
Abstract
A stable and inducible expression of metabotropic glutamate receptor type 4, 7, and 8 was obtained in T-REx 293 cells using the tetracycline system. Tetracycline administration to the cell medium resulted in rapid induction and time-dependent expression of mGlu receptors, which also correlates with its functionality in a cAMP accumulation assay. The pharmacological properties of recombinant mGlu receptors were verified using orthosteric and allosteric ligands. Data suggest that the Tet-on inducible system is suitable for functional mGlu receptors’ expression and characterization by means of the cAMP accumulation assay. It makes this system a precise, reproducible, and large-scale screening method, as well as a reasonable tool to study signaling properties of mGlu receptors.
Keywords
Introduction
The metabotropic glutamate (mGlu) receptors play an important neuromodulatory role throughout the brain by regulation of cell excitability and synaptic transmission. 1 They are G protein–coupled receptors (GPCRs) with seven-transmembrane (7TM) α-helices interconnected by three extracellular and three intracellular loops. Until now, eight receptor subtypes (mGluR1-8) have been cloned and divided into three major groups (mGlu I, II, and III). Group I includes mGlu1 and 5, group II contains mGlu2 and 3, and group III includes mGlu4, 6, 7, and 8 receptors. In contrast to group I, which is mainly expressed at postsynaptic sites and associated with Gq/11 type protein and phospholipase C, groups II and III are primarily localized presynaptically, linked to Gαi/o protein, and negatively coupled to adenylyl cyclase. 2
Among the group III mGlu receptors, which regulate neurotransmitter release, mGlu4 receptor plays a key modulatory role in many CNS and non-CNS pathways. Intervention in glutamatergic neurotransmission through the mGlu4 receptor has been proposed for the treatment of many neurodegenerative diseases, such as Parkinson disease, Alzheimer disease, stroke, and epilepsy. 3 The role of the mGlu4 receptor has proved itself to be also related with psychiatric disorders such as anxiety 4 and schizophrenia.5,6 Thus, new mGlu4 receptor ligands, especially allosteric modulators with improved selectivity and reduced side effects, have become a very attractive target for therapeutic intervention.7–9 Since only a few ligands for the mGlu4 receptor type are known, there is a need for developing and identifying more potent and specific compounds that can be used in clinical trials.
Expression of mGlu receptors in heterologous mammalian cells is a method for their functional characterization and a tool to study the selective agonist, antagonist, and allosteric effects. For the expression of many GPCRs, including mGlu receptors, the HEK293 cell line derived from a human embryonic kidney is commonly used. Besides the low maintenance costs and appropriate cell division rate, it provides a high efficiency of transfection and protein production. 10 The commercially available HEK293 cell line comes from original transformation, as described by Graham et al. 11 Although the generated cell line was morphologically characterized as epithelioid, the pattern of intermediate filament protein expression (NF-M, NF-L, NF-H, α-internexin) was similar to early differentiating neurons and to both PC12 and Ntera-2 cell lines, themselves defined as neuronal. 12 Moreover, the messenger RNA (mRNA) level of numerous GPCRs and GPCR signaling-related genes endogenously expressed in HEK293 cells was identified. 13 It makes this cell line a convenient environment to express most GPCRs, including mGlu receptors, and to study their signaling without the need for cotransfection.
Since the maintenance of constitutive mGlu receptors’ expression in mammalian cells is problematic,14,15 a stable cell line with inducible expression seemed to be a better solution. We decided to use T-REx 293 cells with an inducible expression system, also containing a cleavage site for the Flp recombinase (FRT). As a result, the transgene is incorporated into a specific site of the genome under the control of a tetracycline-regulated promoter. T-REx 293 cells are characterized by a constitutively produced tetracycline repressor (TetR), which inhibits the transcription from the cytomegalovirus (CMV) promoter. The presence of tetracycline results in repressor binding and its inactivation. In consequence, the operator is released and the protein of interest is expressed.
In the present study, we describe the generation and characterization of stable cell lines with an inducible expression of mGlu receptors as a suitable method for functional screening experiments with the use of cAMP accumulation assay.
Materials and Methods
Plasmid Constructs
DNA fragments encoding variant 1 (canonical sequence) of human metabotropic receptors group III—mGlu4 receptor (NM_000841), mGlu7 receptor (NM_000844.2), and mGlu8 receptor (NM_000845)—were supplied from the University of Missouri–Rolla (pcDNA3.1-GRM4, pcDNA3.1-GRM8) and OriGene (Rockville, MD) (pCMV6-XL4-GRM7). The coding sequences were subcloned into the multicloning site of the pcDNA5/FRT/TO vector (Invitrogen, Carlsbad, CA), under the control of a tetracycline-regulated promoter (CMV/TetO2) using proper restriction enzymes (mGlu4; BamHI and PmeI, mGlu7; NotI, mGlu8; HindIII and ApaI). After the restriction analysis of the received plasmids, the transfection of T-REx 293 cells was performed.
Cell Culture and Transfection
The HEK293 cell line containing an inducible expression system (Flp-In T-REx 293 cell line; Invitrogen) was maintained in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (tetracycline free), 2 mM Glutamax I (Lonza, Basel, Switzerland), and 100 µg/mL Zeocin and 10 µg/mL blasticidin (Invitrogen). For stable transfection, T-REx 293 cells were treated with a mixture of plasmids (0.1 µg pTet-hGRM4, 7, or 8 and 0.9 µg pOG44) and GeneJuice transfection reagent according to the manufacturer’s instructions (Novagen, Madison, WI). After 48 h, the cells were grown in DMEM supplemented with 100 µg/mL hygromycin B (Invitrogen), allowing for the selection of cells with stably integrated pTet-hGRM4, 7, or 8 plasmid. Expression of mGlu receptors was induced by the addition of 0.75 µg/mL tetracycline (final concentration) to the cell medium. The time of cell induction was performed for 24 h with regard to reverse transcription (RT)–PCR analysis. In the case of Western blotting and cAMP accumulation experiments, the cells were induced for 2, 4, 8, 24, 48, or 72 h.
RT-PCR
Stably transfected cells were grown to 60% confluence in DMEM supplemented with 10% fetal bovine serum and 2 mM Glutamax I. Then, protein synthesis was induced by 0.75 µg/mL tetracycline treatment. After 24 h of mGlu4, 7, and 8 receptor induction, the cells were washed twice with phosphate-buffered saline (PBS) and collected in RNA isolation reagent (Trizol; Invitrogen). Following this, DNA contamination removal and reverse transcription reaction were performed according to the manufacturer’s instructions (QuantiTect Reverse Transcription Kit; Qiagen, Hilden, Germany). PCR with specific primers ( Table 1 ) was carried out using Taq PCR Master Mix Kit (Qiagen). The results were visualized on 1.5% agarose gel stained with propidium iodide.
Sequences of Designed Starters, the PCR Conditions, and the Size of Amplicons.

Western Blotting
After growth in a medium with tetracycline, stably transfected cells with pTet-hGRM4 plasmid were washed twice with PBS and solubilized in lysis buffer containing 20 mM Tris-HCl, 100 mM NaCl, 1 mM EDTA, 0.5% Triton X-100, and protease inhibitor cocktail (Thermo Scientific, Waltham, MA) for 20 min on ice. After centrifugation at 13,000 × g for 15 min at 4 °C, supernatants were mixed with 2× Laemmli buffer and heated at 95 °C for 5 min. Then, 10 µg of proteins obtained from cell lysates was separated on 8% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) gels and transferred to nitrocellulose membranes according to the standard Western blot procedure (Invitrogen). After transfer, the membranes were incubated in TBST (Invitrogen) with 5% nonfat milk (BioRad, Hercules, CA) at 4 °C overnight. Then, incubation with primary antibodies for mGlu4 receptor and β-actin (ab53088 [AbCam, San Francisco, CA]; A5441 [Sigma, St. Louis, MO]) was carried out at room temperature for 2.5 h. The secondary horseradish peroxidase (HRP)–conjugated antibodies were incubated with membranes at room temperature for 1 h (W402B and W401B; Promega, Madison, WI). Detection was performed using Pierce ECL Western blotting substrate (Thermo Scientific) and a Fujifilm (Tokyo, Japan) LAS-4000 analysis system.
Forskolin-Induced cAMP Production Assay
A homogeneous time-resolved fluorescence (HTRF) cAMP dynamic 2 assay was performed according to the manual’s instruction from Cisbio (Codolet, France). Stably transfected cells were grown to 60% confluence in supplemented DMEM. Several hours before the experiment (2, 4, 8, 24, 48, or 72 h), mGlu receptor expression was induced by adding 0.75 µg/mL tetracycline. Twenty hours before the experiment, fetal bovine serum (FBS) and Glutamax I deprivation was performed. After the induction step, the cells were scraped and centrifuged. A cell pellet was suspended in Hanks-HEPES buffer containing 130 mM NaCl, 5.4 mM KCl, 1.8 mM CaCl2, 0.8 mM MgSO4, 0.9 mM NaH2PO4, 20 mM HEPES, and 3.25 mM glucose (pH 7.4). Then, 50 µL of cell suspension was pipetted to a 96-well plate containing a final concentration of 10 µM forskolin (in the case of the mGlu7 receptor, 3 µM forskolin) added with the appropriate concentration of compound and/or L-glutamate. The cell suspension was incubated together with forskolin and compounds at 37 °C for 5 min. Following this, 10 µL of the cell suspension was transferred to a 384-well plate with 5 µL of cAMP-d2 conjugate in lysis buffer by means of an automated pipetting system (Tecan Evo 200; Tecan, Mannedorf, Switzerland). Then, 5 µL of anti-cAMP cryptate conjugate was added. After 1 h of incubation at room temperature, the fluorescence at 620 nm and 665 nm was read (Tecan Infinite M1000). The results were calculated as the 665-nm/620-nm ratio multiplied by 104. The specific signal was inversely proportional to the concentration of cAMP in the sample.
Compounds
L-glutamate was acquired from Sigma. L-(+)-2-Amino-4-phosphonobutyric acid (L-AP4), cis-2-[[(3,5-Dichlorophenyl)amino]carbonyl]cyclohexanecarboxylic acid (VU0155041), 6-(4-Methoxyphenyl)-5-methyl-3-(4-pyridinyl)-isoxazolo[4,5-c]pyridin-4(5H)-one hydrochloride (MMPIP) were purchased from Tocris Bioscience (Bristol, UK) and 2-(4-bromobenzylthio)-N-(4-sec-butylphenyl)acetamide (AZ12216052) was from Axon Medchem (Groningen, the Netherlands). L-Glu and L-AP4 were dissolved in Hanks-HEPES buffer. Reference allosteric compounds were dissolved in DMSO and used in experiments as a 3-µM VU0155041, 1-µM MMPIP, and 10-µM AZ12216052 concentration.
Data Analysis
The concentration-response curves of L-glutamate and representative compounds were generated using the nonlinear regression. The curves were fitted to a 3-parametric logistic equation, allowing for the determination of EC50 values. Analyses of the variance (one-way ANOVA) and post hoc Tukey test were used to compare the means of the mGlu4 receptor expression level depending on the time of induction. GraphPad Prism version 5.04 was used for analysis (GraphPad Software, San Diego, CA).
Results
RT-PCR Analysis of mGlu4, 7, and 8 Receptors Expression in Isolated Clonal Cell Lines
T-REx 293 cells were stably transfected with the human mGluR4, 7, and 8 coding sequences subcloned into a pcDNA5/FRT/TO vector. After transfection, hygromycin-resistant cell clones were selected for each receptor. All of the clones were analyzed with respect to mRNA (RT-PCR) and protein (Western blotting) expression and, in particular, receptor functionality (cAMP production assay). Receptor-specific clones with the highest transgene expression and the proper EC50 value for L-glutamate were chosen. The mRNA expression in those clonal cells was determined after a 24-h incubation with tetracycline. Nontransfected cells were used as a control ( Fig. 1 ).
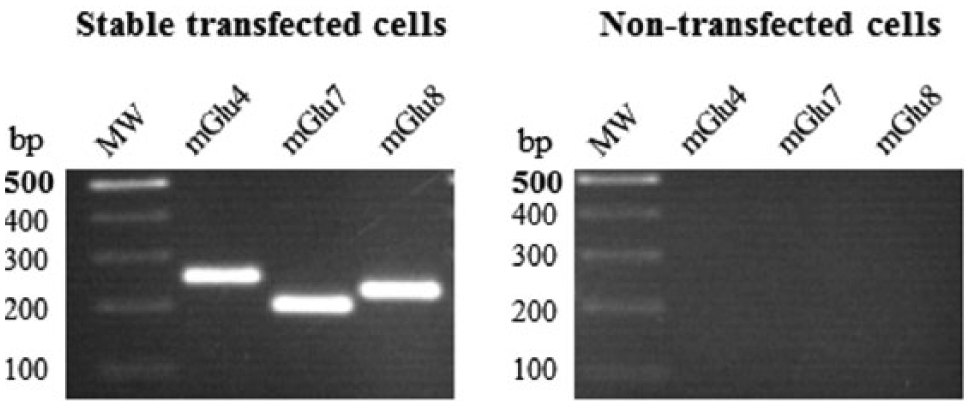
Reverse transcription–PCR analysis of mGluR4, 7, and 8 receptor expression in stable transfected cell lines after 24 h of Tet induction and in nontransfected T-REx 293 cells.
Characterization of mGlu4 Receptor Inducible Cell Line
The kinetics of mGlu4 receptor expression and functionality were examined dependent on the time of cells’ induction. Cells were collected after 0, 2, 4, 8, 24, 48, and 72 h of incubation with tetracycline. The expression of the mGlu4 receptor at the protein level was analyzed by immunoblotting. The functionality of the receptor was tested by the forskolin-induced cAMP production assay, where the specific signal was inversely proportional to the concentration of cAMP in the sample.
Statistically significant differences in the time-dependent mGlu4 receptor expression were observed ( Fig. 2A ). Although mGlu4 receptor protein was barely detectable after 2 h of induction, the extensive increase of the luminescence signal was seen within 4 h. The highest level of mGlu4 receptor expression was reached after 24 h of cell induction. After 48 h of incubation with the antibiotic, the protein level started to decrease ( Fig. 2B ). The functionality of the mGlu4 receptor, similar to the protein expression, was dependent on the cell induction time ( Fig. 2C ). Stimulation of adenylyl cyclase with forskolin resulted in an increased level of accumulated cAMP after cell induction longer than 8 h. In the case of cells grown in a medium with tetracycline for 2 h and 4 h, the cAMP level was lower ( Fig. 2D ). EC50 were consistent with literature only for 48 h and 72 h of induction of cells, reaching values lower than 10 ( Table 2 ). After a longer time of tetracycline treatment, the concentration-response curve for L-glutamate was characterized by a wider span; therefore, in further experiments, mGlu4 receptor-expressing cells were induced for 72 h.
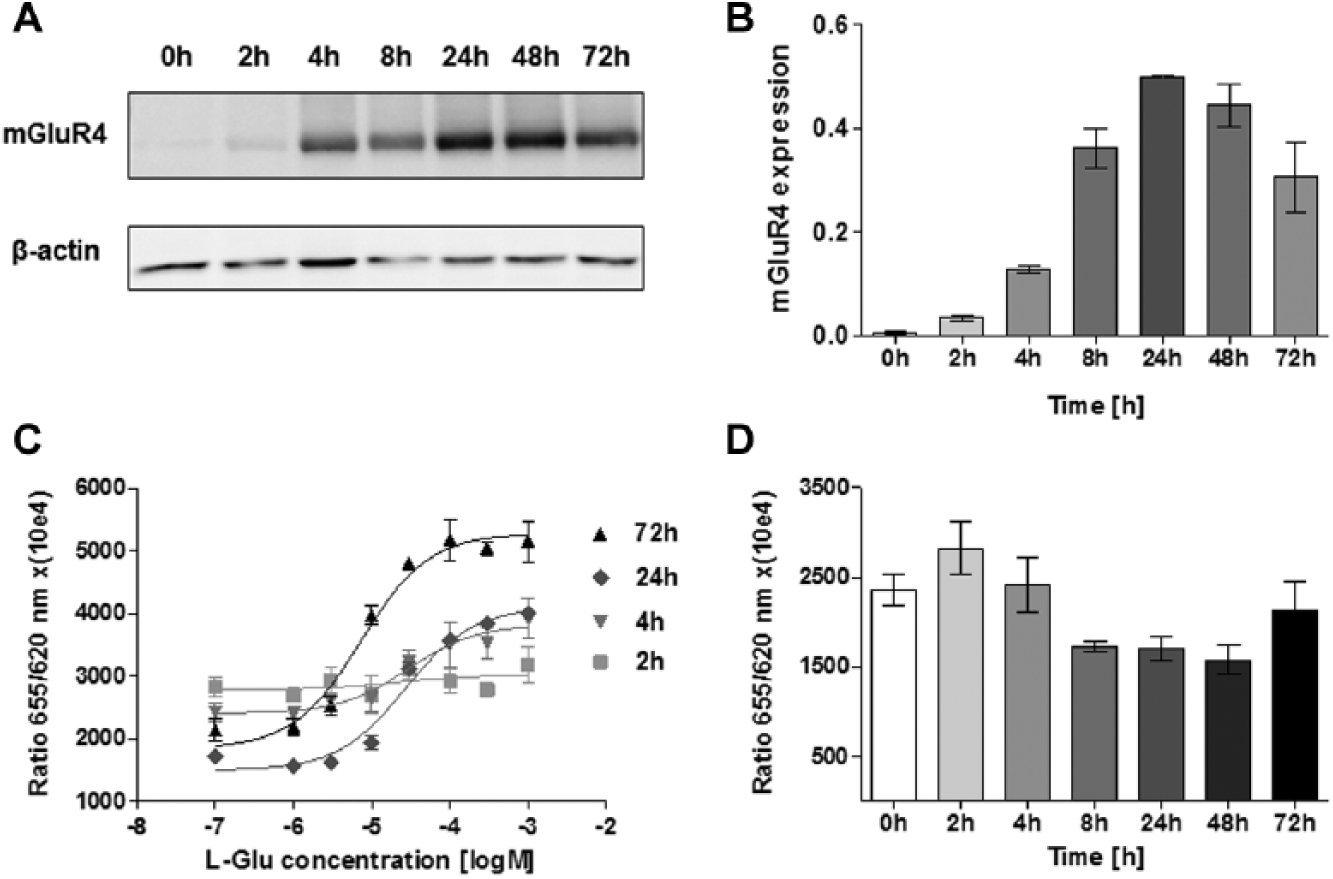
The kinetics of mGlu4 receptor expression and functionality dependent on the time of cells’ induction. (
EC50 and Span Values of L-Glutamate Dose-Response Curves Depending on Time of mGlu4 Receptor Cell Induction and in Cells with mGlu4 Receptor Expression after 72 h of Tet Induction (0%, 20%, 40%, 60%, 80%, and 100% of mGlu4-Expressing Cells) Mixed with Nontransfected Cells (Values from a Single Experiment Run in Triplicate).
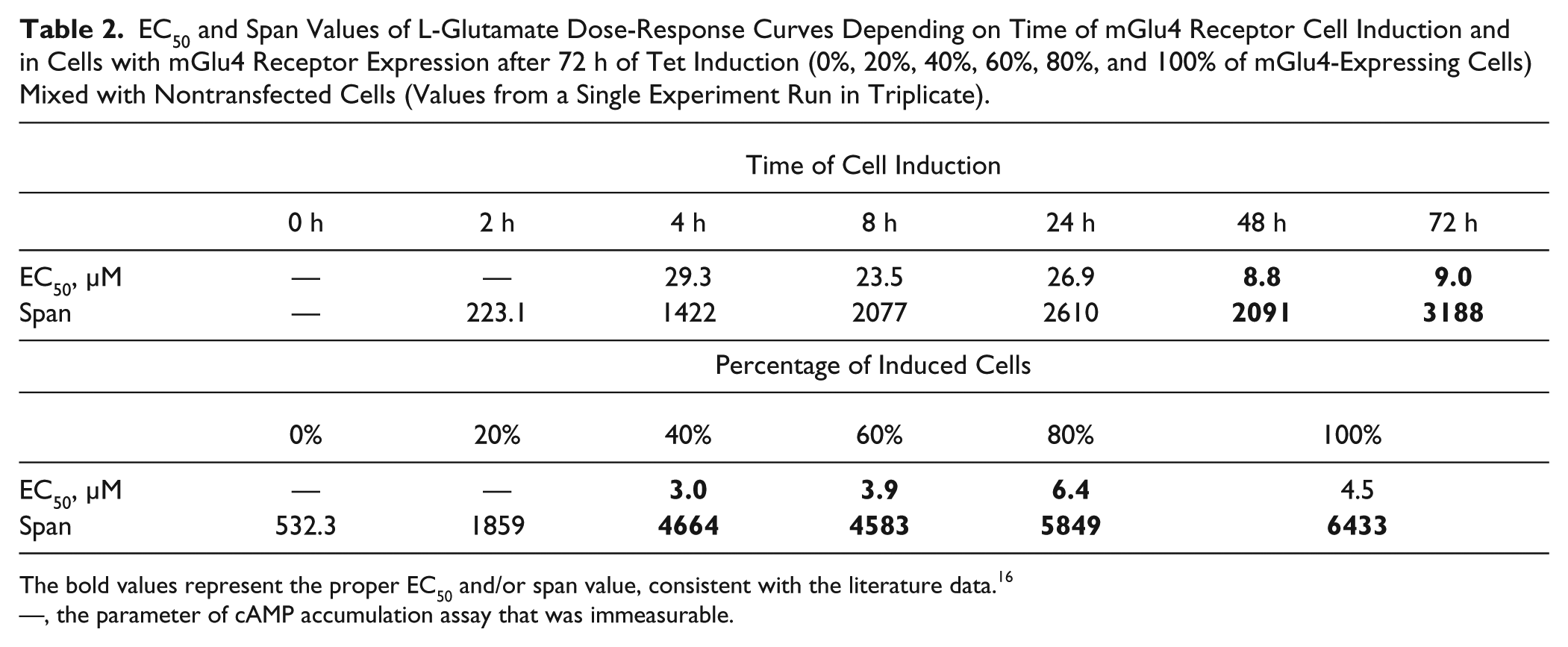
The bold values represent the proper EC50 and/or span value, consistent with the literature data. 16
—, the parameter of cAMP accumulation assay that was immeasurable.
To examine the correlation between the expression level of the mGlu4 receptor and its functionality, we mixed hGRM4–T-REx 293 cells after 72 h of induction and nontransfected T-REx 293 cells in various ratios. When a greater percentage of mGlu4 receptor-induced cells was used, a lower EC50 and a wider span of the concentration-response curve for L-glutamate were observed ( Fig. 3A ). A proper EC50 value was obtained for 40% of the mGlu4 receptor-induced cells and did not change significantly for 60%, 80%, or 100% of these cells. The span increased simultaneously with the rise of mGlu4 receptor cells after induction and reached the highest value for 100% of these cells ( Table 2 ). The endogenous cAMP level accumulated by stimulation of adenylyl cyclase increased also in cell mixes containing 40% or more concentration of mGlu4 receptor-induced cells ( Fig. 3B ).
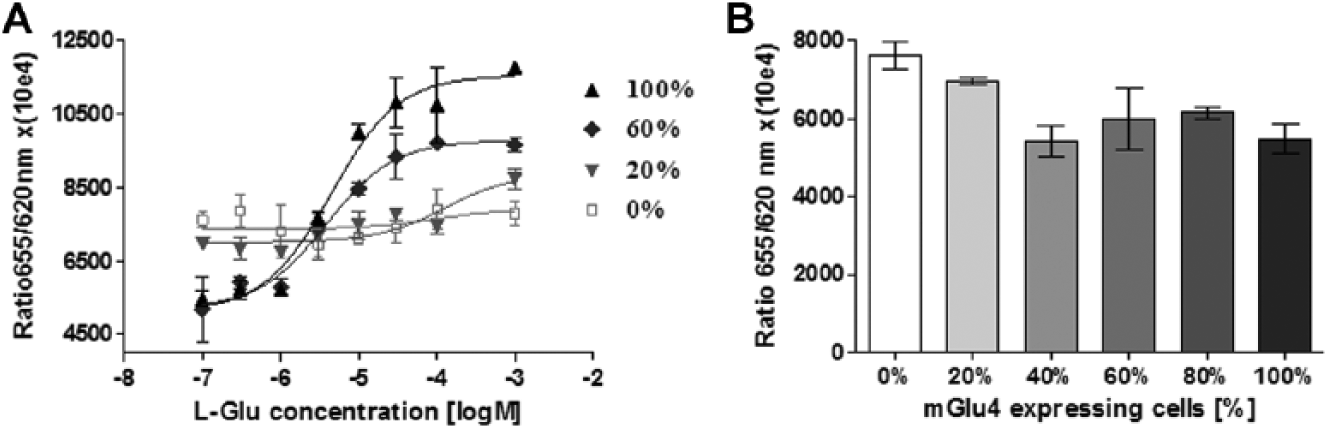
Changes in the mGlu4 receptor functionality dependent on the amount of induced cells. (
Functional Characterization of mGlu Receptor Group III Using Reference Compounds
The receptor functionality was measured by means of a forskolin-induced cAMP production assay after 72 h of mGlu4 receptor cell induction. In the case of mGlu7 and mGlu8 receptor cells, a 48-h incubation with tetracycline was sufficient. In all functional experiments, L-glutamate, the endogenous neurotransmitter, was used as a reference orthosteric agonist. To confirm the activation of mGlu4, 7, and 8 receptors in the obtained cell lines, we conducted the analogous experiments in the presence of L-AP4, a synthetic orthosteric agonist highly selective for group III mGlu receptors. L-glutamate and L-AP4 decreased the forskolin-induced cAMP production in the mGlu4 receptor cell line with EC50 = 6.7 ± 0.6 µM and EC50 = 0.1 ± 0.05 µM, respectively. 16 Similarly, the used orthosteric agonists inhibited the forskolin-induced cAMP accumulation with EC50 = 554 ± 183 µM and EC50 = 71 ± 24 µM in mGlu7 receptor cells 17 and an EC50 = 4.6 ± 1.0 µM and EC50 = 0.14 ± 0.04 µM in the mGlu8 receptor cell line. 18 The obtained EC50 values for orthosteric ligands (mean ± SD from four independent experiments) were consistent with the literature in all tested cell lines ( Fig. 4 ).
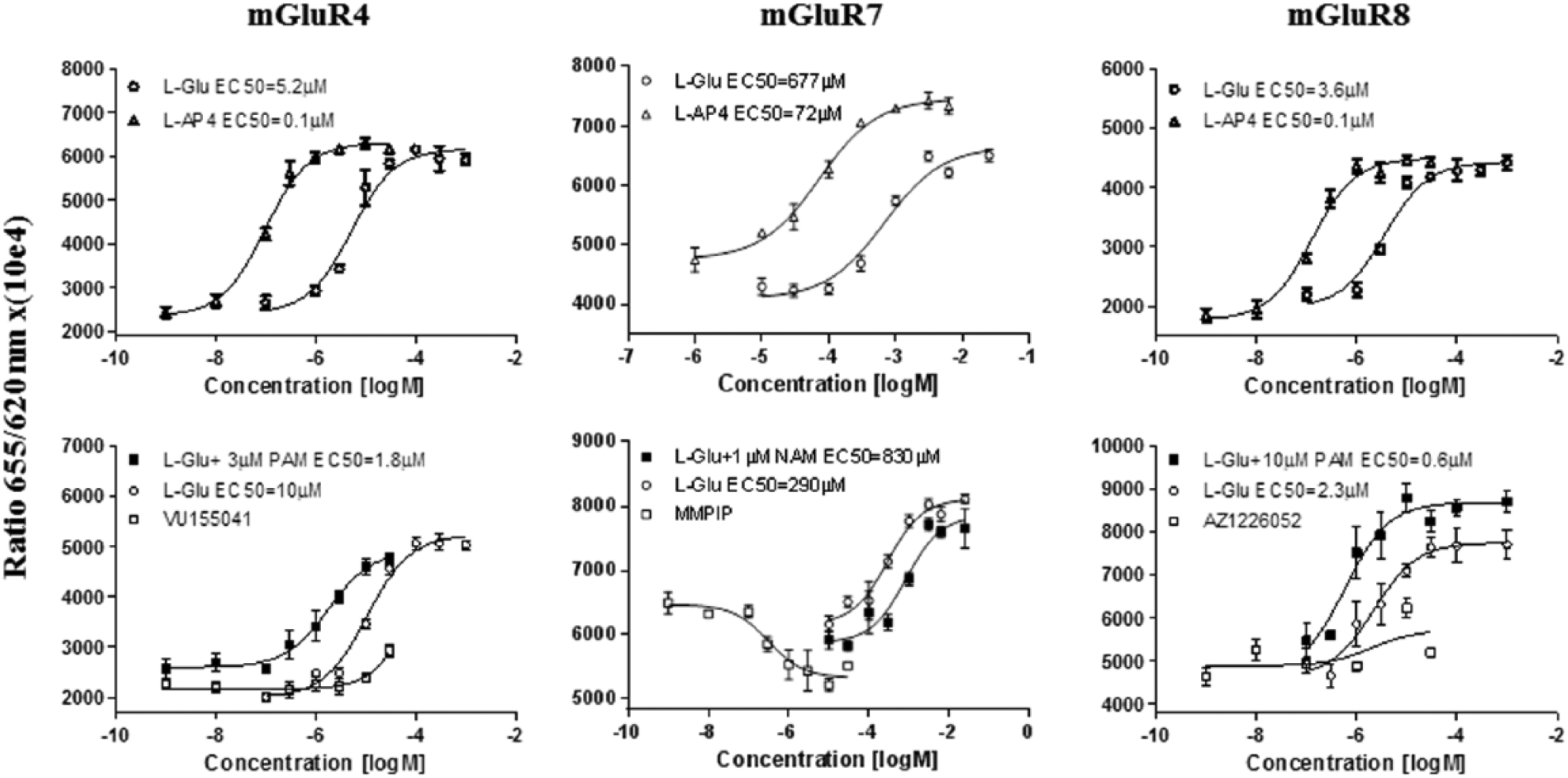
Representative concentration-response curves for the endogenous ligand (L-glutamate) and specific for the group III synthetic ligand L-AP4 in mGlu4, mGlu7, and mGlu8 receptor cells (top panel). Potentiator activity of reference allosteric modulators (VU0155041 for mGlu4 receptor, MMPIP for mGlu7 receptor, and AZ12216052 for mGlu8 receptor) demonstrated as representative concentration-response curves for L-glutamate in the absence and presence of each reference allosteric modulator (bottom panel). The mGlu4 cells were induced for 72 h; mGlu7 and mGlu8 receptor cells were induced for 48 h.
Because of our interest in allosteric modulation, several compounds with known allosteric modulator activity were analyzed. Concentration-response curves for L-glutamate in both the absence and presence of each reference modulator were made. Moreover, the concentration-response curves for each modulator in the absence of the orthosteric agonist were performed. The positive allosteric modulator VU0155041 potentiated the response to the orthosteric agonist in the mGlu4 receptor cell line by shifting the dose-response curve of L-glutamate to the left with EC50 = 1.2 ± 0.4 µM (mean ± SD from three independent experiments). 3 AZ12216052 induced a leftward shift of the L-glutamate concentration-response curve in the mGlu8 receptor cell line with EC50 = 1.0 ± 0.4 µM (mean ± SD). 19 In contrast, the negative allosteric modulator MMPIP 20 raised the basal cAMP level and, in the 1-µM final concentration, decreased the activity of L-glutamate in a noncompetitive fashion with a slight change in the EC50 value of the L-glutamate dose-response curve (EC50 = 1016 ± 261 µM, mean ± SD). 21 The results were consistent with the available literature data ( Fig. 4 ).
Discussion
In most receptor studies conducted in vitro in cell lines, the fundamental thing is to establish the heterologous expression of the gene of interest. Transient expression is an acceptable and commonly used model for the evaluation of receptor pharmacological properties, function, signal transduction, or rapid screening of mutagenesis products. On the other hand, low throughput, high costs of repeated transfections, and significant changes in the expression level among the population of cells and experiments make this model insufficient. For large-scale screening studies, generation of constitutively expressing stable cell lines is necessary. However, the maintenance of mGlu receptors stably expressed in mammalian cells is problematic.15,22 It can be partially related to glutamate (the endogenous mGlu receptor agonist) secretion to the medium and persistent activation of transfected mGlu receptors. 14 Based on our experience with mGlu4, 7, and 8 receptor expression, after a few weeks of antibiotic selection, the stably transfected cells were characterized by poor adhesion, rounded shape, and low cell division rate. This suggested that stable mGlu receptor overexpression might have detrimental impacts on HEK293 cells’ growth and viability.
To overcome this problem and downregulate the expression of receptors during the maintenance of the cells, we created HEK293 cell lines with stable and inducible mGlu receptor expression using the Flp-In T-REx system. The coding sequences of mGlu4, 7, and 8 receptors were cloned under the control of a tetracycline-regulated promoter (CMV/TetO2), and their transcripts were confirmed in isolated clonal cells. In conventional stably transfected cells, an expression of the gene of interest is constant, and in the Tet-on system, its transcription is turned on after tetracycline induction (Tet-on system). Cells can be grown to the required density before antibiotic administration, which significantly reduces the potential transgene “toxicity.” Moreover, a tetracycline-responsive promoter should provide rapid and tight control of transgene expression, unlike the constitutive expression, where any changes are beyond the experimenter control. 23 Nevertheless, the low responsiveness of the tetracycline expression system in HEK293 cells was postulated by Howe et al. 24 Using the Tet-off system, the authors observed a minimal response to the tetracycline of the Glu6 receptor in HEK293 cells, in contrast to the D3 receptor expression tightly controlled in the GH3 cell line. In contrast to Howe et al., we observed statistically significant differences in the mGlu4 receptor expression in T-REx 293 cells (modified HEK293 cells), dependent on the time of cell induction. The extensive increase of the luminescence signal was observed within 8 h, the peak was reached after 24 h, and after 48 h of cell induction, the protein level started to decrease. Moreover, after tetracycline elimination from the medium, the mGlu4 receptor expression was silenced within 56 h (data not shown). The barely detectable background level of the mGlu4 receptor and its high signal-to-background ratios obtained for the Tet-on system was also observed by Downey et al. 25 for the ecdysone-based system expressing the mGlu5 receptor. Although it seems that the promoter’s responsiveness mainly depends on the cell’s genetic background, it can be also determined by the type of inducible system (IPTG, Ecdysone, Tet-on, Tet-off), the exogenous-inducing agent (muristeron, ponasterone A, tetracycline, doxycycline), and the protein of interest.
The group III mGlu receptors are linked to the Gαi/o protein and negatively coupled to adenylyl cyclase. As a result, activation of the receptors inhibits cAMP formation. Thus, receptor functionality can be measured by means of a forskolin-induced cAMP production assay, which was applied in this study. The influence of the mGlu4 receptor expression level on cAMP accumulation assay parameters was tested for the different tetracycline induction times. In this experiment, higher receptor expression improved both the EC50 and the span values for L-glutamate, which confirmed the direct correlation between the amount of mGlu4 receptors and their functionality. Since the proper parameters were reached after 48 h and 72 h of cell induction, the receptor functionality therefore contrasts with the receptor expression, where the highest values were apparent after 24 h. The receptor functionality could be also related to the time needed for its adequate posttranslational processing and delivery to the cell membrane, which is problematic in the case of many GPCRs expressed in heterologous cells. 26 This hypothesis was confirmed in the experiment with the mGlu4 receptor-expressing cells mixed in various ratios with nontransfected T-REx 293 cells. The acceptable EC50 value of the L-glutamate concentration-response curve was noticed in 40% of the mGlu4 receptor-induced cells. No significant changes in this parameter were observed in cell mixes containing 60%, 80%, or 100% of the mGlu4 receptor-expressing cells. In contrast to the EC50 values, the span increased with the rise of mGlu4 receptor-induced cells in the mixes and reached the highest value in 100% of these cells. This result can be partly related to the increase in cAMP level after stimulation by forskolin observed in mixes with the greater amount of induced hGRM4–T-REx 293 cells. Analogous changes in endogenous cAMP level were noticed after a different time of cell induction. Although adenylyl cyclase is activated directly by forskolin beyond the receptor, the increase of endogenous cAMP level is correlated with the greater amount of mGlu4 receptor in the cells. These results can be derived from dual regulation of adenylyl cyclase described for the µ-opioid receptors activated by morphine. 27 Similar to group III mGlu receptors, the opioid receptors are linked to G proteins and negatively coupled to adenylyl cyclase. In consequence, morphine stimulation inhibits adenylyl cyclase. Nevertheless, an exposure of the µ-opioid receptors to morphine longer than 12 h resulted in a relatively stable increase in adenylyl cyclase–specific activity. Late positive regulation of the enzyme, as well as changes in activity of other downstream effectors, is responsible for the narcotic dependence and tolerance. 28 The mGlu receptors can be activated during tetracycline treatment by amino acids other than L-Glu contained in cell medium. Continuous low activity of the mGlu receptors analogous to the opioid receptors can cause the positive regulation of adenylyl cyclase activity. In consequence, after a longer time of induction, as well as in mixes with greater amount of mGlu4 receptor-induced cells, an endogenous cAMP level tends to increase.
Taking into account the above results, the best assay conditions were established. Thus, the pharmacological properties of mGlu receptors, type 4, 7, and 8, in the obtained cell lines were verified after 48 h or 72 h of mGlu receptor induction. The mGlu receptors, type 4, 7, and 8, were stimulated by L-glutamate, the endogenous neurotransmitter, and L-AP4, the selective agonist for the group III mGlu receptors. The results were consistent with the literature for those two orthosteric agonists as well as for allosteric modulators. VU0155041 and AZ12216052 were effective in shifting the glutamate concentration-response curve in mGlu4 and mGlu8 receptor cell lines, respectively, while MMPIP acted through negative cooperativity with L-glutamate in mGlu7 receptor cells. These effects indicate that pharmacological properties of the mGlu receptors in this system are unaltered and can be used for the identification and characterization of novel orthosteric as well as allosteric compounds. Moreover, establishing a similar condition for mGlu4, 7, and 8 receptor cell lines in the same experimental system provides an essential tool for the discovery and valuation of type selective drugs.
In our system, receptor functionality is measured by means of HTRF-based cAMP detection. The obtained signal is based on the energy transfer between donor (cAMP antibody with europium cryptate) and acceptor (cAMP labeled with a modified allophcocyanin dye, d2) with time-resolved fluorescence resonance energy transfer (TR-FRET). The unique caged structure of cryptate provides higher signal stability and therefore allows for enhanced assay sensitivity. Time-resolved measurement of fluorescence allows the elimination of short-lived background fluorescence from buffers and media compositions. Also, interferences caused by different photophysical properties of tested compounds are minimized by dual-wavelength detection. 29 In contrast to the GloSensor method, which measures cAMP production in real time, the time-resolved measurements of fluorescence are characterized by higher sensitivity, robustness, and a low compound interference rate. 30 Thus, the system under investigation allows rapid measurements of ligand activity in a high-throughput screening format with the use of all three mGlu receptor cell lines simultaneously.
In most cases concerning studies of mGlu receptors, transient transfected cells are used. Only two systems with inducible mGlu receptor expression have been described so far: the IPTG-based system expressing the mGlu1 receptor 22 and the ecdysone system for mGlu4, 5, and 8 receptor expression. 25 Although both systems give tightly controlled receptor expression, the highest protein level is obtained via the Tet-on system. Moreover, tetracycline doses necessary to induce the Tet-on system are much lower and nontoxic, in contrast to the chemical agents for ecdysone system induction. 31 Thus, the generation of stably transfected T-REx 293 cells using the Tet-on system and validation of the high level and controlled expression of functional mGlu receptors combined with highly sensitive and effective cAMP detection assay enabled us to establish a reasonable and high-throughput screening platform for the identification and pharmacological characterization of new chemical compounds besides being a great tool to study signaling properties of the mGlu receptors.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by projects No UDA-POIG.01.03.010-12-100/08-00 and No K/ZDS/00287. Moreover, Barbara Chruścicka acknowledges the financial support from the project Interdisciplinary PhD Studies “Molecular sciences for medicine” (co-financed by the European Social Fund within the Human Capital Operational Programme).