
Abstract
Human-induced pluripotent stem cell–derived cardiomyocytes (hiPS-CMs) at different stages (approximate days 30, 60, and 90) were used to determine the appropriate stage for functional and morphological assessment of drug effects in vitro. The hiPS-CMs had spontaneous beating activity, and β-adrenergic function was comparable in all stages of differentiation. Microelectrode array analyses using ion channel blockers indicated that the electrophysiological properties of these ion channels were comparable at all differentiation stages. Ultrastructural analysis using electron microscopy showed that myofibrillar structures at days 60 and 90 were similarly distributed and more mature than that at day 30. Analysis of motion vectors in contracting cells showed that the velocity of contraction was the highest at day 90 and was the most mature among the three stages. Gene expression analysis demonstrated that expression of some genes related to myofilament and sarcoplasmic reticulum increased with maturation of morphological and contractile properties. In conclusion, day 30 cardiomyocytes are useful for basic screening such as the assessment of electrophysiological properties, and days 60 and 90 are the appropriate differentiation stage for morphological assays. For the assay of contractile function associated with subcellular components such as sarcoplasmic reticulum, day 90 cardiomyocytes are the most suitable.
Keywords
Introduction
The breakthrough technology introduced by Takahashi and Yamanaka in 2006 enabled the derivation of pluripotent stem cells by reprogramming of somatic cells through introduction into cells of a set of transcription factors.1,2 The human-induced pluripotent stem cells (hiPSCs) closely resembled the previously described human embryonic stem cells (hESCs), 3 including their abilities to self-renew and pluripotency, which enabled them to differentiate into numerous kinds of functional cells.2,4 These iPSC-derived functional cells are expected to be useful in research related to regenerative medicine and as tools for drug efficacy/safety screenings. In addition, phenotypes of iPSC-derived functional cells generated from patients having various disease/disorders could reflect those associated with their disease, and they are expected to be valuable for the investigation of disease mechanisms and the discovery of targets for new drugs.5,6
Many studies have demonstrated that hiPSC-derived cardiomyocytes (hiPS-CMs) have contractile ability and express cardiac-specific genes similar to normal cardiomyocytes.7–9 These cells have received much attention because of their potential as tools for drug efficacy/safety screenings.10,11 For example, in drug safety screening, electrophysiological assays including the human Ether-a-go-go Related Gene (hERG) inhibition and QT prolongation using hiPS-CMs might predict clinical outcomes, such as arrhythmia.12–14 Although these studies demonstrated the utility of hiPS-CMs at day 20 to 40 of differentiation in electrophysiological analyses, it has been reported that functional sarcoplasmic reticulum (SR) for calcium handling in hESC-derived cardiomyocytes at day 25 to 31 of differentiation is less mature than those in the adult heart 15 and that the functional and morphological maturation depends on the duration of culture.16,17 Therefore, to emulate the drug response in the adult human heart more accurately, hiPS-CMs generated after a longer differentiation period are expected to be more valuable for functional and morphological assays than those after a short period of differentiation (day 20–40). However, the functional and morphological characteristics of hiPS-CMs at day 60 of differentiation and beyond, and comparison of the characteristics at different stages of differentiation, have not been described. In addition, although comparison of global gene expression of hESC-derived cardiomyocytes and hiPS-CMs at a specific stage of differentiation with those of undifferentiated hES/hiPS cells and adult heart have been reported,18,19 the similarities and differences of global gene expression of hiPS-CMs at different stages of differentiation and their comparison with that of adult heart have not been shown.
In the present study, to more fully understand the functional and morphological characteristics and global gene expression profiles at various stages of differentiation in culture, we focused on the change in characteristics in hiPS-CMs with period of differentiation and characterized the functional and morphological features and the global gene expression pattern in hiPS-CMs at days 50 to 60 and 80 to 90 of differentiation and compared these to those observed at days 20 to 30 of differentiation and with adult ventricle tissue. We thus determined the appropriate differentiation stage for assessment of drug physiological effects or toxicity.
Materials and Methods
hiPSC Culture and Differentiation into Cardiomyocytes
The hiPSC line (253G1) from Kyoto University was maintained as previously described.2,4 Undifferentiated hiPSCs were maintained on a feeder layer of mitomycin-C–treated mouse embryonic fibroblasts in primate ES medium (ReproCELL, Kanagawa, Japan) supplemented with 4 ng/mL recombinant human basic fibroblast growth factor (bFGF; PeproTech, Rocky Hill, NJ) in a 5% CO2/95% air environment. Differentiation of hiPSCs was induced using the method described previously with slight modifications.20,21 Briefly, the colonies of hiPSCs were isolated in dissociation solution (ReproCELL), and they formed embryoid bodies (EBs) in suspension culture in low cell binding 90 mm dishes (Nunc, Tokyo, Japan) in basic media (StemPro34 [Invitrogen, Carlsbad, CA] containing 2 mM glutamine, 4 × 10–4 M monothioglycerol [Sigma, St. Louis, MO], and 50 µg/mL ascorbic acid [Sigma]). The following cytokines were also used at the relevant differentiation stage: days 0 to 1, bone morphogenetic protein 4 (BMP4; 0.5 ng/mL); days 1 to 4, BMP4 (10 ng/mL), bFGF (5 ng/mL), and activin A (3 ng/mL); days 4 to 8, dickkopf homolog 1 (DKK1; 150 ng/mL) and vascular endothelial growth factor (VEGF; 10 ng/mL); and after day 8, VEGF (10 ng/mL), DKK1 (150 ng/mL), and bFGF (5 ng/mL), all in the basic media described above. All factors were purchased from R&D Systems. Cultures for the differentiation were maintained in a 5% CO2/95% air environment. During differentiation, the medium was replaced every 3 to 4 d, and hiPSC-derived cardiomyocytes were cultured until days 20 to 30, 50 to 60, and 80 to 90 of differentiation (d30CMs, d60CMs, and d90CMs, respectively) and used in the present study. The purification of hiPS-CMs from beating EBs was performed as previously described. 1 At day 14 of differentiation, beating EBs were dissociated by pipetting with a thin glass pipette. On the next day of the dissociation, beating cell clusters, which had obvious large beating regions, emerged from the particles of dissociated beating EBs. We confirmed that the percentage of cardiac troponin T (cTnT)–positive cells in the beating cell clusters at day 90 was 82.4% when 6697 cells were analyzed.
Reverse-Transcription PCR
RNA was isolated with the RNeasy micro kit (Qiagen, Hilden, Germany) from undifferentiated hiPSCs, d30CMs, d60CMs, and d90CMs according to the manufacturer’s instructions. The RNA concentrations were determined by absorbance at 260 nm using a NanoDrop ND-8000 spectrophotometer (NanoDrop Technologies, Wilmington, DE). RNA samples were stored at –80 °C until assay. Reverse-transcription (RT) into complementary DNA and the PCR were carried out on 33 ng of the total RNA from each sample using a BcaBEST RNA PCR Kit (Takara, Shiga, Japan). PCR-related primers of genes that were OCT3/4, NKX2.5, MYH6, and GAPDH are detailed in
Fluorescent Immunostaining for Cardiac Markers
Fluorescent immunostaining was performed on d30CMs, d60CMs, and d90CMs as previously described.21,22 Several d30CMs, d60CMs, and d90CMs were picked up and dissociated using 10 mg/mL collagenase IV for 2 to 3 h at 37 °C. The dissociated cells were seeded onto collagen I–coated dishes. Two or 3 days after seeding the dishes, the cells were fixed in 4% formaldehyde for 30 min and subsequently permeabilized with 2.5% Tween 20 (Wako, Osaka, Japan) at room temperature. The cells were incubated at 4 °C overnight with the following primary antibodies diluted in Dulbecco’s PBS(–) buffer (PBS, Gibco, Invitrogen): polyclonal anti-GATA4 (1:100 dilution; Abcam, Tokyo, UK), monoclonal anti–cardiac actin (1:50 dilution; Fitzgerald, Acton, MA), monoclonal anti-cTnT (1:100 dilution; Abcam), monoclonal anti–myosin cardiac heavy chain 6 (MYH6; 1:100 dilution; Abcam), and polyclonal anti–myosin light chain 2 (MLC2; 1:100 dilution; Abcam). Fluorescent dye–conjugated secondary antibodies, Texas red–conjugated goat anti-rabbit IgG (1:100; Vector Laboratories, Burlingame, CA), FITC-conjugated rabbit anti-mouse IgG (1:40; Dako, Glostrup, Denmark) or FITC-conjugated swine anti-rabbit IgG (1:40; Dako) were applied to the cells for 1 h at room temperature. After the reaction, the cells were washed three times with washing buffer (PBS containing 0.5% Triton-X 100 [Sigma] and 0.5% bovine serum albumin [Sigma]). Nuclei were stained with 5 µg/mL Hoechst 33342 (Sigma) for 10 min at room temperature, and the cells were washed again three times with washing buffer. The specimens were then observed under a Biozero fluorescence microscope BZ-9000 (Keyence, Osaka, Japan).
Effect of Drugs on Spontaneous Contraction
To investigate the effect of drugs on the frequency of beating in d30CMs, d60CMs, and d90CMs, a β-adrenergic agonist, isoproterenol (Calbiochem, San Diego, CA) was selected as a model compound. The number of beats was counted at 20 s intervals for 30 min before and after the addition of 1 µM isoproterenol to the medium.
Assessment of Electrophysiological Function
Microelectrode arrays analysis was performed to investigate the electrophysiological characteristics of d30CMs, d60CMs, and d90CMs using the MED 64 system (Alpha MED Sciences) referred to in previous reports.23–25 These hiPS-CMs were transferred onto the MED-probe dishes (Alpha MED Sciences) and cultured at 37 °C in a humidified atmosphere of 95% air and 5% CO2 for 3 to 7 d to allow attachment. Field potential duration (FPD) of the hiPS-CMs was recorded with this system. The tested drugs included lidocaine, a sodium channel blocker (Sigma-Aldrich); nifedipine, a calcium channel blocker (MP Biomedicals, Japan); and E-4031, an hERG-type potassium channel blocker (Wako) known to cause a QT prolongation. All extracellular recordings were performed for 60 s. Baseline was recorded prior to drug application and after addition of drug recording continued for 30 min at 37 °C.
Transmission Electron Microscopy
Ultrastructural details of hiPS-CMs were investigated as previously described. 22 For transmission electron microscopy, d30CMs, d60CMs, and d90CMs were fixed in 2.5% glutaraldehyde in 0.1 M phosphate buffer (pH 7.2) for 2 h, postfixed in 1% OsO4 in the same buffer for 1.5 h, dehydrated in graded ethanol series, and embedded in Epoxy resin (TAAB, Berkshire, UK). Ultrathin (90–100 nm) sections were cut and double-stained with uranyl acetate and lead citrate and viewed using an H-7600 transmission electron microscope (Hitachi, Tokyo, Japan).
Assessment of Contractile Function
To investigate the contractile velocity in hiPS-CMs, movie images were captured with a BIOZERO fluorescence microscope BZ-9000 (Keyence, Osaka, Japan), and the contractile velocity was analyzed using motion analysis software (VW-H2 MA; Keyence, Osaka, Japan). Initiation points of motion vectors were plotted randomly on a single cell body images, and the velocity of contraction was calculated by dividing the length of a motion vector by time during a contraction. The motion vector that showed the fastest velocity in vectors plotted randomly on a single cell was taken as the velocity of contraction for the cell.
Global Gene Expression Analysis
Gene expression profiles of d30CMs, d60CMs, and d90CMs were compared with those of undifferentiated hiPSCs and adult human ventricle tissue (Cureline, San Francisco, CA). Human iPS-CMs were sampled at each stage three times from the same sources but at different times during the culture stage. The adult ventricle tissue used was obtained from three different cadavers. Total RNA (10 ng) was prepared from each sample and used in the whole-genome DASL (cDNA-mediated annealing, selection, extension, and ligation) assay (Illumina, San Diego, CA) according to the manufacturer’s recommended procedure. The fluorescent intensities were measured by scanning the arrays on a BeadArray Reader (Illumina). Scanned data were then uploaded to a computer and analyzed with BeadStudio software (Illumina) via the gene expression module (whole-genome DASL Assay). The data were further exported to GeneSpring GX version 11.0 (Agilent Technologies, Santa Clara, CA), and gene ontology analysis was performed. The detailed method is described in the supplementary materials.
Statistical Analysis
Statistical analyses to test for significant differences were Student’s t test and Welch’s t test for comparison of two groups with equal variances and unequal variances, respectively. For both tests, values of p ≤ 0.05 were considered statistically significant. All statistical calculations except the data obtained by global gene expression analysis were carried out using the SAS system (SAS Institute, Cary, NC). The analytical methods on the data from global gene expression analyses are described in the supplementary materials.
Results
Differentiation of hiPS Cells into Cardiomyocytes In Vitro
The expression of cardiac marker genes were analyzed by RT-PCR ( Fig. 1A ). OCT3/4 expression was detected in the undifferentiated hiPSCs but not in d30CMs, d60CMs, or d90CMs. Expression of Nkx2.5, which is a cardiac transcription factor, and MYH6, which is a contractile cardiac protein, was detected in d30CMs, d60CMs, and d90CMs but not in undifferentiated hiPSCs. Immunofluorescence staining was positive for cardiac-specific transcription factors, GATA4, and cardiac-specific proteins, cardiac actin, cTnT, MYH6, and MLC2 in d30CMs, d60CMs, and d90CMs, and all the hiPS-CMs possessed cardiac myofibrils formation ( Fig. 1B ).
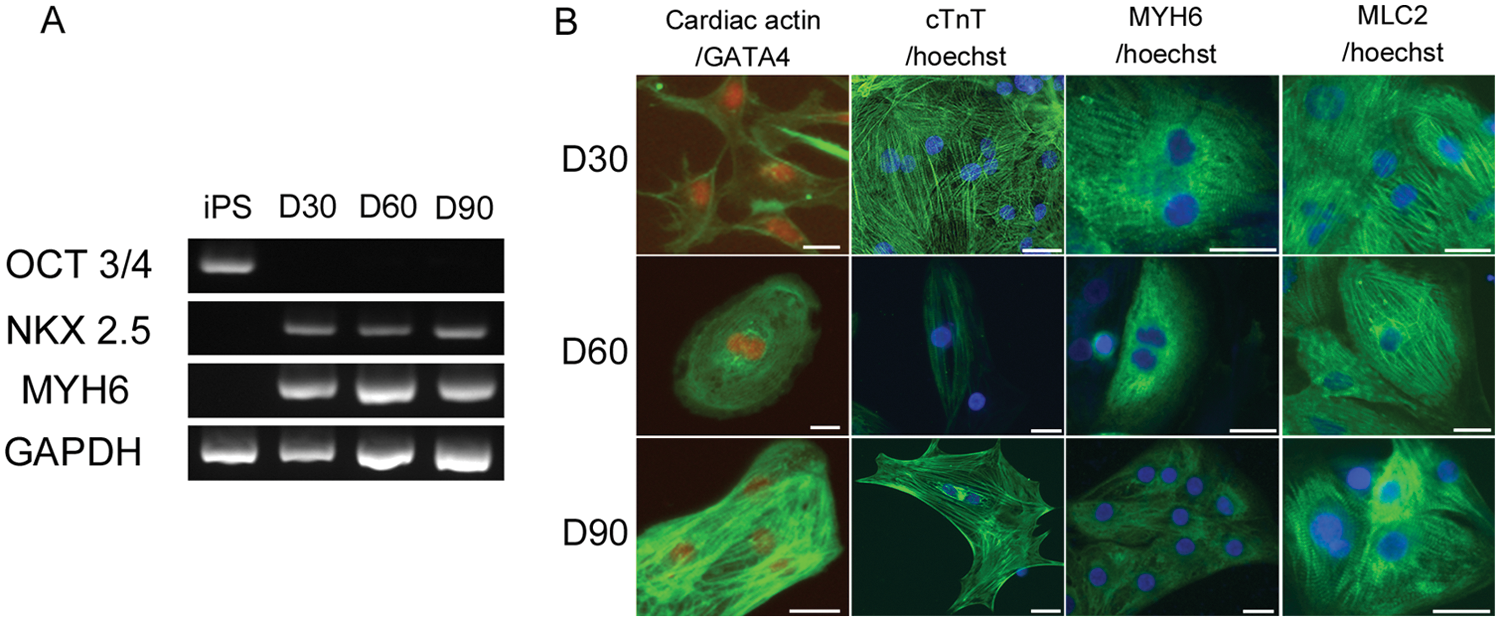
Expression of cardiac markers in the differentiated cardiomyocytes from hiPS cells. (
Characterization of Functional and Electrophysiological Properties
The beating (contraction) rate in d90CMs (35.3 ± 25.9 beats per minute [bpm]; n = 17) was significantly lower than in d30CMs (61.3 ± 20.8 bpm; n = 21) and tended to be lower than in d60CMs (46.6 ± 39.9 bpm; n = 17; Fig. 2A ). Isoproterenol (1 µM) significantly increased the beating rate in d30CMs (43.8 ± 20.4 bpm to 83.4 ± 31.6 bpm; n = 5), d60CMs (27.0 ± 9.5 bpm to 53.6 ± 15.6 bpm; n = 7), and d90CMs (31.3 ± 24.3 bpm to 72.4 ± 28.9 bpm; n = 7; Fig. 2B ). However, the beating rate was not significantly different among the three stages of differentiation of hiPS-CMs. In electrophysiological analyses, lidocaine (0–1000 µM) dose-dependently decreased the amplitude between the first positive peak and negative peak of the field potential, which reflects an action potential upstroke, in d30CMs, d60CMs, and d90CMs. A clear decrease was observed at 100 µM (d30CMs: 53.4% ± 29.2%; d60CMs: 71.3% ± 21.6%; d90CMs: 77.1% ± 19.0%, compared with vehicle [100%], respectively; Fig. 2C ). Nifedipine at 100 nM shortened the duration from the first negative peak (small arrow) to the first positive peak (large arrows) in FPD, which was consistent with the effect of calcium channel blockade on action potential duration, in d60CMs (52.3% ± 23.8%) and d90CMs (75.9% ± 17.4%) compared with vehicle (100%; Fig. 2D ). At 1000 nM nifedipine, beating was completely stopped in hiPS-CMs at all stages ( Fig. 2D ). E-4031 (0–100 µM) dose-dependently prolonged the FPD in d30CMs, d60CMs, and d90CMs compared with vehicle. The ratios of the prolongation of FPD at 100 µM were 133.4% ± 10.7%, 117.7% ± 28.0%, and 193.6% ± 94.9% in d30CMs, d60CMs, and d90CMs, respectively, compared with vehicle (100%; Fig. 2E ).
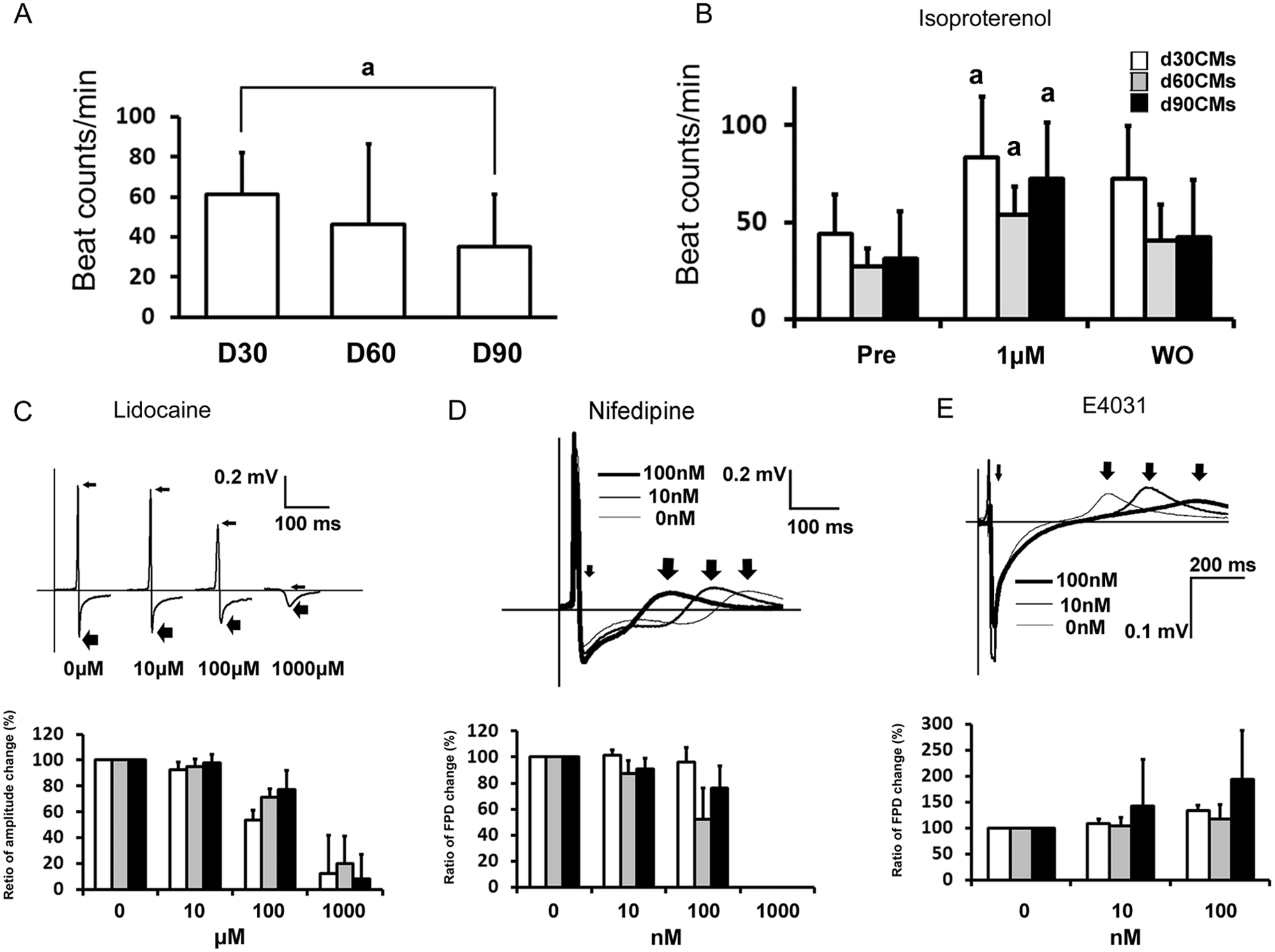
Functional properties of d30CMs, d60CMs, and d90CMs. (
Characterization of Morphological Components
In d30CMs, thin myofibrils had alignment and appeared as organized sarcomeric pattern

Ultrastructural analysis of d30CMs, d60CMs, and d90CMs. d30CMs (
Characterization of Contractile Function
Although most of the beating single cells isolated from d30CMs, d60CMs, and d90CMs with collagenase IV had a spherical shape, some beating rod-shaped cells were also noted in d60CMs and d90CMs ( Fig. 4A ). Immunofluorescence staining for GATA4 and cardiac actin was positive in the rod-shaped cells. The percentage of rod-shaped cells in d30CMs, d60CMs, and d90CMs was 0.0% ± 0.0%, 1.0% ± 1.1%, and 3.2% ± 1.7% (n = 3 each), respectively. The velocity of contraction in single cells was the fastest in the rod-shaped cells in d90CMs (7 ± 2.4 µm/s, n = 5), and then the sphere-shaped cells in d90CMs (4 ± 0.7 µm/s, n = 6), d60CMs (3 ± 2.9 µm/s, n = 5), and d30CMs (2 ± 0.8 µm/s, n = 5), in that order ( Fig. 4B , C ).
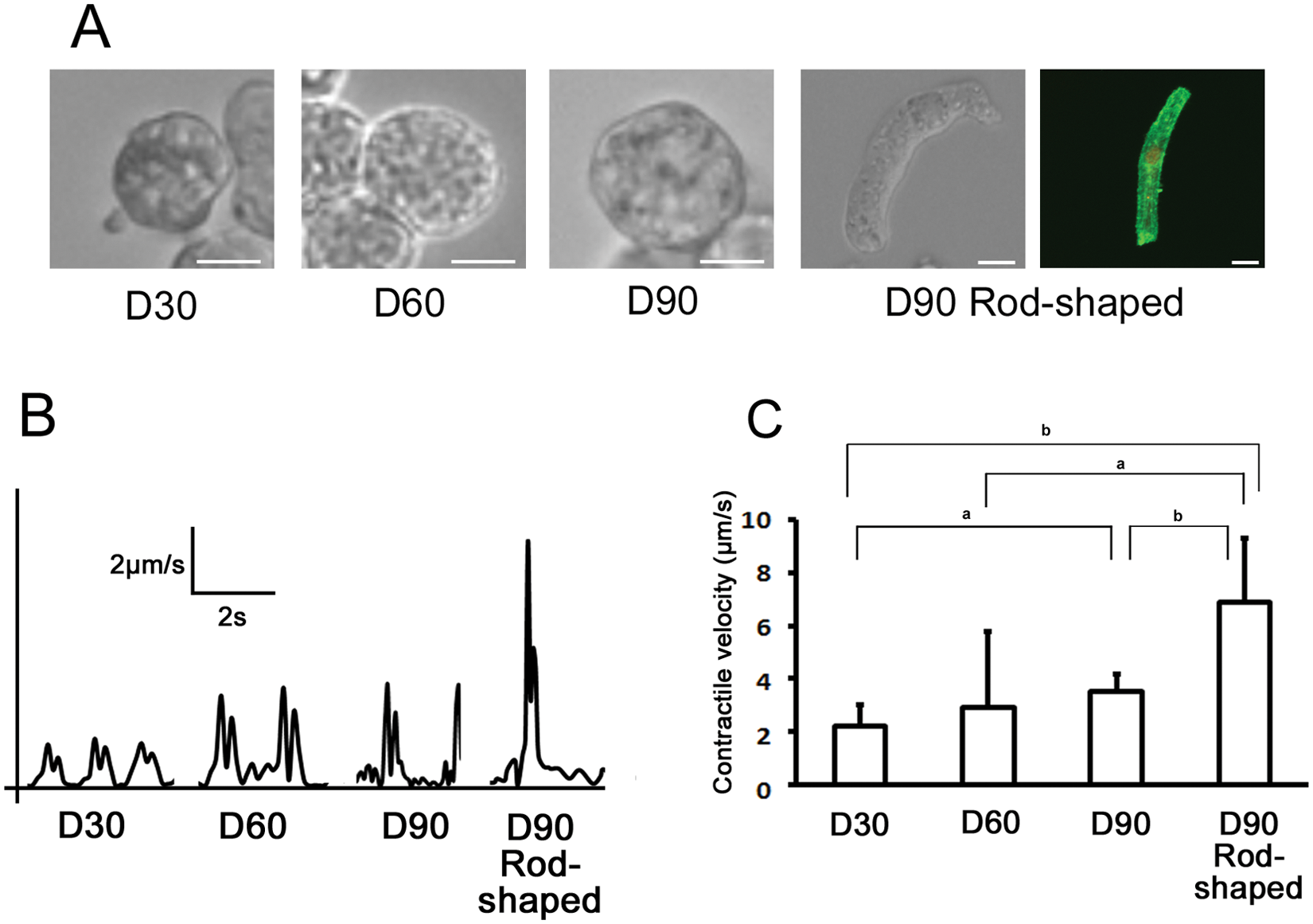
Change of contractility in human-induced pluripotent stem cell–derived cardiomyocytes (hiPS-CMs) with period of differentiation. (
Differential Gene Expression Analysis: Identification of Cardiac Enriched Genes
In global gene expression analyses, hierarchical clustering analysis by Pearson correlation showed close clustering of independent replicates reflecting high experimental reproducibility (
Fig. 5
). Furthermore, the dendrogram showed that gene expression pattern of hiPSCs and that of the differentiated cardiomyocytes or human adult left ventricle (LV) tissue were not grouped together, whereas the pattern of gene expression of hiPS-CMs at every stage (d30CMs, d60CMs, and d90CMs) was grouped with that of LV. This suggested that the gene expression profile of hiPS-CMs at each stage was more closely related to that of LV but clearly distinct from that of parental hiPSC controls. In addition, microarray analysis identified 1544, 1974, and 2862 genes in d30CMs, d60CMs, and d90CMs, respectively, that were expressed at levels up to fivefold higher or lower compared with undifferentiated hiPSCs. Among these, 670, 641, and 539 genes were upregulated and 874, 1333, and 2323 were downregulated in d30CMs, d60CMs, and d90CMs, respectively, relative to hiPSCs. The ranking of the gene ontology (GO)–based annotation related to upregulated genes in hiPS-CMs at each stage showed that the corrected p values of genes related to muscle contraction by GO accession number (GO:6936) and muscle system process (GO:3012) decreased with period of differentiation (
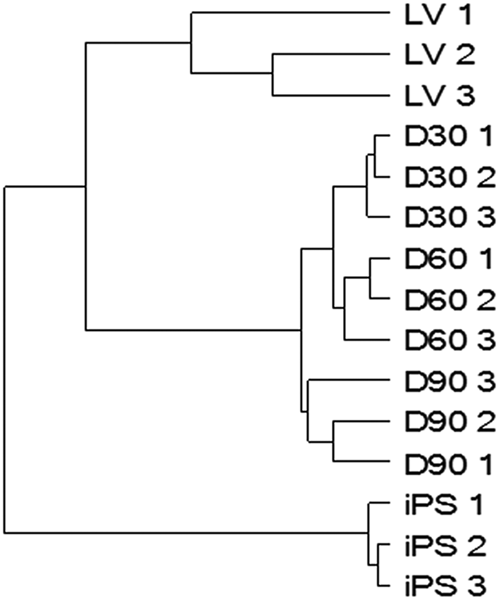
Microarray analysis and hierarchical clustering analysis. Genes expressed by human-induced pluripotent stem cell–derived cardiomyocytes (hiPS-CMs) at every stage (D30: d30CMs, D60: d60CMs, D90: d90CMs) are clustered together with those expressed by human adult left ventricle (LV) tissue and distinguished from those expressed by hiPSCs (iPS). Note the close clustering of independent biological replicates depicted in the dendrogram.
Comparison of Gene Expressions Related to Cardiac Components
Comparison of gene expressions related to gene ontology categories, namely, muscle contraction (GO:6939), adenoceptors (GO: 4935), sarcomeric proteins (GO:30017), myosin filament (GO:32982), actin filament (GO:5884), and sarcoplasmic reticulum (SR; GO:16529) in hiPS-CMs at various stages with undifferentiated hiPSCs or adult ventricle tissues are shown as heat maps in
Figure 6
. The levels of expression of many cardiac-specific genes such as troponin T2 (TNNT2), myosin heavy chain 7 (MYH7) and actin alpha cardiac (ACTC 1) were higher in hiPS-CMs in all stages than in undifferentiated hiPSCs and were close to those in the adult ventricle tissue. The levels of expression of genes related to structural constituent of muscle such as SSPN (sarcospan) and SGCA (sarcoglycan) were higher in hiPS-CMs in all stages than in undifferentiated hiPSCs and approached the levels in the adult ventricle tissue with increasing period of differentiation. The expression of genes encoding proteins related to SR, phospholamban (PLN), ryanodine receptor 2 (RYR2), sarcolipin (SLN; known as cardiac ryanodine receptor), and ATPase Ca2+ transporting cardiac muscle slow twitch 2 (ATP2A2; SERCA2) at all stages of differentiation was higher than in undifferentiated hiPSCs and was close to that in the adult ventricle tissue. The level of expression of ryanodine receptor 1 (RYR1), which is known as skeletal muscle-type ryanodine receptor, was lower (4% to 29%) in all stages of differentiation of hiPS-CMs than in undifferentiated hiPSCs and was similar to that in the adult ventricle. Although the levels of expression of calsequestrin 2 (CASQ2) and S100 calcium-binding protein A1 (S100A1) increased considerably with period of differentiation, it was still lower than in the adult ventricle tissue. The highlight and details of fold-change data in heat maps are described in
Table 1
and
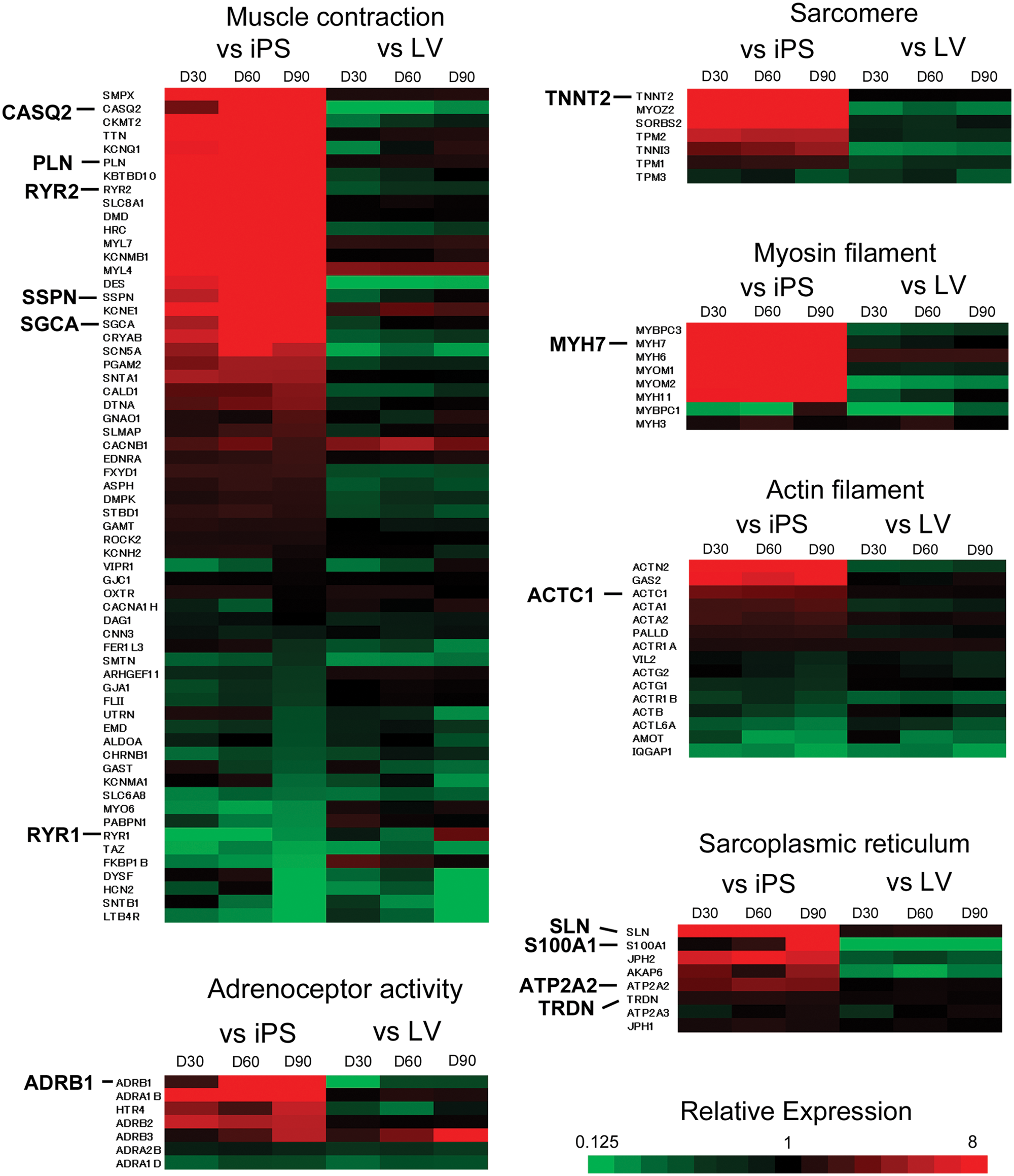
Relative gene expression levels related to muscle contraction (GO:6936), adenoceptor activity (GO:4935), sarcomere (GO:30017), myosin filament (GO:32982), actin filament (GO:5884), and sarcoplasmic reticulum (GO: 16529) in human-induced pluripotent stem cell–derived cardiomyocytes (hiPS-CMs) at each stage were compared with those in undifferentiated hiPSCs and adult ventricle. Genes with a higher expression levels in d30CMs, d60CMs, or d90CMs compared with undifferentiated hiPSCs or the adult ventricle tissue are indicated in red, whereas those with a lower expression level are shown in green. iPS, undifferentiated hiPSCs; LV, adult left ventricle; D30, d30CMs; D60, d60CMs; D90, d90CMs.
Comparison of Gene Expressions in hiPS-CMs at Various Stages with Undifferentiated hiPSCs or Adult Ventricle Tissues
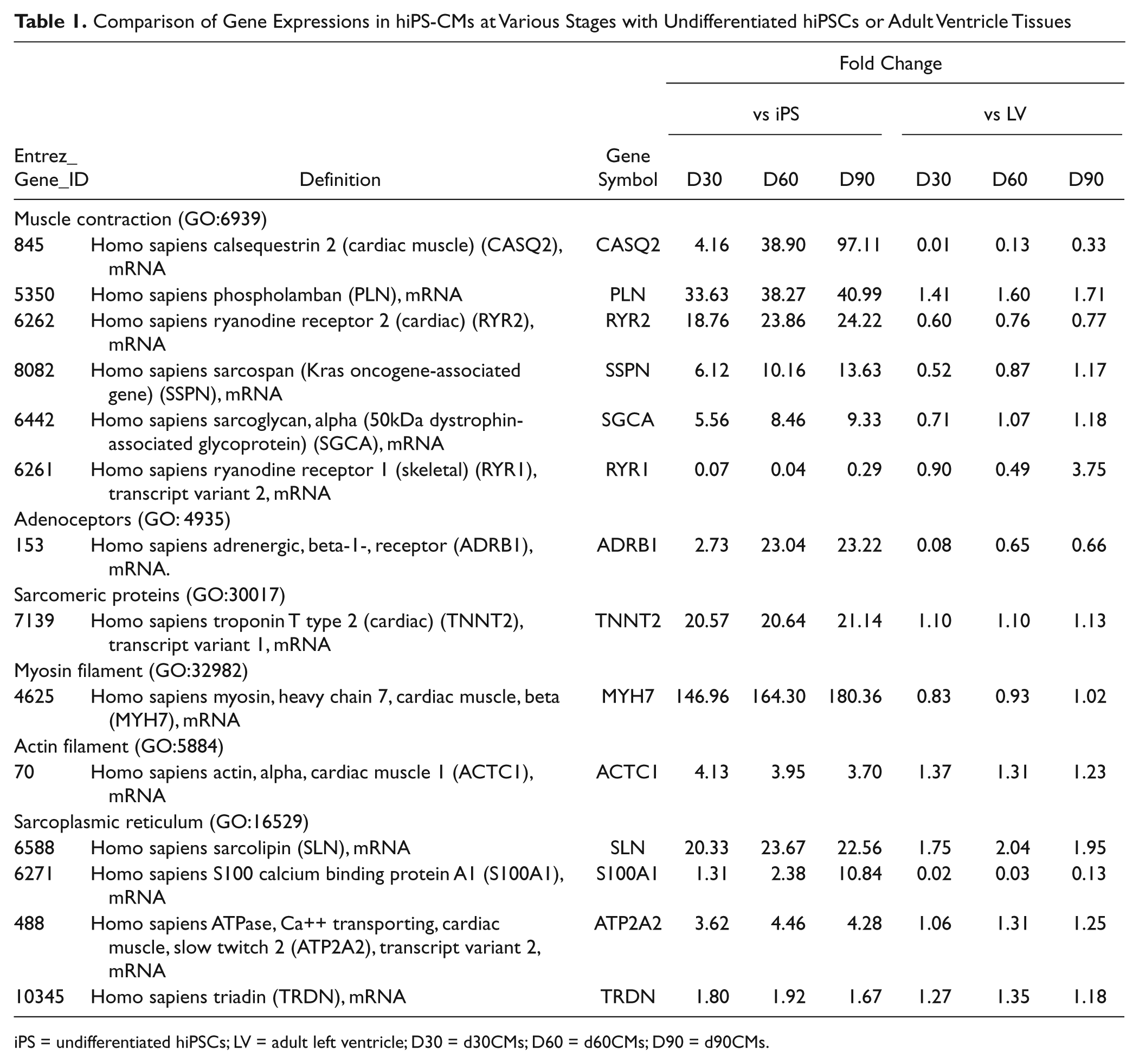
iPS = undifferentiated hiPSCs; LV = adult left ventricle; D30 = d30CMs; D60 = d60CMs; D90 = d90CMs.
Discussion
In the present study, to more fully understand the characteristics of hiPS-CMs at various stages of differentiation in culture and to determine the appropriate differentiation stage for assessment of pharmacological effects or toxicity of drug candidates, we characterized the functional and morphological components and global gene expression in hiPS-CMs at different stages of differentiation and documented the similarities and differences among them. This is the first report, to our knowledge, that has evaluated the characteristics of hiPS-CMs at day 60 or at later stages of differentiation and compared these and the corresponding global gene expression with those of the adult heart. The expression of cardiac-specific genes and proteins in hiPS-CMs differentiated in culture for 20 to 30, 50 to 60, and 80 to 90 d that were spontaneously contracting was confirmed by RT-PCR and immunohistological staining. The GO-based annotation analyses of upregulated genes showed that the muscle system process and muscle contraction were among the top three categories and that the corrected p values for these annotations decreased with period of differentiation. Therefore, the essential cardiac characteristics such as spontaneous beating and those related to heart development were already developed at day 30 of differentiation, and gene expression in hiPS-CMs related to muscle function was strongly associated with period of differentiation.
We found that spontaneous beating rate in hiPS-CMs decreased with the developmental day. The spontaneous beating activity of ventricular-like hESC-derived embryoid bodies cultured for 40 to 95 d tends to decrease with time of culture. 26 Therefore, this suggested that hiPS-CMs differentiated into ventricular-like myocytes with the developmental day in this culture system. Beating rates at all stages increased by exposure to isoproterenol. In global gene expression analyses, the gene expression level of adrenergic β-1 receptor (ADRB1) in hiPS-CMs at various stages was higher than that in undifferentiated hiPSCs. These results showed that functions related to the β-adrenergic pathway were already activated at day 30 of differentiation in vitro. In electrophysiological analyses, lidocaine, a sodium channel blocker, dose dependently shortened the amplitude between the first positive peak and negative peak of the FPD at all stages. E4031, an hERG-type potassium channel blocker, caused dose-dependent prolongation of the FPD in hiPS-CMs at each stage of differentiation. These results are consistent with a previous report using another sodium channel blocker, quinidine, and E4031in hiPS-CMs at days 14 to 15 of differentiation. 12 Nifedipine, a calcium channel blocker, shortened FPD in d60CMs and d90CMs at 100 nM. Although FPD did not change in d30CMs at this concentration in the present study, beating completely stopped at 1000 nM nifedipine in the hiPS-CMs not only in d60CMs and d90CMs but also in d30CMs. In the previous report, nifedipine at 1000 nM stopped contraction of hiPS-CMs at day 60. 27 Several reports have demonstrated that hiPS-CMs at day 20 to 40 of differentiation have electrophysiological properties and respond to ion channel blockers.12–14 In the present study, electrophysiological responses to ion channel blockers in d30CMs were similar to those in d60CMs and d90CMs, although the sensitivity to the calcium channel blocker was weaker. These functional assays showed that functions related to the β-adrenergic pathway and electrophysiological properties in hiPS-CMs did not change markedly until day 90 of differentiation. In this respect, d30CMs are useful for basic screening of these functions because of the short time needed to prepare the cultures.
Ultrastructural analyses showed that the initial irregular myofibrillar distribution in d30CMs gradually organized into parallel myofibril arrays and ultimately resulted in the generation of well-defined sarcomeres in d60CMs. These morphological properties did not change noticeably between d60CMs and d90CMs. Recently, we demonstrated that doxorubicin, a cardiotoxic drug, induced myofilament disruption in cardiomyocytes isolated from d60CMs. 18 This suggested that d60CMs are useful for assessment of cardiotoxic drugs that induce morphological changes in cardiomyocytes. Intriguingly, although the morphological components in d30CMs were immature compared with those in d60CMs and d90CMs, gene expression levels of muscle-related proteins such as ACTC1, TNNT2, and MYH7 in d30CMs were higher than those in undifferentiated hiPSCs and similar to those in adult ventricle tissue. In addition, expression of genes related to structural constituent of muscle such as SSPN and SGCA, which are dystrophin-associated glycoproteins, increased with period of differentiation and approached those of adult ventricle tissue. These results suggest that molecules related to the structural constituent are essential for morphological maturation in hiPS-CMs, and the expression of their related genes might be a good marker of morphological maturation.
Motion analysis showed that the velocity of contraction, an index of cardiac contractile function, was higher in sphere-shaped cells isolated from d90CMs than those from d30CMs. Expression of genes related to SR, which releases and stores calcium ions for contractile function, such as PLN, RYR2, SLN, TRDN (triadin), and ATP2A2 (SERCA2) was higher in hiPS-CMs at every stage than those in undifferentiated hiPSCs and close to those in adult ventricle tissue. On the other hand, expression levels of CASQ2, which associates with RYR2 and regulates calcium ion release from SR, 28 and S100A1, which is an EF-hand Ca2+ sensor protein and is related to Ca2+ homeostasis in the contraction process, 29 gradually increased with period of differentiation. Therefore, the change in gene expression levels of CASQ2 and S100A1 might be closely related to the development of cardiac contractile function. These results suggest that contractile function in d90CMs is the most mature among the stages examined in this study. However, to investigate the detailed contractile function, hiPS-CMs with better developed SR function are needed because the expression levels of some genes related to SR in d90CMs are still lower than in adult ventricle tissue. Interestingly, we found a few rod-shaped cardiomyocytes that were positively stained for GATA4 and cardiac actin in the cells isolated from d60CMs and d90CMs, and their numbers were higher in d90CMs than in d60CMs. The velocity of contraction in rod-shaped cells isolated from d90CMs was much higher than that of sphere-shaped cells in hiPS-CMs at every stage. The cell length in hES-CMs at day 35 of differentiation was longer compared with those at day 10 to 21 and day 21 to 35 of differentiation, and this was coupled with a progressive ultrastructural development from an irregular myofibrillar distribution to an organized sarcomeric pattern. 16 Therefore, the rod-shaped cells might have more mature contractility associated with morphological components than sphere-shaped cells. Further investigation to improve the effective production of rod-shaped cells in hiPS-CMs might help understand the maturation process leading to adult cardiomyocytes.
In conclusion, d30CMs may be useful for basic screening of pharmacological effects on functions related to the β-adrenergic pathway or electrophysiological properties, and d60CMs and d90CMs may be appropriate for toxicological assessment of drugs on the morphology of myofibrils. For the assay of the effects of drugs on contractile function associated with subcellular components such as SR, d90CMs appear to be most suitable among the stages examined in this study. Further improvements in inducing the maturation of hiPS-CMs by advances in culture systems, the discovery of maturation-promoting factors, or the development of new equipment to assist separation of mature cardiomyocytes might aid development of improved model systems that possess adult heartlike phenotypes for drug screening or drug safety risk assessment and, moreover, might contribute to future use of these cells in regenerative medicine.
Footnotes
Acknowledgements
Human iPS cell lines, 253G1, were kindly provided by Kyoto University under a material transfer agreement. We thank Dr. Hirofumi Nagai for supportive encouragement; Drs. Nobuyuki Nishida and Makoto Miyamoto of Drug Safety Research Laboratories, Takeda Pharmaceutical Company Limited, and Eimei Sato of Laboratory of Animal Reproduction, Graduate School of Agricultural Science, Tohoku University, for their helpful suggestions; and Shingo Ookubo, Ryoutaro Hori, Maya Morita, and Yoshiko Okai of Drug Safety Research Laboratories, Takeda Pharmaceutical Company Limited, for their excellent technical assistance.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.