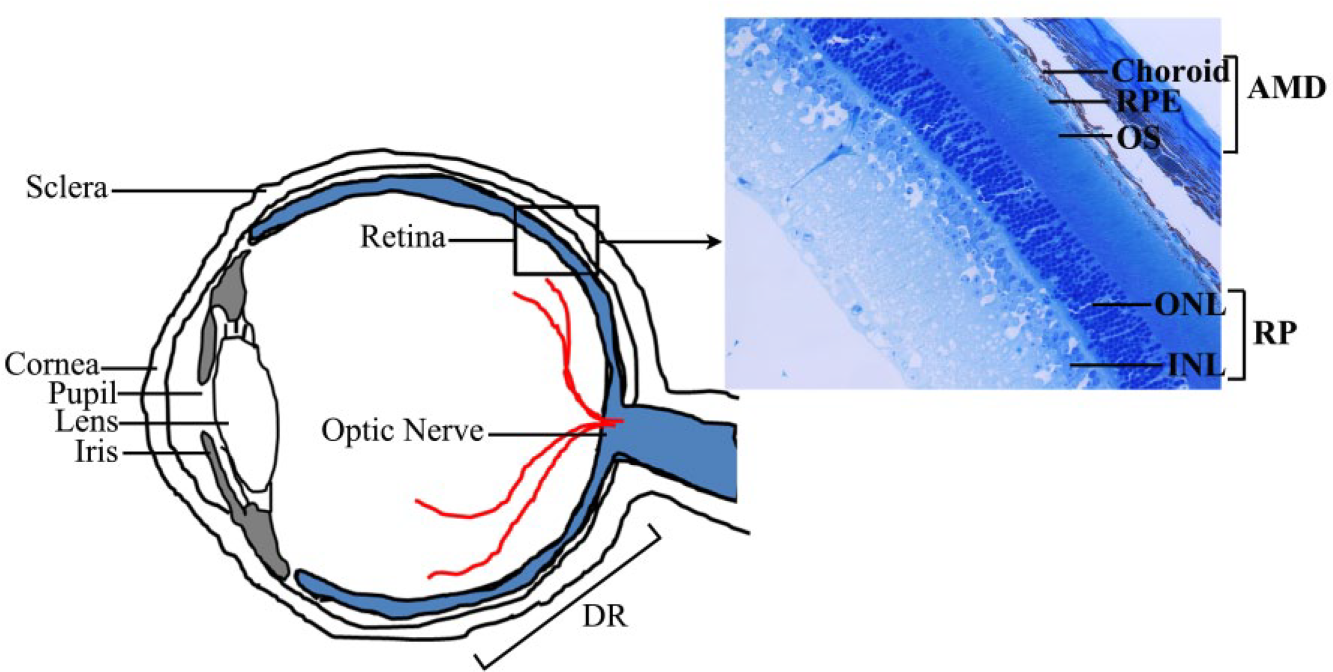
Schematic view of a cross section of the human eye highlighting the various cell types affected in the retinal diseases AMD, RP, and DR. INL, inner nuclear layer; ONL, outer nuclear layer; OS, outer segment; RPE, retinal pigment epithelium.
Nuclear Receptor Superfamily
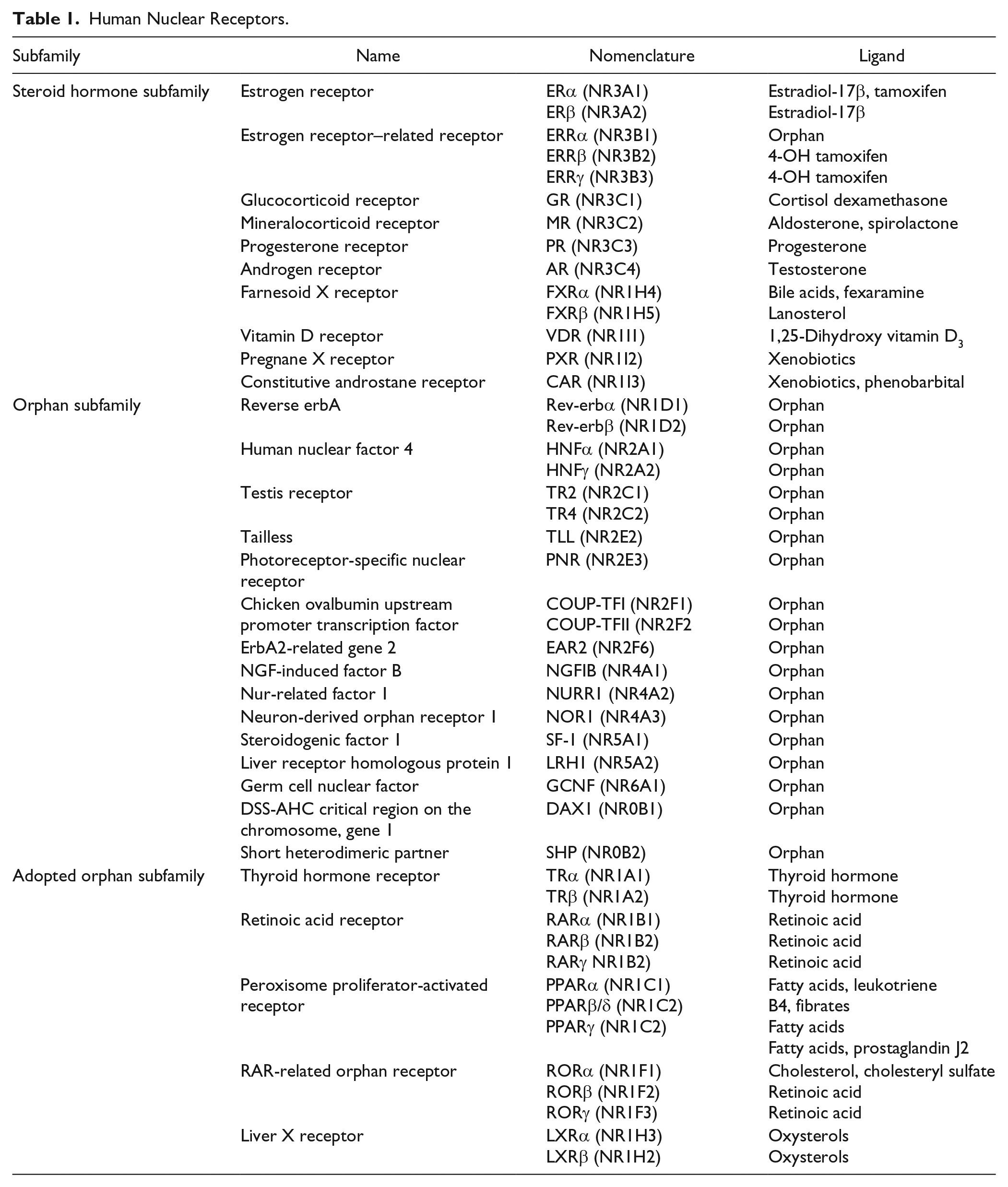
NRs are primarily defined as ligand-regulated transcription factors that modulate target gene expression. The first NRs cloned were the glucocorticoid receptor (GR; NR3C1) and the estrogen receptor (ERα; NR3A1) in 1985 and 1986, respectively.14,15 This was closely followed by the discovery of receptors for mineralocorticoids, progesterone, androgens, and fat-soluble vitamins A and D. 4 Comparison of sequences led to identification of an evolutionary conserved template and domain structure for the members of the NR superfamily. Overall, the conserved NR structure is modular and consists of six subregions ( Fig. 2A , domains A–F). However, the sequence homologies of these subregions vary within the NR family.1,13 The amino terminus (also known as the A/B region) is the most divergent among receptors, and it contains a hormone-independent transactivation domain known as activation function 1 (AF-1), which is recognized by coactivators and/or other transcription factors. The region C is the most conserved domain, has two zinc-finger motifs, and contains a DNA-binding domain (DBD). Noteworthy are two exceptions: the dosage-specific sex reversal-adrenal hypoplasia congenital critical region on the X chromosome (DAX1; NR0B1) and short heterodimeric partner (SHP; NR0B2), both of which lack the DBD. The D region, which has a low degree of conservation, forms a “hinge domain,” as it is localized between the DBD and the ligand-binding domain (LBD). The E region contains the ligand-dependent transactivation domain (AF-2) and is well conserved between the various family members, but diverges sufficiently to ensure ligand selectivity. The F region is further toward the carboxyl terminus, present in only a few receptors to date, and its role remains unclear in NR function. Overall, the NR superfamily can functionally be categorized into three main groups on the basis of their ligand- and DNA-binding properties. 16 The NRs of the first category, the classical steroid hormone receptors, activate transcription by binding to the response element located within the promoter region of their target genes ( Fig. 2B ). The second category of NRs is the orphan receptors. They are named as such because they have the conserved domain structure of NRs, but their endogenous ligands have not been identified to date. Orphan receptors for which the ligand has been identified make up the third family of NRs and are referred to as adopted orphan receptors ( Table 1 ).
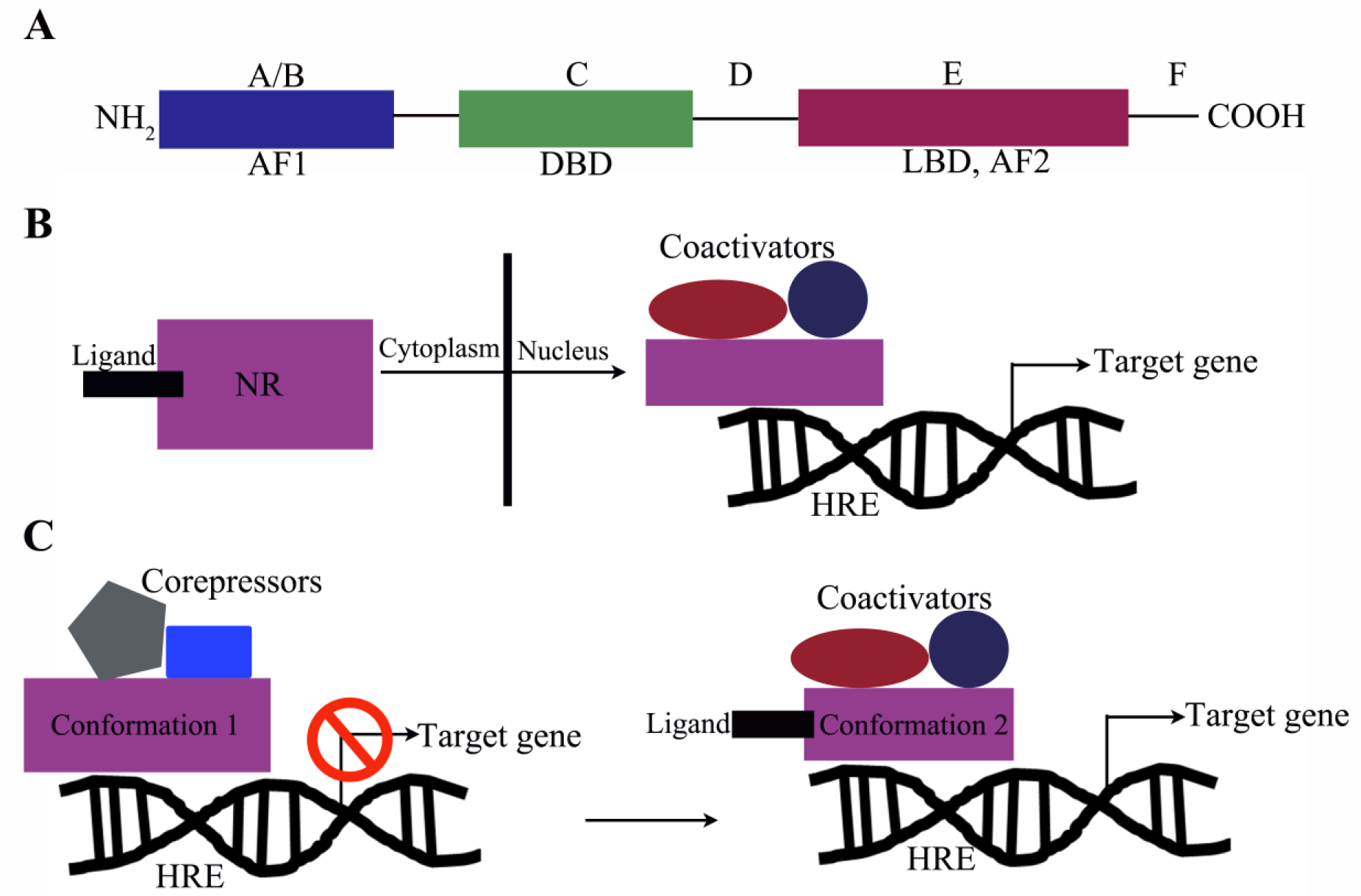
NR domain structure and mechanism of action. (
Human Nuclear Receptors.
NRs typically function as ligand-dependent transcription factors. Upon ligand binding, NRs recognize specific DNA-response elements called hormone receptor elements (HREs), located within the promoters of their cognate target genes, where the ligand-NR complex can modulate the ability of HREs to recruit a range of other transcriptional proteins that alter the rate of gene expression ( Fig. 2B , C ). Besides ligands and the sequence of HREs, an additional level of specificity is provided by the preferences of NRs for their binding partners. NRs can be organized into distinct oligomeric states, including homodimers (e.g., steroid receptors) and heterodimers with the “promiscuous” retinoid X receptor (RXR) (e.g., peroxisome proliferator-activated receptors [PPARα, β, γ; NR1C1, 2, 3], 17 thyroid hormone receptors [TRα, β; NR1A1, 2], and several orphan receptors), or remain monomeric (e.g., steroidogenic factor 1 [SF-1; NR5A1]). In the absence of a ligand, NRs are either present in the cytoplasm, where they form a complex with heat shock proteins and immunophilin chaperones, 18 or constitutively bound to the HREs in the nucleus, where they form complexes with corepressor proteins like SMRT/NCOR and HDAC, repressing transcription. 19 In the first scenario, ligand binding leads to dissociation of NRs from the chaperones and translocation to the nucleus, where they bind to the HREs of their target genes ( Fig. 2B ). Conversely, in the second scenario, ligand binding leads to dissociation of the corepressor complex and recruitment of transcription factors/enhancers, leading to activation of gene expression ( Fig. 2C ). This highlights the highly selective and variable nature of NRs.
NRs and Ocular Diseases
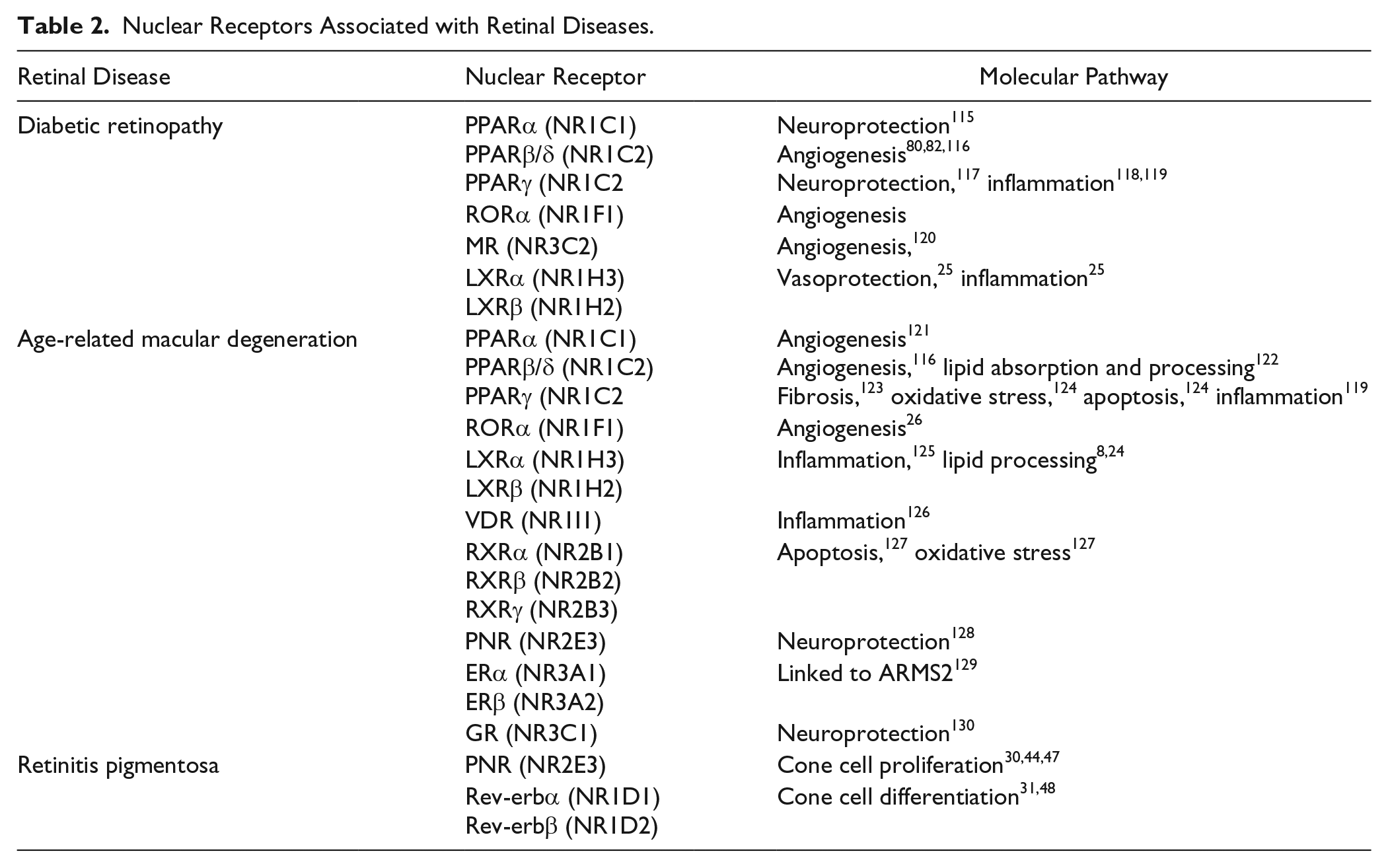
A large number of studies have revealed that members of the NR superfamily serve a pivotal role in modulating gene regulatory programs that control metabolism, inflammation, extracellular matrix regulation, angiogenesis, and numerous other biological and physiological processes.5-7,20 The fundamental roles that NRs play are highlighted by the fact that many of the biological pathways regulated by NRs are altered in human diseases, including common metabolic and cardiovascular disease states, autoimmune diseases, certain dermatologic conditions, and various tumor formations, to name a few.5,21-23 Within the last 15 years, an almost certain significant role for NRs has also emerged in studies of retinal degenerations, including AMD, DR, and RP, in part through studies conducted in genetically modified mouse models.8,11,24-26 Further identification of potential candidate receptors in diseases affecting the outer retina stems from the development of an NR atlas of human retinal pigment epithelial cells, which resulted in a comprehensive evaluation of the expression of all human NRs in these cells. 27 Similarly, candidate receptors in diseases affecting the inner retina arose from targeted evaluation of the expression of select receptors within the retina.11,28-31 Collectively, these studies have provided a great deal of information regarding the impact of dysregulated NR signaling on the molecular pathogenesis of various diseases of the eye. It is remarkable that several of the NRs identified in the retina belong to the “orphan receptor” category, without known ligands. 11 This fact in and of itself opens up the field of ophthalmology to questions concerning unidentified receptor ligands and their role in basic eye physiology and disease. Importantly, it also fuels interest in synthetic ligands that may eventually find applications as therapeutic agents to treat retinal diseases. The rest of this review will focus on the role of NRs in regulating molecular pathways, which are important in the pathogenesis of retinal diseases, including RP, DR, and AMD, and the prospects of using NRs as therapeutic targets for drug development. Table 2 summarizes human NRs potentially involved in retinal diseases.
Nuclear Receptors Associated with Retinal Diseases.
Retinitis Pigmentosa
RP is a group of heterogeneous inherited neurodegenerative retinal diseases. The prevalence of RP is approximately 1 in 4000 people, totaling close to 1.5 million affected individuals worldwide.32,33 It is characterized by progressive night blindness, gradual loss of the visual field, a decline in electroretinogram-recorded amplitudes, and compromised visual acuity. 32 The pathogenesis of RP can be explained by a continuum of metabolic maladies that lead to morphological and functional compromise to the retina, including the death of rod photoreceptors, followed by progressive loss of cone photoreceptors; loss of the retinal pigment epithelial cells, which serve as vital support cells to the overlying photoreceptors; and eventually total blindness.34-38 The primary cause of photoreceptor degeneration, although influenced by various processes, is genetic and can be mapped to defects in multiple gene loci. To date, RP has been localized to a number of chromosomes, including 1, 3, 4, 6, 7, 8, 11, 14, 16, and 19 and X chromosome.36,39-41 A recent update of all the genes involved in retinal diseases can be found through the Retinal Information Network (RetNet, https://sph.uth.edu/retnet/). In general, four classes of mutations can be described in the pathogenesis of RP on the basis of the functional molecular mechanisms that are disrupted: (1) mutations that disturb visual transduction, (2) mutations affecting vitamin A metabolism, (3) mutations that impair photoreceptor differentiation and homeostasis, and (4) mutations affecting the turnover of photoreceptor outer segments.11,42
Several research groups have shown that select NRs are actively involved in photoreceptor physiology. For example, NR2E3, also known as photoreceptor cell–specific NR (PNR), plays a role in the regulation of photoreceptor-specific genes necessary for cell maintenance and phototransduction and during development and differentiation in rod photoreceptor cells.29,43,44 With regards to RP, Coppieters et al. identified a heterozygous missense mutation (G56R) in the first zinc finger of the Nr2e3 gene in a large Belgian family with this disease. 45 Furthermore, the autosomal recessive retinal degeneration 7 (rd7) mouse, identified at the Jackson Laboratory, has been presented as a murine model for NR2E3-associated retinal disease.30,46 These rd7 mice are characterized by retinal folding associated with retinal spots and late-onset retinal degeneration 46 and exhibit phenotypes similar to those of patients with Nr2e3 mutations, namely, a significant increase in S-cones and progressive degeneration of rod and cone photoreceptor cells. 30 Significant crosstalk is often found between various NRs. 2 In fact, NR2E3 has been shown to interact with another orphan NR, NR1D1 (Rev-Erbα), in mice, and functions within the same transcriptional system.29,31,47 In support of this, shRNA-mediated knockdown of Nr1d1 in the mouse retina reportedly results in a retinal phenotype similar to that observed in the rd7 mice. 31 Furthermore, the therapeutic strategy of gene delivery of Nr1d1 to the retina, performed in neonatal Nr2e3rd7/rd7 mice, was able to rescue retinal spotting and dysplasia associated with Nr2e3 loss. Finally, Nr1d1 gene delivery was also able to reregulate the key genes that are vital for photoreceptor homeostasis, which were misregulated by the absence of Nr2e3 in Nr2e3rd7/rd7 mice. 48 This study established Nr1d1 as a potential therapeutic target that may be used as a tool for gene therapy. In addition to gene therapy, high-throughput screens for receptor ligands have identified small molecules that bind to the NR2E3. 49 One of the ligands identified, compound 11a, was further tested in retinal explants and shown to induce expression of genes with DR1 and DR4 motifs in their promoters. 50 Currently, there are no NR-based treatments for RP in clinical trials. This presents both a great challenge and an opportunity for the development of drugs or gene therapy–based therapeutic agents for RP.
Diabetic Retinopathy
DR is a common microvascular complication associated with diabetes, and is characterized by hyperglycemia, loss of pericytes, microaneurysms, and preretinal neovascularization, which can eventually lead to vision loss by hemorrhage and retinal detachment.51,52 Diabetes is now recognized as a global epidemic with a projected patient population of 380 million by 2025.53-55 As the global prevalence of diabetes increases, so will the incidence of DR. Approximately one-third of individuals with diabetes will develop some degree of retinopathy, which currently is the leading cause of blindness in patients aged 20–74 years, in the developed and developing countries. 56 Broadly defined, DR can manifest in two forms: (1) nonproliferative DR and (2) proliferative DR. Nonproliferative DR can be further classified into mild, moderate, and severe stages that may or may not involve the development of diabetic macular edema, the most common cause of diabetes-related vision loss, while proliferative DR is characterized by retinal neovascularization.57,58 The possibility of developing DR can be decreased by early detection and control of blood glucose, blood pressure, and lipid intake.52,53 Although a strong link has been established between hyperglycemia and the development and progression of DR, the causal mechanism that leads to damage to the retinal microvasculature has yet to be clearly established.59,60
Similar to the brain, the retina is compartmentalized from the circulation by the inner blood–retina barrier, which is formed by the retinal microvasculature (retinal microvascular endothelial cells and pericytes), and by the outer blood–retina barrier, which is formed by the capillary endothelium and the tight junctions of retinal pigment epithelial cells. 61 During DR, the inner blood–retina barrier is compromised, which may lead to diabetic macular edema. There is strong evidence that growth factors like vascular endothelial growth factor (VEGF), which regulates vascular permeability, play a key role in the development of edema. 62 Existing treatment modalities for macular edema include laser photocoagulation, corticosteroids, and anti-VEGF agents, and their success is most likely due to targeting inflammation as well as VEGF-A.61,63,64 However, therapies for nonproliferative DR in the absence of macular edema are still lacking.
PPARγ (NR1C3) is a DR candidate receptor, as it has been shown to play an important role in a variety of biological processes dysregulated in diabetes and DR, including adipogenesis, glucose metabolism, angiogenesis, and inflammation. 65 Thiazolidine derivatives like rosiglitazone and pioglitazone act as synthetic ligands for PPARγ, and are being used as oral antihyperglycemic agents for the therapy of non-insulin-dependent diabetes mellitus. 66 Studies investigating the role of PPARγ activation on the pathogenesis of proliferative DR have shown that treatment with rosiglitazone can inhibit retinal leakage, as well as leukostasis, in the streptozotocin-induced model of DR, in part through inhibition of NF-κB and subsequent suppression of ICAM-1 expression, which aids in leukocyte adhesion to endothelial cells.65,67-69 Additionally, activation of PPARγ leads to suppression of pro-inflammatory gene expression and cytokine secretion.70-72 Furthermore, the PPARγ agonists rosiglitazone and troglitazone have been shown to inhibit retinal neovascularization in a murine model of oxygen-induced ischemia. 73 In addition to regulating inflammation, PPARγ can negatively impact VEGF-mediated angiogenesis via modulation of cyclooxygenase 2, which is a key modulator of inflammatory angiogenesis.74-76 PPARγ agonists are already being used to treat diabetes and other metabolic disorders. These studies, along with PPARγ’s established ability to inhibit inflammation and angiogenesis, portend of their potential utility as an alternative and/or complement to laser photocoagulation and anti-VEGF therapy in the treatment of DR. 77
Another PPAR isoform, beta/delta (PPARβ/δ; NR1C2), may also be a promising target for DR. In addition to controlling oxidative metabolism at a ubiquitous level, PPARβ/δ has been shown to be a molecular modulator of inflammation, 78 mechanistically through repression of NF-κB/AP-1-based transcription of the inflammatory response. 79 A recent study reported that PPARβ/δ antagonism in a model of oxygen-induced retinopathy led to a decrease in preretinal neovascularization. 80 GSK0660, a PPARβ/δ antagonist, was also shown to significantly decrease serum-induced human retinal microvascular endothelial cell proliferation and tube formation, two commonly used in vitro–based angiogenic assays, in a dose-dependent manner.80,81 Another study from the same group showed that GSK0060 stabilized tight junctions in human retinal microvascular endothelial cells treated with VEGF, concomitant with reducing VEGFR1/2 expression, suggesting a role in vascular permeability, 82 and providing further support for pharmacologic inhibition of PPARβ/δ as the basis for therapeutic targeting of retinal neovascularization.
The retinoic acid receptor-related orphan receptor alpha (RORα; NR1F1) is a lipid-sensing NR that has been shown to regulate the inflammatory response in allergy and autoimmune diseases.83-85 A recent report by Sun et al. demonstrated that the absence of RORα inhibited pathological angiogenesis in a model of oxygen-induced retinopathy, by directly impacting the expression of suppressor of cytokine signaling 3 (SOCS3), which is a critical regulator of tissue inflammation. 26 Furthermore, treatment with an inverse agonist of RORα, SR1001, significantly repressed pathological angiogenesis in oxygen-induced retinopathy. Thus, RORα may also serve as a novel target for drug development to treat pathological retinal angiogenesis without directly altering levels of angiogenic growth factors, such as VEGF, which are essential for physiological angiogenesis.
Another NR of interest in DR is the liver x receptor (LXRα, β; NR1H2, 3). LXRs serve as cholesterol sensors and regulate cholesterol homeostasis. 86 Importantly, they also appear to be at the intersection of lipid metabolism and inflammation, as activation of LXRs has also been shown to modulate macrophage function and result in potent anti-inflammatory effects.86,87 In a recent study of DR, a link between LXR expression in endothelial progenitor cells, critical for mediating vascular repair, and micro- and macrovascular complications was observed. 25 Mice in which LXR was absent developed remarkable acellular capillaries and endothelial progenitor cell dysfunction similar to that observed in streptozotocin diabetic mice fed a high-fat diet. The most promising findings of this study were from the treatment of diabetic mice with the LXR agonist GW3965, which resulted in decreased micro- and macrovascular changes, including fewer acellular capillaries and reduced activated retinal glial cells, as demonstrated by glial fibrillary acidic protein (GFAP) immunoreactivity. Collectively, these studies point to the need to not only further consider targeting NRs such as the PPARs, ROR, and LXRs, as pharmacological targets for the treatment of DR, but also examine additional lipid- and immune-regulating NRs in the pathobiology of DR.
Age-Related Macular Degeneration
AMD is the leading cause of progressive vision loss in the elderly population, affecting 60 million people worldwide.88,89 The pathogenesis of early AMD involves accumulation of lipid and protein-filled extracellular deposits called drusen under the retinal pigment epithelial cells.90-94 Clinically, AMD progresses from an early to intermediate stage, and eventually to advanced stages, which can be classified into geographic atrophy or “late dry” and neovascular or “wet” AMD. The late dry form is characterized by the death of retinal pigment epithelial cells and subsequent loss of overlying photoreceptors in the central macula and vision loss.92,95-97 Wet AMD affects approximately 10% of the patients suffering from the disease, and is characterized by a breach through the retinal pigment epithelial cell layer and choroidal neovascularization within the outer retinal space, ultimately leading to scarring in the macular region.96,98 At present, there are no effective treatments available for the late dry clinical subtype, and success with anti-VEGF therapy of wet AMD has been seen in only a subset of patients. 99 Therefore, there is a critical unmet need to identify new targets and develop alternate therapeutic approaches to help people suffering from vision loss due to AMD. The challenge with finding promising treatments for AMD is in part a reflection of the complexity of the disease, in which genetic, environmental, and systemic factors all play a role.8,96 The discovery of a large number of risk factors, in turn, has resulted in the identification of multiple molecular pathways that are believed to be dysregulated in AMD pathogenesis, including lipid metabolism, inflammation, extracellular matrix turnover, angiogenesis, complement dysregulation, and mitochondrial dysfunction, to name a few. 96 Noteworthy is that interest in studying NRs in AMD stems from the fact that the NR superfamily members are known to regulate many of these AMD-associated pathways in other diseases, such as cancer, atherosclerosis, and autoimmune disease.5,6,8,22,24,96 The complexity of the disease can also be attributed to the tight interplay of cells, including the retinal pigment epithelial cells, photoreceptors, choroidal endothelial cells, and immune cells such as macrophages and microglia, the dysfunction of each of which contributes to AMD progression. Retinal pigment epithelial cell function and its role in drusen formation, and the secretion of angiogenic factors, are extensively studied, as the overall health of these cells is affected in all clinical subtypes of AMD. 96 Because of this, recently, an NR atlas investigating the expression of all 48 human NRs in retinal pigment epithelial cells was developed, which led to the identification of several AMD candidate genes currently being studied. 27
As mentioned earlier, LXRs are vital regulators of cholesterol homeostasis, glucose homeostasis, detoxification of bile acids, immunity, and neurological functions. 100 A recent study elegantly demonstrated promising results in targeting the LXR receptor for the treatment of neovascular AMD. The investigators found that the expression of the cholesterol transporter, ABCA1, also an LXR target gene, decreases as a function of age in both human and mouse macrophages, 101 and in the absence of the ABCA1 receptor, mice develop larger and more severe experimentally induced neovascular lesions. 101 Of significance, it was purported that the cholesterol content of macrophages specifically impacts macrophage phenotype and polarity, which consequently effects development of choroidal neovascularization. Though the impact of targeting LXR in dry AMD was not studied, the results support the hypothesis that targeting LXR may be effective in treating or removal of lipid-rich drusen, characteristic of early AMD. Overall, this study, in combination with other reports demonstrating activation of LXR, can attenuate inflammatory responses and support further pursuit of targeting LXRs for drug development in the treatment of choroidal neovascularization, as well as inflammatory mechanisms associated with AMD.102-105
Different isoforms of PPARs also have the potential to be viable targets for AMD, as they are involved in the regulation of established AMD pathogenic pathways, such as lipid metabolism, extracellular matrix remodeling, angiogenesis, and inflammation.8,24 Direct evidence comes from one study that examined the distribution of PPARγ in ocular samples from human dry and wet AMD patients and found increased immunoreactivity within the retina. 106 Changes in the expression levels of PPARγ in retinal pigment epithelial cells, as a function of age and disease, were not reported. Interestingly, in cell cultures of ARPE19 cells, a spontaneously arising retinal pigment epithelial cell line from a 19-year-old donor, following treatment with hydrogen peroxide, there was an increase in PPARγ along with VEGF, heme-oxidase 1, and MMP-9 expression. Evidence is also available in support of potentially targeting PPARγ in wet AMD, based on in vivo treatment of rats following experimentally induced choroidal neovascularization. 107 Conceding that there is an absence of reports that directly link the PPARα or PPARβ/δ isoforms with human AMD pathology, functional studies in ARPE19 and human primary retinal pigment epithelial cells have conjectured that these PPARs play a role in AMD pathology by regulating inflammation, oxidative stress, and lipid accumulation. 8 While there are no PPAR Food and Drug Administration (FDA)–approved drugs for AMD, there are FDA-approved drugs for PPARγ and α. Additionally, recent development of highly specific and selective compounds for PPARβ/δ, such as GW501516, GW0742X, and GSK3787, has improved the study of the PPARβ/δ pathway in disease models such as obesity and atherosclerosis, which may provide improved insight into the regulation of inflammation and lipid metabolism through this pathway. Future studies utilizing these PPAR-targeted drugs in in vitro and in vivo models of AMD have the potential to lead to the development of new therapeutic approaches treating pathways associated with AMD progression. 108
The aryl hydrocarbon receptor (AhR) is a member of the bHLH/PAS (basic helix–loop–helix/Per–Arnt–Sim) family of heterodimeric transcriptional regulators, with similar mechanisms of action to NRs. Although traditionally thought of as a receptor involved in toxin clearance, it has also been shown to play important roles in regulation of vascular development, angiogenesis, and inflammation.109-112 Several recent studies have reported a potential role for AhR in AMD.112-114 These studies found that genetic ablation of AhR in mice leads to development of ocular features of dry AMD. One study further reported an age-related decline of AhR activity in human retinal pigment epithelial cells, suggesting a potential age-related compromise in normal retinal epithelial cellular clearance mechanisms. 113 Interestingly, the AhR knockout mice also displayed progressive choroidal thinning. A follow-up study identified a role of AhR in neovascularization. In aged mice, the absence of AhR resulted in an exacerbation of experimentally induced choroidal neovascularization concomitant with increased extracellular matrix dysregulation and inflammation. 112 These studies solicit exploring the therapeutic benefits of targeting the AhR signaling pathway as a means for treating deposition of extracellular matrix material, angiogenesis, and fibrosis associated with AMD.
Discussion
Though not exhaustive, we have provided an overview of the potential role of NRs in the pathobiology of several vision-impairing retinal diseases. This is a relatively young field of research with few studies to date. However, the abundance of NRs in the eye strongly support the concept of the eye as a specialized secondary endocrine organ, in which NRs most likely orchestrate complex events during development, cell differentiation, cell homeostasis, and by extension, disease. Furthermore, the overwhelming number of studies characterizing functions of various NRs in systemic and degenerative diseases that share common pathogenic pathways with retinal disorders underscore the need to examine the role of NRs in the initiation and progression of complex ocular diseases. Finally, it will be critical to fully tease out cell-specific activity of NRs, including their responses following treatment with classically defined modulators, including activators and inhibitors. A thorough evaluation of receptor activity may lead to identification of cell-specific responses reflecting the complex pharmacology of NRs. This will be critical in determining the utility of targeting NRs for treatment in the eye with selective modulators that have the potential to decrease detrimental off-target effects.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Goldis Malek is the recipient of funding from the National Eye Institute (NEI EY02868), the Brightfocus Foundation for Macular Degeneration Research, and the Edward N. & Della L. Thome Memorial Foundation AMD Research Award. Additional funds come from an unrestricted Research to Prevent Blindness award and NEI core grant (NEI P30 EY005722) to the Department of Ophthalmology at Duke University.