
Abstract
Recent progress of genetic studies has dramatically unveiled pathogenesis of acute myeloid leukemia (AML). However, overall survival of AML still remains unsatisfactory, and development of novel therapeutics is required. CCAAT/enhancer binding protein α (C/EBPα) is one of the crucial transcription factors that induce granulocytic differentiation, and its activity is perturbed in human myeloid leukemias. As its reexpression can induce differentiation and subsequent apoptosis of leukemic cells in vitro, we hypothesized that chemical compounds that restore C/EBPα expression and/or activity would lead to myeloid differentiation of leukemic cells. Using a cell-based high-throughput screening, we identified 2-[(E)-2-(3,4-dihydroxyphenyl)vinyl]-3-(2-methoxyphenyl)-4(3H)-quinazolinone as a potent inducer of C/EBPα and myeloid differentiation. Leukemia cell lines and primary blast cells isolated from human patients with AML treated with ICCB280 demonstrated evidence of morphological and functional differentiation, as well as massive apoptosis. We performed conformational analyses of the high-throughput screening hit compounds to postulate the spatial requirements for high potency. Our results warrant a development of novel differentiation therapies and significantly affect care of patients with AML with unfavorable prognosis in the near future.
Introduction
Recent advances in genetic studies have led to a dramatic progress in understanding the pathogenesis of acute myeloid leukemia (AML). However, long-term survival of AML still remains unsatisfactory. In particular, AMLs with monosomal or complex karyotypes have shown the worst prognosis, with a 3-year rate of overall survival being only 12%. 1 Moreover, AMLs arising from myelodysplastic syndrome (MDS) or secondary to previous cytotoxic chemotherapy have a lower rate of remission than de novo AMLs. Patients with high-risk MDS have a 3-year survival rate of less than 10%. 2 Considering the fact that conventional chemotherapies do not have specific targets and demonstrate various levels of toxicity, there is a great demand for novel targeted therapeutics. One of the best approaches is to focus on a block of differentiation, which is a hallmark of all subtypes of AML. All-trans retinoic acid (ATRA) is now widely used as the first-line therapy for one subtype of AML, t(15;17) positive acute promyelocytic leukemia (APL), and induces differentiation of leukemia cells and eventually leads to apoptosis. Although it has been shown that ATRA can induce remission and lead to cure in nearly 70% of patients with APL, 3 its application in other types of myeloid leukemia is limited. In addition, relapse can occur in the course of treatment. Although arsenic trioxide has a high rate (85%) of successful remission induction in patients with APL resistant to ATRA, an 18-month relapse-free survival is 60%. 4
Expression of CCAAT/enhancer binding protein α (C/EBPα) is increased and maintained during granulocytic differentiation and rapidly downregulated during the alternative monocytic pathway. 5 Conditional expression of C/EBPα in stably transfected myeloid precursor cells triggers neutrophilic differentiation, concomitant with upregulation of the granulocyte colony-stimulating factor receptor (G-CSFR) and secondary granule proteins. 5 In mice deficient in C/EBPα, there is a block in granulocytic differentiation at the myeloblast stage, while all the other blood cell types are present and intact. 6 Thus, C/EBPα is necessary and sufficient for neutrophil differentiation. Consistent with its importance in normal myeloid differentiation, expression and/or function of C/EBPα are perturbed in various types of myeloid leukemias by different mechanisms (transcriptional silencing, translational inhibition, posttranslational modification, decrease in DNA binding, or point mutations resulting in increased production of a dominant negative form). 7 Thus, restoration of C/EBPα expression and/or activity could overcome the block of differentiation and lead to growth arrest and apoptosis of leukemic cells.
In the current study, we established a stable cell line carrying luciferase gene driven by an artificial promoter composed of a tetramer of C/EBP binding sites, which responds to C/EBPα activity. By using this indicator line in a cell-based high-throughput screen, we identified one chemical compound, 2-[(E)-2-(3,4-dihydroxyphenyl)vinyl]-3-(2-methox-yphenyl)-4(3H)-quinazolinone (referred to as ICCB280), which induced myeloid differentiation of leukemia cell lines by increasing expression of C/EBPα and C/EBPε and decreasing expression of c-Myc. Importantly, exposure of primary blast cells from patients with AML to ICCB280 led to differentiation and apoptosis as well. Interestingly, G-CSFR, which is one of targets of C/EBPα, was upregulated by the treatment with ICCB280, and the addition of G-CSF further enhanced the differentiation induced by this compound. These results indicate that the use of ICCB280 alone or in combination with other agents, such as G-CSF, may provide a novel means of treatment of AML. Given the fact that in addition to AML, abnormalities in C/EBPα expression and/or function have been also reported in other malignancies such as lung 8 and liver, 9 we believe that small-molecule agents that selectively target C/EBPα, such as ICB280, may find broad applications in treating cancers.
Materials and Methods
Reagents
ATRA (cat. R2625) was purchased from Sigma (St. Louis, MO). 2-[(E)-2-(3,4-dihydroxyphenyl)vinyl]-3-(2-methoxy-phenyl)-4(3H)-quinazolinone (ICCB280) was purchased from ChemBridge (San Diego, CA). Stock solutions for the compounds were prepared in DMSO and stored at −20 °C. The drugs were diluted in fresh medium before each experiment, and the final DMSO concentration was <0.5%.
Determination of the Purity and Molecular Weight of ICCB280
High-performance liquid chromatography–mass spectrometry (HPLC-MS) analysis was performed with a Waters Alliance-Micromass ZQ instrument, an ESI source, and Waters 2489 UV/visible detector at 254 nm (Milford, MA). Analysis was performed using an analytical Waters Symmetry C18 column (3.5 mm, 2.1 × 100 mm) operating at 0.5 mL/min with a linear gradient of 0% to 100% of acetonitrile in water (containing 0.1% formic acid, v/v) over 10 min.
Cells
HL-60 (American Type Culture Collection [ATCC], Manassas, VA; CCL-240), K562 (ATCC; CCL-243), and U937 (ATCC; CRL-1593.2) cells were maintained in RPMI 1640 with 10% fetal bovine serum at 37 °C with 5% CO2. CV-1 (ATCC; CCL-70) and 293T (ATCC; CRL-3216) were maintained in Dulbecco’s modified Eagle’s medium (DMEM) with 10% fetal bovine serum. Primary AML patient blasts were collected from peripheral blood after obtaining patients’ informed consents, which were approved by the institutional review boards at Beth Israel Deaconess Medical Center. Patient 0502 was diagnosed as having acute myelomonocytic leukemia with abnormal eosinophils (AML M4Eo) and carried inv(16)(p13;q22). Patient 0505 was diagnosed as having acute monocytic leukemia (M5) with normal karyotype. Mutations in the CEBPA gene were reported in neither patient. Primary AML blast cells were isolated using Ficoll-Paque Plus (Amersham Biosciences, Piscataway, NJ), as previously described, 10 and maintained in culture in RPMI 1640 with 10% fetal bovine serum in the presence of G-CSF (60 ng/mL) at 37 °C with 5% CO2.
Plasmid Constructs
Firefly luciferase gene controlled by a minimal thymidine kinase (TK) promoter and a tetramer of C/EBP-binding sites from the human G-CSFR promoter (4xCEBP-luc) was previously described.
11
To make 4x mutCEBP-luc, oligonucleotides containing mutations abolishing C/EBP binding (AAGGTGTT
Generation of the C/EBP Activity Indicator Cell Line
U937 cells were cotransfected with the ScaI-linearized 4xCEBP-luc construct together with the linearized plasmid containing neomycin-resistant gene (pSV40-neo) by electroporation using 250 V and 960 µF in Gene Pulser II (BioRad, Hercules, CA), followed by selection in 1 mg/mL G418. Single clones were isolated by limiting dilution in 96-well plates.
Generation of HL-60 Cells Stably Expressing shRNAs against CEBPA
293T cells were cotransfected with C/EBPα shRNA in pGhU6 vector or the shRNA control and lentiviral constructs Gag-Pol and Env. HL-60 cells were then infected with virus that was harvested and concentrated using a Centricon Plus-70 100000 MWCO column (Millipore, Billerica, MA). Infected cells were detected by enhanced green fluorescent protein flow cytometry analysis.
High-Throughput Screening of Chemical Libraries
Stable U937-C/EBP clones were maintained in the RPMI 1640 phenol red–free medium, 10% fetal bovine serum (FBS), 100 U/mL penicillin G, 100 µg/mL streptomycin, 0.25 µg/mL amphotericin B, and 1 mg/mL G418 for selective propagation. Then, 30 µL per well (2400 cells) of U937-C/EBP cells was plated in 384-well, flat-bottom, white polystyrene plates (Nalgene, Rochester, NY). All plates were prepared in duplicates. Assay plates were then incubated in 5% CO2 and at 37 °C for 24 h. Next, 100 nL of each compound dissolved in DMSO was added by a pin-transfer robot to give concentrations of 10 to 30 µM. ATRA and DMSO were added in every plate as positive and negative controls, respectively. Plates were incubated in 5% CO2 and at 37 °C for another 24 h, followed by the addition of 30 µL Bright Glo (Promega, Madison, WI) to each well. Plates were incubated at room temperature for at least 3 min, and the luciferase activity was read in sequential order using the LJL Analyst Reader (Molecular Devices, Sunnyvale, CA) in luminescence mode. The plates we screened are as follows: Institute of Chemistry and Cell Biology (ICCB) Bioactives 1, National Institute of Neurological Disorders and Stroke (NINDS) Custom Collection, SpecPlus Collection, ChemBridge Microformat, Commercial Diversity Set 1, Philippines Plant Extracts 1&2, Starr Foundation Extracts 1, ICCB Discretes 3&4, and Diversity-Oriented Synthesis Collection (http://iccb.med.harvard.edu/retired-compound-libraries/). All equipment and robotic instrumentation used for screening were provided by the ICCB at Harvard Medical School (http://iccb.med.harvard.edu/).
Nitroblue Tetrazolium Reduction Assay
Reduction of nitroblue tetrazolium (NBT) by respiratory burst products was performed as previously described. 5
Growth Inhibition Assay
Cells were plated at 10,000 cells/well with specified concentrations of compounds and incubated for 48 h. Growth inhibition was assessed by the MTS assay using CellTiter 96 AQueous One solution proliferation kit (Promega). IC50 was calculated using Prism software (GraphPad Software, La Jolla, CA).
RNA Isolation, Northern Blot Analysis, and Quantitative Real-Time PCR Analysis
Total RNA was isolated by the RNAeasy kit (Qiagen, Valencia, CA) according to the manufacturer’s instruction. In each lane, 20 µg RNA denatured in formamide was fractionated on 1% agarose-2.2 M formaldehyde gels. RNA was transferred to nylon membranes and probed as described previously. 5 Membranes were hybridized with probes for human G-CSFR 5 and c-Myc. 14 To ensure uniform levels and integrity of RNA samples loaded in each lane, the blot was stripped and rehybridized to probes specific for GAPDH. 15 Quantitative real-time (RT)–PCR analysis was performed as previously described. 13
Protein Sample Preparation and Western Blotting
Whole-cell lysates were prepared in cell lysis buffer (20 mM Tris-HCl [pH 7.5], 150 mM NaCl, 1 mM EDTA, 1 mM EGTA, 1% Triton, 1 mM β-glycerophosphate, 1 mM Na3VO4, 1 mM NaF, protease inhibitor cocktail set III [Millipore], and 1 mM phenylmethanesulfonyl fluoride [PMSF]). Lysates were cleared by centrifugation (14,000 g for 5 min in a precooled centrifuge) and boiled with Laemmli sample buffer for 3 min. Protein lysates (10–30 µg) were loaded on sodium dodecyl sulfate (SDS)/polyacrylamide gels and blotted onto polyvinylidene fluoride (PVDF) membranes (Millipore). Antibodies against C/EBPα (sc-61), C/EBPβ (sc-150), C/EBPε (sc-158), Sp-1 (sc-59), and c-Myc (sc-40) were purchased from Santa Cruz Biotechnology (Santa Cruz, CA). β-Actin antibody was from Sigma-Aldrich (A5316). All antibodies were used at 1:1000 dilution.
Apoptosis Analysis
Apoptosis was assessed using an Annexin-V-FLUOS staining kit (Roche, Penzberg, Germany) according to the manufacturer’s instructions.
Conformational Analysis
Energy-minimized conformers for the three isomeric molecules (ICCB280 and its meta- or para-methoxy isomer) were generated in silico. Structures were first drawn using Marvinsketch (Marvin 6.2.2, 2014; ChemAxon, Budapest, Hungary) and then converted into protein data bank format using the Open Babel molecular toolkit. 16 To reduce the necessary processing power, an initial steepest descent geometry optimization was performed using an MMFF94 force field (250 steps). Following the initial optimization, a weighted rotor conformer search was performed (200 iterations), followed by a conjugate gradient geometry optimization (250 steps) to generate the final 3D conformational structure of each isomer. Each structure was visualized and angles were calculated using the PyMOL molecular viewing program (Version 1.5.0.4; Schrödinger, LLC, New York, NY).
Statistical Analysis
Differences between the experimental groups were compared using the Student t test, and p values less than 0.05 were considered statistically significant.
Results
Establishment of the Cell-Based High-Throughput Drug Screen for Inducers of C/EBPα Activity
To detect C/EBPα activity, we designed a luciferase construct (4xC/EBP-luc), in which a firefly luciferase gene was placed under control of a tetramer of C/EBP binding sites from the human G-CSFR gene promoter upstream from the minimal TK promoter ( Fig. 1A ). The response of this reporter was tested in transient cotransfection assays in CV-1 cells. As shown in Figure 1B , addition of C/EBPα expression vector increased the luciferase activity 10-fold. We then transfected U937 human AML cells with the 4xC/EBP-luc construct to generate a stable cell line (U937-C/EBP). Multiple individual clones were isolated and subcloned at limiting dilution. It has been published before that treatment of U937 cells with ATRA increases levels and DNA-binding activity of C/EBPα.5,17,18 Therefore, several stable clones were tested for the response of luciferase activity to treatment with ATRA ( Fig. 1C ), and clone 10 was chosen for the high-throughput screening (HTS). Of note is that clones that harbored the mutated C/EBP binding sites showed no increase in luciferase activity upon ATRA exposure ( Fig. 1C ), indicating that an increase in luciferase activity in ATRA-treated U937-C/EBP cells is specific for C/EBP proteins.
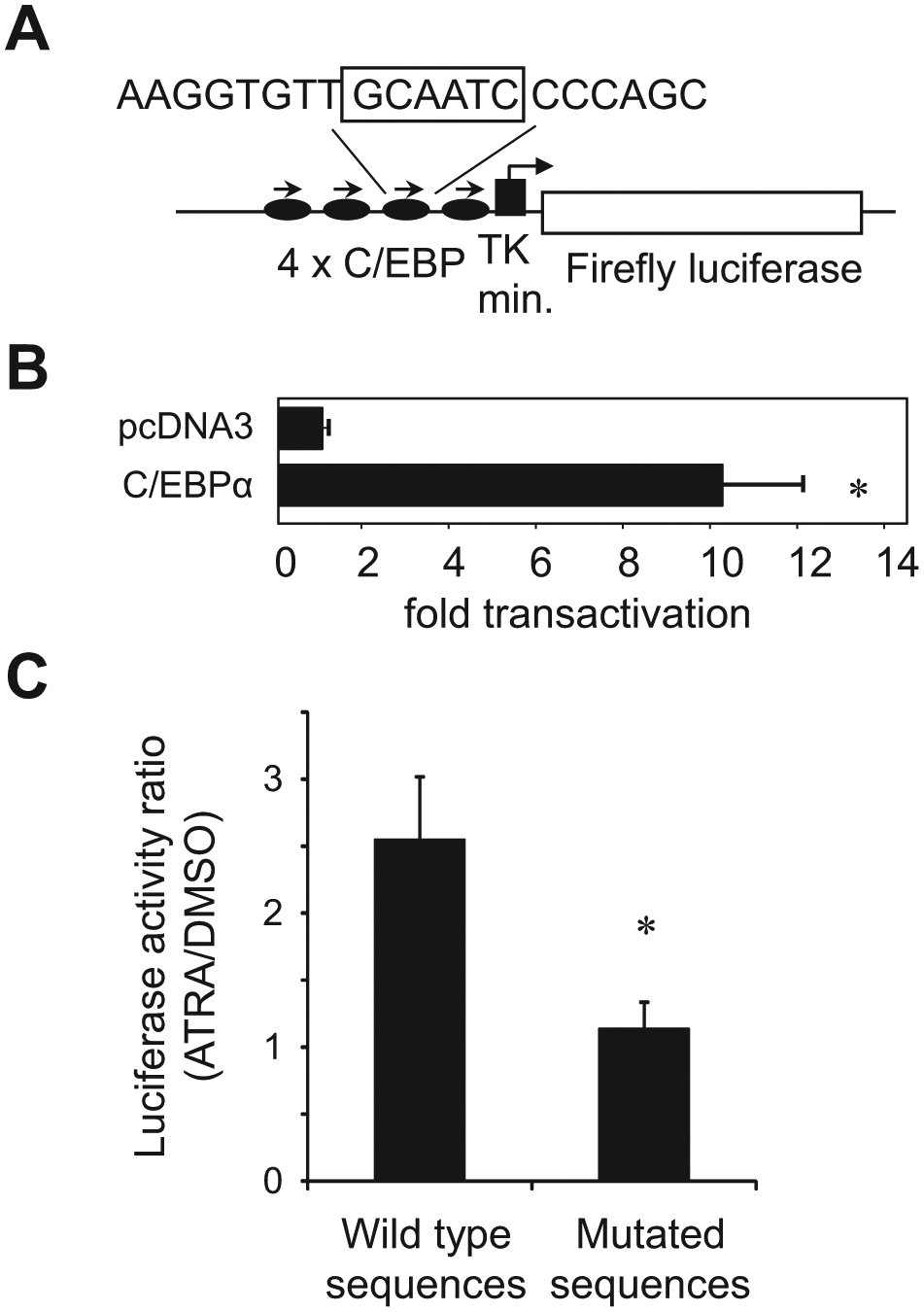
Establishment of the cell line which detect CCAAT/enhancer binding protein α (C/EBPα) activity. (
A detailed timeline of the high-throughput screen is shown in
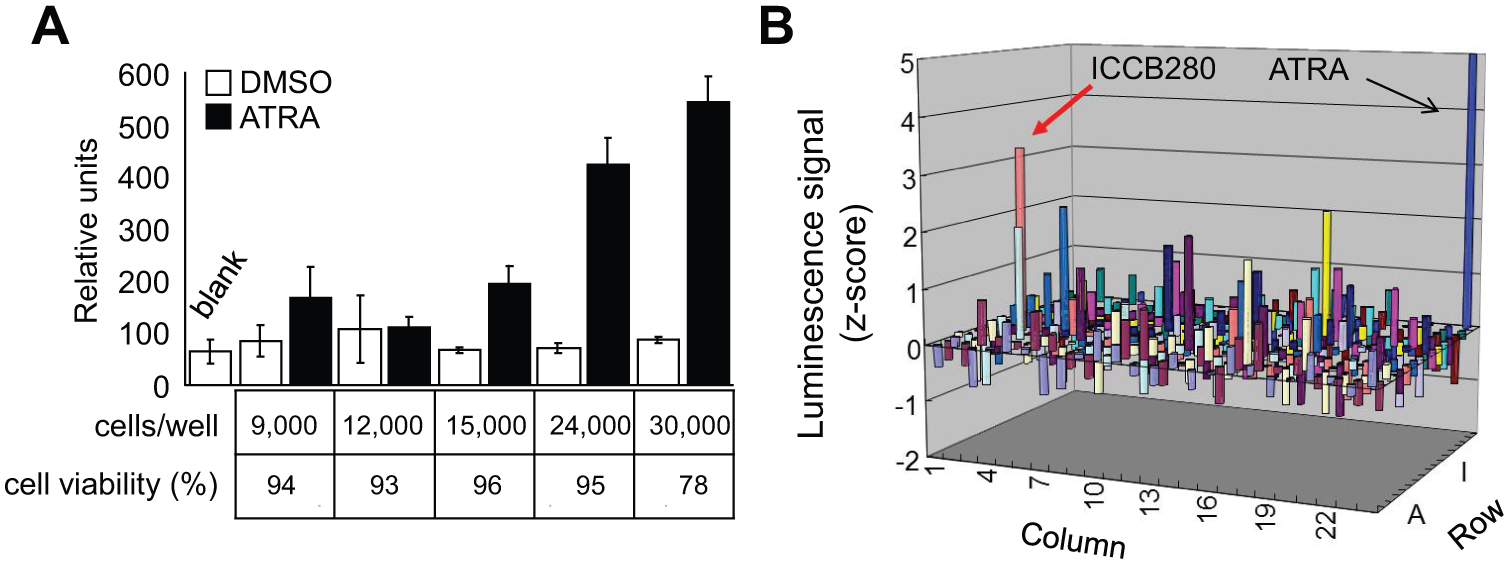
High-throughput screening of chemical compounds that increase CCAAT/enhancer binding protein (C/EBP) expression and/or activity. (
ICCB280 Induces Granulocytic Differentiation, Growth Arrest, and Subsequent Apoptosis of Leukemia Cells
Out of the 320 initial hits, we proceeded to a second screen to identify compounds that have the ability to induce granulocytic differentiation. We used the HL-60 leukemic cell line as it has been extensively used as a model for myeloid differentiation for more than 30 years. 19 We treated HL-60 cells with individual compounds at 10 µM each and examined Wright-Giemsa-stained cytospins for morphological changes. We also performed an NBT reduction assay detecting production of superoxide anion, which indicates functional maturation of leukemic cells to granulocytes. The strongest differentiation-promoting activity was demonstrated for ICCB280 ( Fig. 3A ). Seven-day treatment with ICCB280 induced granulocyte-like morphological changes such as a decrease in the nucleus-to-cytoplasm (N/C) ratio and nuclear lobulation in 83% of the cells, whereas no cells showed granulocyte-like morphology in cells treated with DMSO ( Fig. 3B , left panels). Approximately 46% of the cells were also positive in the NBT assay, compared to 1% of DMSO-treated control cells ( Fig. 3B , lower right panel, black arrows). In addition, ICCB280 induced an increase in surface CD11b expression, which is one of the characteristics of neutrophilic differentiation ( Fig. 5B ). Furthermore, treatment of HL-60 cells with ICCB280 suppressed the cell growth in 48 h, as assessed by the MTS assay ( Fig. 3C ).
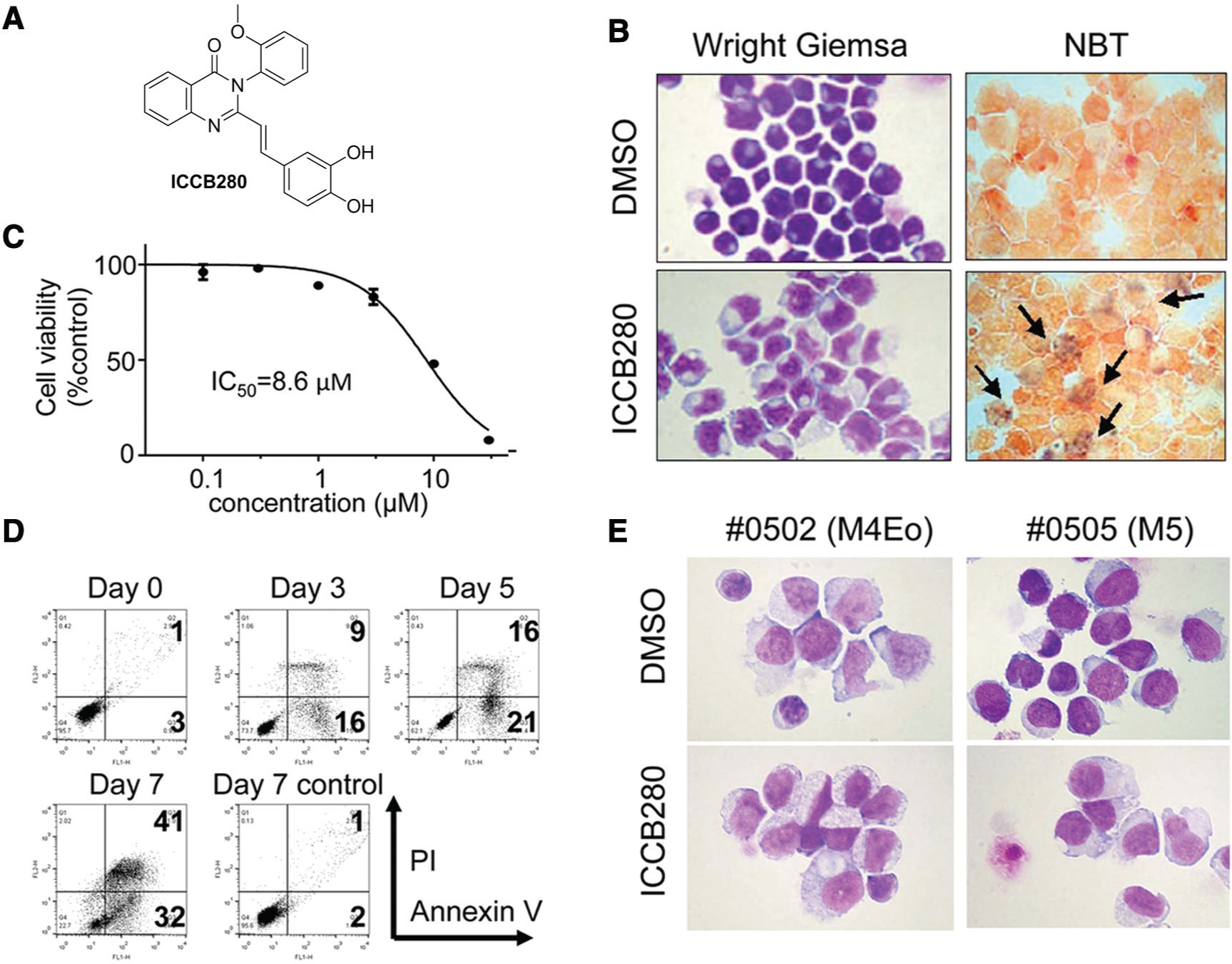
Identification of ICCB280. (
Once differentiated, neutrophils have a half-life of only 6 to 10 h in the circulation and are eventually cleared by constitutive apoptosis. 10 Therefore, induction of the differentiation-induced apoptosis would be a key to eliminate leukemic cells in differentiation therapies. To examine whether ICCB280 induces apoptosis, HL-60 cells were treated with ICCB280 and apoptosis was assessed by the Annexin V assay. An increase in early apoptotic cells (Annexin V+/PI–) was followed by that of late apoptotic cells (AnnexinV+/PI+), indicating that cell death was indeed caused by apoptosis ( Fig. 3D ). Furthermore, 41% of cells treated with ICCB280 for 7 days underwent apoptotic cell death (late apoptotic cells) ( Fig. 3D ). These results indicate that differentiated cells induced by ICCB280 eventually undergo apoptosis.
To confirm that ICCB280 has therapeutic potential, we treated two samples of primary human acute leukemia cells with ICCB280 in vitro. Leukemic blasts were isolated from either bone marrow or peripheral blood from patients with acute myeloid leukemia subtype M4 or M5, in which ATRA is not effective. After a 5-day treatment with ICCB280, leukemic cells showed a decrease in N/C (
Fig. 3E
) in 90% the cells. Moreover, the Annexin V assay showed that ICCB280 induced more cell death than the DMSO vehicle control (
ICCB280 Induces Upregulation of C/EBPα and C/EBPε but Not C/EBPβ
Since the high-throughput screen was specifically designed to induce C/EBPα expression/activity, we sought to determine whether the differentiation-promoting effect of ICCB280 leukemic cells was indeed mediated by C/EBP proteins. All C/EBP family members have the capacity to bind to the G-CSFR promoter-derived C/EBP site. Therefore, in addition to C/EBPα, we also examined two other C/EBP family members, C/EBPβ and C/EBPε. It has been show that in the absence of C/EBPα, C/EBPβ can induce emergency granulocytic differentiation by cytokine stimulation. 20 C/EBPε, on the other hand, is induced by C/EBPα 21 and is required for terminal neutrophilic maturation. 22 When HL-60 cells were treated with ICCB280, a time-dependent increase in CEBPA messenger RNA (mRNA) was detected by quantitative RT-PCR ( Fig. 4A ). Consistent with these data, upregulation of C/EBPα protein was observed as early as on day 4, followed by an increase in C/EBPε on day 6 ( Fig. 4B ). In contrast, no changes in C/EBPβ expression were noted through the 8 days of the experiment compared to control cells ( Fig. 4B ). These results demonstrate that ICCB280 upregulates C/EBPα at the transcriptional level.
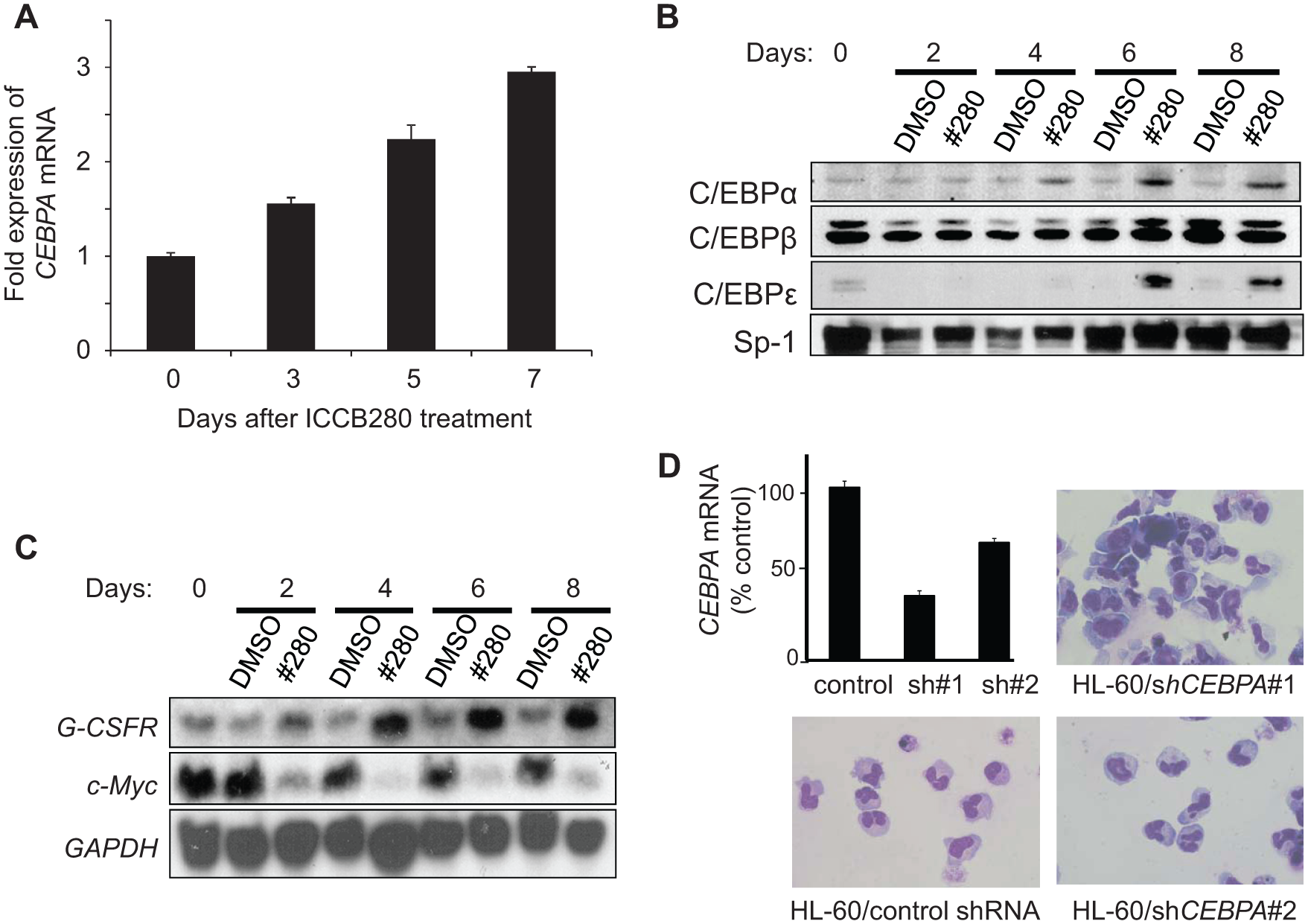
ICCB280 leads to an increase in CCAAT/enhancer binding protein α (C/EBPα) expression and modulates its target genes. (
ICCB280 Modulates Expression of C/EBPα Target Genes
Next, we examined whether ICCB280 alters expression of downstream targets of C/EBPα. Northern blot analysis demonstrated that mRNA expression of G-CSFR (CSF3R), which is a known target of C/EBPα, is rapidly upregulated in cells treated with ICCB280 (
Fig. 4C
). In contrast, both c-Myc mRNA and its protein were strongly downregulated by ICCB280 (
Fig. 4C
and
ICCB280 and G-CSF Cooperatively Induce Granulocytic Differentiation
Upregulation of CSF3R by ICCB280 prompted us to hypothesize that cells treated with ICCB280 may be more susceptible to G-CSF and that a combination of ICCB280 and G-CSF would enhance neutrophilic differentiation. As shown in Figure 5A , in the absence of ICCB280, HL60 cells did not respond to the treatment with G-CSF. However, when treated with ICCB280 together with G-CSF for 7 days, HL-60 cells showed morphology consistent with neutrophilic differentiation, and the N/C ratio was even lower than in cells treated with ICCB280 alone ( Fig. 5A ). In addition, flow cytometric analysis detected more surface CD11b-positive cells treated with the ICCB280/G-CSF combination (47%) than G-CSF (6%) or ICCB280 (25%) alone ( Fig. 5B ). These results suggest that ICCB280 induces upregulation of G-CSFR, which renders leukemic cells more susceptible to G-CSF stimulation.
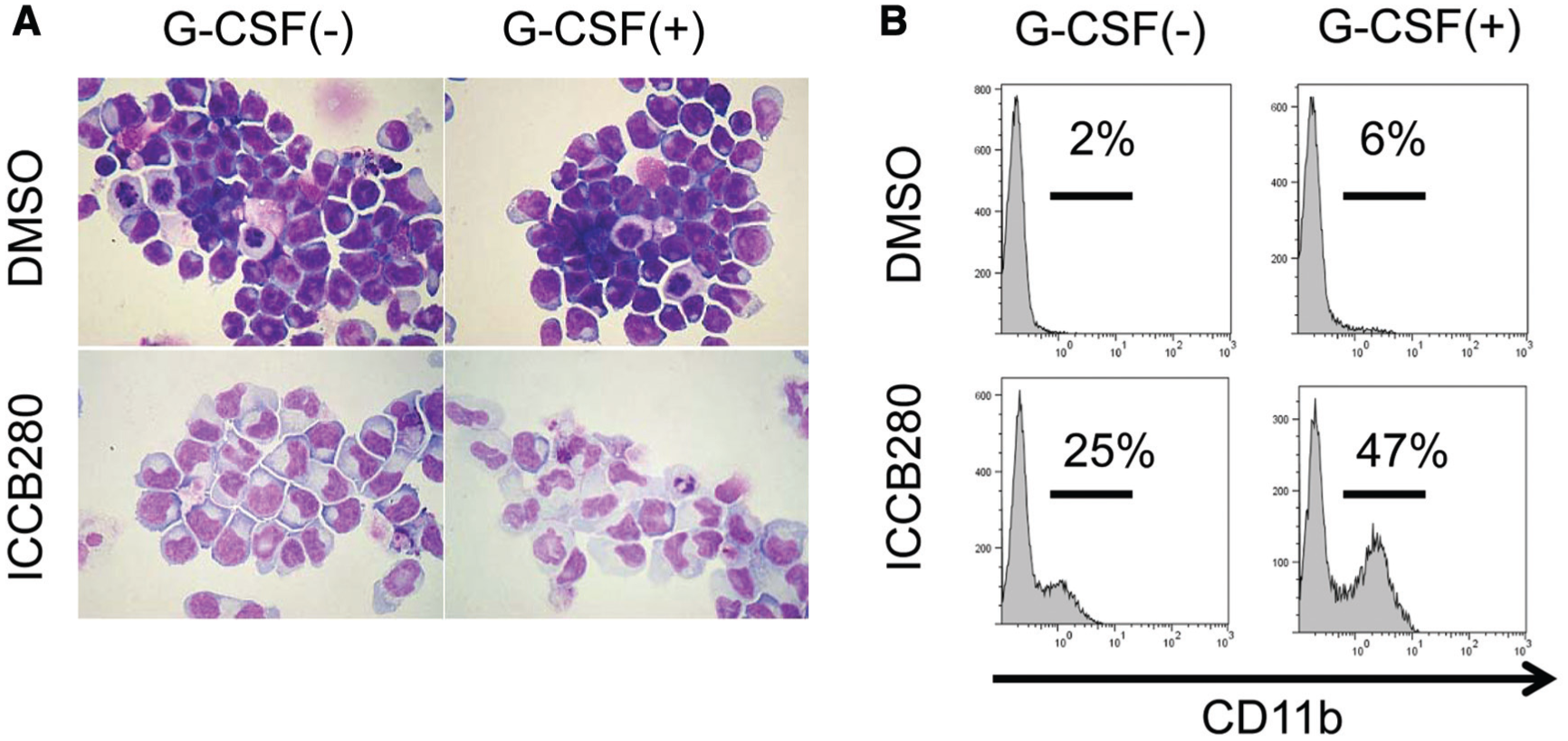
Granulocyte colony-stimulating factor (G-CSF) enhances ICCB280-induced myeloid differentiation. HL-60 cells were treated with either 10 µM ICCB280 or 0.03% DMSO in the presence or absence of 6 µg/mL G-CSF for 7 days. (
ICCB280 Adopts a Unique 3D Conformation That Affects Its Potency
To identify key structural attributes that are critical to the activity of ICCB280, we searched for its analogues. We identified 61 2-(2-arylvinyl)-3-aryl-4(3H)-quinazolinone analogues available at the Institute of Chemistry and Cell Biology (Harvard Medical School), and these compounds were screened at 10 µM each for induction of morphological changes and NBT positivity (
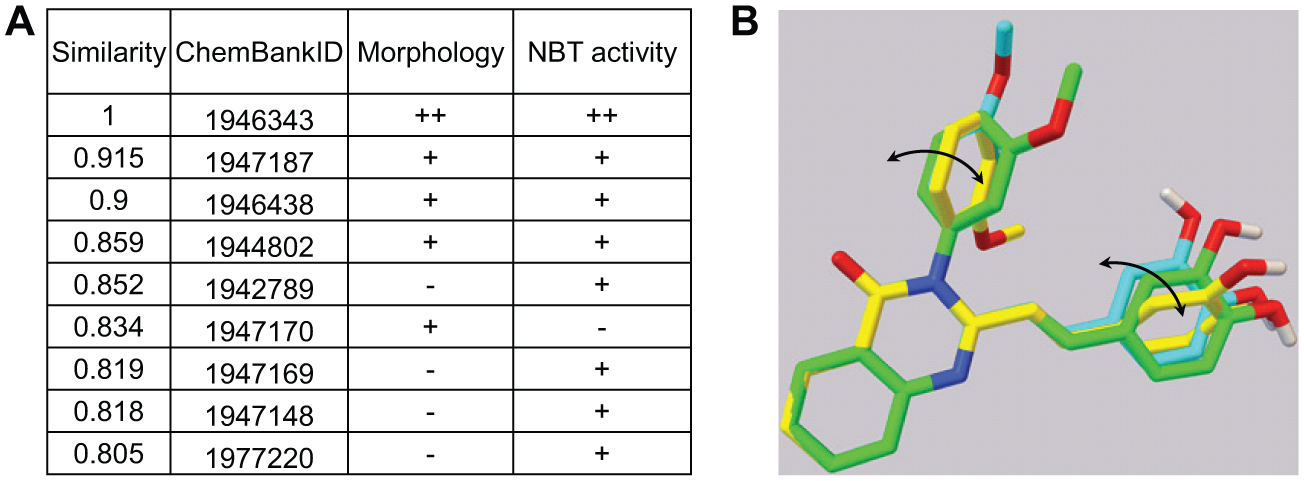
Conformational analyses of ICCB280. (
Discussion
Introduction of ATRA as the first-line therapy for t(15;17)-positive acute promyelocytic leukemia has dramatically improved prognosis, with a cure rate being nearly 70%. 23 However, despite tremendous efforts made even prior to the great success of ATRA, a differentiation therapy for other types of AML has been unavailable so far. Among the promising targets for differentiation therapy were two nuclear receptors, vitamin D receptor (VDR) and peroxisome proliferator-activated receptor gamma (PPARγ), which were described to play crucial roles in the induction of differentiation of leukemic cells. 24 However, neither vitamin D compounds nor PPARγ agonists have shown clinical impacts. 24 Thus, identification of a new strategy for differentiation therapy is still warranted.
Given that C/EBPα transcription factor plays a central role in a myeloid differentiation program, here we describe a successful establishment of a cell-based HTS to identify novel chemical compounds that enhance C/EBPα expression and/or activity and thus induce granulocytic differentiation. We identified ICCB280 as a lead compound, which led to myeloid differentiation of leukemia cells morphologically and functionally, accompanied by increased expression/activity of C/EBPα and its downstream target genes. Importantly, we showed that ICCB280 indeed induced differentiation-like morphological changes and apoptosis in primary human acute leukemia cells from two patients with AML in this proof-of-principle study. In addition to its role in myeloid leukemia, we have shown that C/EBPα is necessary for lung alveolar cell development, and downregulation of C/EBPα is detected in approximately half of primary human lung cancers.8,25,26 Thus, our C/EBPα-inducing strategy may find applications in the treatment of solid tumors as well. Further studies are necessary to demonstrate the clinical effectiveness and safety of this class of chemical compounds.
We demonstrated that ICCB280 exhibits antileukemic properties, including terminal differentiation, proliferation arrest, and apoptosis through activation of C/EBPα and affecting its downstream targets. One such target is C/EBPε, which is transcriptionally and directly regulated by C/EBPα and responsible for progression of later stages of granulocytic differentiation of myeloid cells. 21 In addition to transcriptional transactivation of neutrophil-specific genes, such as CD11b and G-CSFR, C/EBPα also hinders cell cycle progression by inhibiting cyclin-dependent kinase 2 (CDK2) through physical interaction with p21. 27 ICCB280-mediated inhibition of cell proliferation can be also explained by a dramatic downregulation of c-Myc by ICCB280, consistent with our previous report demonstrating C/EBPα-mediated transcriptional downregulation of c-Myc gene expression. 14 As Myc proteins contribute to cell proliferation by activating the cell cycle and inducing DNA replication, 28 our results indicate that downregulation of c-Myc contributes to ICCB280-induced proliferation arrest. Furthermore, ICCB280 caused 80% reduction in cell growth in 48 h, whereas differentiation and apoptosis were observed at much later time (5–7 days). These results indicate that growth arrest precedes differentiation and apoptosis induced by ICCB280.
Although hematopoietic cytokines are essential for differentiation of normal hematopoietic stem and progenitor cells into mature cells, only modest effects are observed in leukemia cell lines or primary human AML cells, both in vitro and in vivo. 24 One of the reasons may be lack of signals from G-CSF due to low expression of its receptor, G-CSFR. Indeed, G-CSFR is restricted in a subset of myelomonocytic cell lines and shows considerable variability among primary AML blasts. 29 We showed that ICCB280 induced upregulation of G-CSFR in HL-60 cells, which do not express appreciable amounts of G-CSFR on the surface under normal conditions. 29 It is of interest to note that G-CSF significantly enhanced ICCB280-induced differentiation of leukemia cells. Therefore, a combination of chemical compounds that induce C/EBPα activity and hematopoietic cytokines such as G-CSF makes it a plausible strategy to induce differentiation of leukemic blasts more effectively.
The biological activity of an organic compound depends on its interactions with its biological target, and this interplay is profoundly affected by stable 3D structures presented by the organic molecule. Our conformational analyses imply that ICCB280 adopts a unique spatial geometry that might be preferential for its high activity. We envision that the torsional angle and associated conformational change may prove to be critical parameters to guide a future structure-activity relationship (SAR) study, which may result in novel compounds with improved activity.
In summary, we successfully established a cell-based HTS to identify chemical compounds capable of inducing myeloid differentiation. Our data showing that ICCB280 was capable of inducing differentiation and apoptosis of ATRA-resistant patient blasts strongly signify that the activity of this compound can overcome resistance to other current therapies for AML with an unfavorable prognosis. It will be interesting to test the potential synergistic effect of ICCB280 and other agents affecting granulocytic differentiation, such as ERK1/2 or cdc2 inhibitors.13,30 Finally, our data indicate that similar target-oriented drug screening approaches may be applied to other malignancies affecting different specific pathways. It is expected that identification of novel differentiation therapies will benefit patients with AML who are resistant to current treatment in the near future.
Footnotes
Acknowledgements
We are grateful to Drs. Nicola Tolliday, Jared Shaw, and Caroline Shamu at the Institute of Chemistry and Cell Biology (Harvard Medical School) for the help with the drug screening and Dr. Nicholas Lawrence at Moffitt Cancer Center for providing reagents and helpful discussion.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by National Institution of Health grants (K01DK62064 to H.S.R. and R01CA169259 and R21CA17830 to S.S.K.), Scholar in Medicine Award at Harvard Medical School to H.S.R., Department of Surgery at Beth Israel Deaconess Medical Center to L.S., and Uehara Memorial Foundation, an American Cancer Society grant (RSG-13-047), and Harvard Stem Cell Institute Blood Program Pilot Grant (DP-0110-12-00) to S.S.K.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.