
Abstract
Cardiomyocytes derived from embryonic stem cells (ES-CMs) and induced pluripotent stem cells (iPS-CMs) are useful for toxicity and pharmacology screening. In the present study, we found that cardiomyocyte-rich beating cell clusters (CCs) emerged from murine embryonic stem cell (mESC)–derived beating EBs and from human-induced pluripotent stem cell (hiPSC)–derived beating EBs dissociated by gentle pipetting with a thin glass pipette. The percentage of cardiac troponin T (cTnT)–positive cells in the beating CCs obtained from mESC-derived and hiPSC-derived beating EBs was higher (81.5% and 91.6%, respectively) than in beating-undissociated EBs (13.7% and 67.1%, respectively). For mESCs, the yield of cTnT-positive cells from beating CCs was estimated to be 1.6 times higher than that of beating EBs. The bromodeoxyuridine labeling index of mouse ES-CMs and human iPS-CMs in beating CCs was 1.5- and 3.2-fold, respectively, greater than those in beating EBs. To investigate the utility of the cells in toxicity assessment, we showed that doxorubicin, a cardiotoxic drug, induced myofilament disruption in cardiomyocytes isolated by this method. This simple method enables preparation of mouse ES-CMs and human iPS-CMs with better proliferative activity than beating EBs not dissociated by pipetting, and the cardiomyocytes are useful for drug-induced myocardial toxicity testing.
Introduction
For drug discovery and development, genetically modified rodents, immortalized human cell lines, and animal models have provided useful information in studying safety and efficacy of drug candidates. However, most lead candidates that pass various in vitro screening tests fail to be developed due to safety and/or efficacy issues in nonclinical and/or clinical studies. Therefore, additional new drug screening model systems are needed to more closely mimic human biochemistry and physiology1,2 and to improve the success rate.
As with embryonic stem cells (ESCs), induced pluripotent stem cells (iPSCs) have the abilities of self-renewal and differentiation into various cell types in vitro.3,4 Hence, we may be able to produce various kinds of cells possessing the same genetic background from ESCs or iPSCs, and such cells derived from human sources are expected to be invaluable for establishment of in vitro models that closely mimic physiological functions of humans. ESC- and iPSC-derived cardiomyocytes (ES-CMs and iPS-CMs, respectively) have received much attention because of their potential for regenerative medicine and drug screening. 2 For example, in drug safety screening, electrophysiological assays, including hERG inhibition and QT prolongation using human ES-CMs or iPS-CMs, might be more predictive of clinical outcomes, such as arrhythmia.5,6 In addition, previous studies have shown that the morphological characteristics related to the contractile function in mouse and human ES-CMs are similar to cardiomyocytes in the adult heart.7,8
To employ ES-CMs and iPS-CMs in cardiac toxicity assessment, it is necessary to isolate these cells from beating (i.e., spontaneously contracting) embryoid bodies (EBs) or beating cell colonies since they usually contain several other types of cells besides cardiomyocytes resulting from the differentiation process, because the contaminating cells make the assessment of the cardiac toxicity difficult. Although manual dissection using laser capture microscopy 9 or genetic manipulation of ES/iPS cells has been used in many studies for isolation of these cells, these methods are time-consuming and/or require complex techniques with expensive equipment.
In the present study, when murine embryonic stem cell (mESC)–derived beating EBs were dissociated by gentle pipetting with a thin glass pipette, we found that cardiomyocyte-rich beating cell clusters (CCs) emerged from the particles of dissociated beating EBs. On the basis of this observation, we established a simple method for purification of cardiomyocytes. The method was also applicable to purification of cardiomyocyte-rich beating CCs from human-induced pluripotent stem cell (hiPSC)–derived beating EBs. In addition, we tested the effect of a well-known cardiotoxic drug, which induces morphological changes on cardiomyocytes in vivo, on the cardiomyocytes isolated from mESC- and hiPSC-derived beating CCs to evaluate the utility of these cells for use in morphological toxicity assessments.
Materials and Methods
mESC Culture, Differentiation, and Isolation of Cardiomyocyte-Rich Cell Clusters
A sub–cell line of the C57BL/6J mESCs originally established in the Biology Research Laboratories, Pharmaceutical Research Division, Takeda Pharmaceutical Company Limited, Osaka, Japan, was used in this study. Differentiation of mESCs was induced using a modification of the method described previously. 7 Briefly, mESCs were cultured on a feeder layer of mitomycin C–treated mouse embryonic fibroblasts (MEFs) in Dulbecco’s modified Eagle’s medium (DMEM; Sigma, St. Louis, MO) supplemented with 10% fetal calf serum (FCS; Chemicon, Billerica, MA), nonessential amino acids (Gibco, Carlsbad, CA), 2-mercaptoethanol (Chemicon), EmbryoMax ES Cell Qualified Nucleosides (Chemicon), and 2000 U/mL leukemia inhibitory factor (LIF; ESGRO: Chemicon), in a 5% CO2/95% air environment. The mESCs were then trypsinized and cultured in low cell binding 96-well plates (Nunc, Tokyo, Japan) until they formed EBs. The culture medium was the same as described above except for changing the concentration of FCS from 10% to 30% and minus the LIF. mESCs (1 × 104) were transferred into 100 µL of medium in each well of the plate ( Fig. 1A ). On days 5, 6, or 7 of differentiation, beating EBs were dissociated by gentle pipetting two to three times with a glass pipette of 200 to 300 µm internal diameter in the wells. On the next day of culture after the dissociation, small beating CCs isolated from beating EBs were picked up using a glass pipette. In the present study, beating CCs dissociated on day 7 were cultured until day 10 of differentiation and used to investigate the characteristics of the beating CCs by the following analyses.
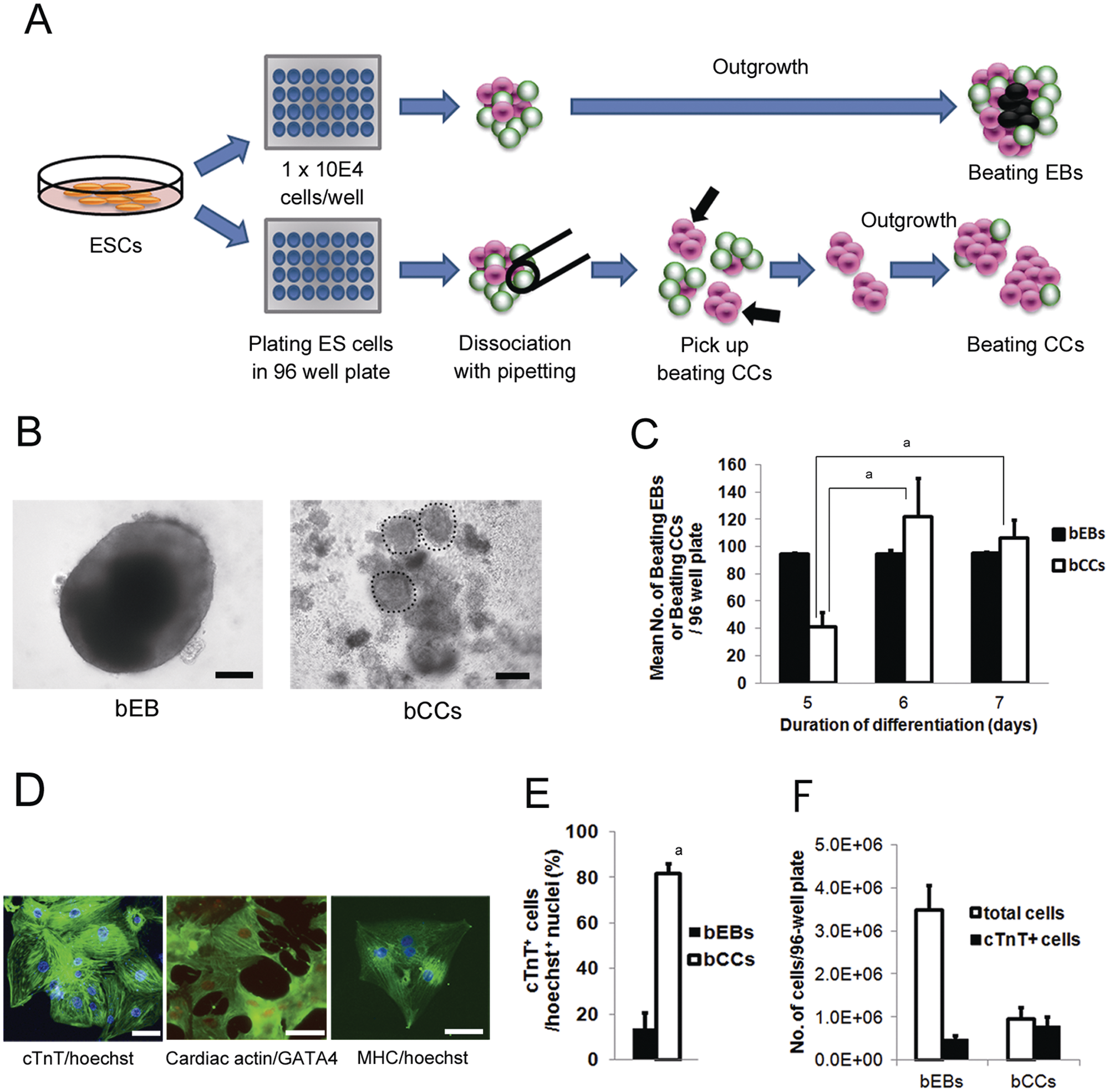
Purification of cardiomyocytes from mouse embryonic stem cells (mESCs). (
hiPSC Culture, Differentiation, and Isolation of Cardiomyocyte-Rich Cell Clusters
The hiPSC line (253G1) from Kyoto University was maintained as previously described.3,16 Undifferentiated hiPSCs were maintained on a feeder layer of mitomycin C–treated MEF in Primate ES medium (ReproCELL, Yokohama, Japan) supplemented with 4 ng/mL recombinant human basic fibroblast growth factor (bFGF; PeproTech, Rocky Hill, NJ) in a 5% CO2/95% air environment at 37 °C. Differentiation of hiPSCs was induced as previously described with slight modifications. 17 Briefly, the colonies of hiPSCs were isolated in dissociation solution (ReproCELL), and they formed EBs in suspension culture in low-cell-binding 90-mm dishes (Nunc) in basic medium (StemPro34; Invitrogen, Carlsbad, CA), containing 2 mM glutamine, 4 × 10 M monothioglycerol (MTG), and 50 µg/mL ascorbic acid (Sigma). The following cytokines were also used according to the differentiation stage: days 0 to 1, bone morphogenetic protein 4 (BMP4; 0.5 ng/mL); days 1 to 4, BMP4 (10 ng/mL), bFGF (5 ng/mL), and activin A (3 ng/mL); days 4 to 8, dickkopf homolog 1 (DKK1; 150 ng/mL) and vascular endothelial growth factor (VEGF; 10 ng/mL); and after day 8, VEGF (10 ng/mL), DKK1 (150 ng/mL), and bFGF (5 ng/mL) in the basic medium described above. All factors were purchased from R&D Systems (Minneapolis, MN). Cultures for the differentiation were maintained in a 5% CO2/95% air environment at 37 °C. On day 14 of differentiation, beating EBs were dissociated with the same pipetting method used for mESC-derived beating EBs. The day after dissociation, beating CCs isolated from beating EBs were picked up and cultured until day 20 of differentiation.
Fluorescent Immunostaining for Cardiac Markers
Several beating EBs and beating CCs derived from mESCs (day 10 of differentiation) and from hiPSCs (day 20 of differentiation) were picked up and dissociated using 10 mg/mL collagenase IV for 3 to 4 h at 37 °C. The dissociated cells were seeded into collagen I–coated dishes. Two or 3 days after seeding the dishes, the cells were fixed in 4% formaldehyde for 30 min and subsequently permeabilized with 2.5% Tween-20 (WAKO, Osaka, Japan) at room temperature. The cells were incubated at 4 °C overnight with the following primary antibodies diluted in Dulbecco’s PBS(–) buffer (PBS; Gibco): monoclonal anti–cardiac troponin T (cTnT, 1:100 dilution; Abcam, Tokyo, Japan), monoclonal anti–myosin cardiac heavy chain (MHC, 1:100 dilution; Abcam), polyclonal anti-GATA4 (1:100 dilution; Abcam), monoclonal anti–cardiac actin (1:50 dilution; Fitzgerald, Acton, MA), and polyclonal anti–myosin light chain 2 (MLC2, 1:100 dilution; Abcam). Fluorescent dye–conjugated secondary antibodies, Texas red–conjugated goat anti-rabbit IgG (1:100; Vector Laboratories, Burlingame, CA), FITC-conjugated rabbit anti-mouse IgG (1:40; Dako, Glostrup, Denmark), or FITC-conjugated swine anti-rabbit IgG (1:40; Dako) was applied to the cells for 1 h at room temperature. The nuclei were stained with 5 µg/mL Hoechst 33342 (Sigma) for 10 min at room temperature. The specimens were then observed under a Biozero fluorescence microscope BZ-9000 (Keyence, Osaka, Japan). For quantitative analysis, fluorescence images were acquired with an IN Cell Analyzer 1000 (GE Healthcare, Tokyo, Japan) to calculate ratios of mouse ES-CMs (mES-CMs) or human iPS-CMs (hiPS-CMs) to the total cells in beating EBs and beating CCs, respectively. Then, the number of cTnT-positive cells per the total number of nuclei was measured with an IN Cell Analyzer 1000 Workstation 3.4 software (GE Healthcare).
Analysis of mESC/hiPSC-Derived Cardiomyocyte Proliferation
Several beating EBs and beating CCs derived from mESCs on day 10 of differentiation or from hiPSCs on day 20 of differentiation were picked up and dissociated using the method described above. The dissociated cells were cultured as a monolayer on a gelatin-coated surface for 2 or 3 days in differentiation medium, then stained with a 5-bromo-20-deoxyuridine (BrdU) Labeling and Detection Kit I (Roche Diagnostics, Tokyo, Japan) according to the manufacturer’s protocol with some modifications. Briefly, BrdU was added to the plates for 1 h. After fixation in 100% methanol for 20 min at −20 °C, the dissociated cells were incubated for 30 min with a 1:10 dilution of anti-BrdU antibody at 37 °C. Then, the cells were incubated for 30 min with a 1:500 dilution of Texas red–conjugated goat anti-mouse IgG (Abcam) to label the BrdU-positive nuclei. To double-stain with cardiac marker, the cells were then incubated with a 1:100 dilution of mouse monoclonal anti-cTnT (Abcam) overnight. FITC-conjugated rabbit anti-mouse IgG (1:40; Dako) was used to detect cTnT-positive cells as described above. Nuclei were stained with 5 µg/mL Hoechst 33342 (Sigma). BrdU-positive cells were examined under a Biozero fluorescence microscope BZ-9000 (Keyence). The BrdU-positive nuclei were counted relative to the total number of nuclei (Hoechst 33342) in mESC- or hiPSC-derived cTnT-positive cells. At least three dishes were analyzed for each condition, and a total of more than 500 myocytes were counted in each experiment.
Induction of Myofibrillar Disarray by Doxorubicin
The assessment of the myofibrillar disarray in mES-CMs and hiPS-CMs induced by doxorubicin was performed as described previously with slight modifications. 18 mES-CMs and hiPS-CMs were isolated from beating CCs on days 19 to 21 and days 50 to 60 of differentiation, respectively, by dissociation using 10 mg/mL collagenase IV for 3 to 4 h at 37 °C and then cultured as a monolayer on gelatin-coated culture dishes for 2 days in differentiation medium. These cells were then treated with 0.5 µM doxorubicin (WAKO) for 24 h and then fixed and stained for cTnT and nuclei as described above. The stained preparations were observed under a Biozero fluorescence microscope BZ-9000 (Keyence).
Statistical Analysis
Statistical analysis was performed with the Student t test and the Welch t test for comparison of two groups with equal variances and unequal variances, respectively. Values of p ≤ 0.05 on both tests were considered statistically significant. All statistical calculations were carried using the SAS system (SAS Institute, Cary, NC).
Results
Differentiation and Purification of Mouse ESC-Derived Cardiomyocytes
Beating EBs emerged after day 5 of differentiation, and 90.6% of all EBs at day 7 were spontaneously beating. When beating EBs were dissociated by gentle pipetting, small beating CCs, which had obvious large beating regions, emerged from the particles of dissociated beating EBs ( Fig. 1A : arrows; Fig. 1B : right panel). The mean number of beating CCs dissociated on day 5 of differentiation was 41 in a 96-well plate, and the number was higher on days 6 and 7 of differentiation (122 and 106, respectively). The mean number of beating EBs in a 96-well plate was similar for days 5, 6, and 7 of differentiation ( Fig. 1C ).
We performed immunofluorescence staining to determine whether the cells in beating CCs expressed cardiac marker proteins ( Fig. 1D ). The cells isolated from beating CCs on day 10 of differentiation displayed sarcomere formation based on positive staining with antibodies directed against cTnT, cardiac actin, and MHC, as well as nuclear expression of the cardiac-specific transcription factor GATA4, which is a typical feature of cardiomyocytes. The percentage of cTnT-positive cells in the beating CCs was 81.5% and was much higher than that in the beating EBs (13.7%) ( Fig. 1E ). On day 10, cells dissociated with 10 mg/mL collagenase IV from beating EBs or beating CCs were stained with trypan blue to count the number of dead cells. In a 96-well plate, the total number of live cells in all the beating EBs and beating CCs were 3.5 × 106 and 9.6 × 105, respectively ( Fig. 1F ). The total number of cTnT-positive cells in beating EBs and beating CCs in a 96-well plate was estimated to be 4.8 × 105 (3.5 × 10× 0.137) and 7.8 × 105 (9.6 × 10× 0.815), respectively. Therefore, the yield of cTnT-positive cells from beating CCs in a plate was estimated to be 1.6 times higher than that of beating EBs ( Fig. 1F ).
Proliferative Activity of Cardiomyocytes Isolated from mESC-Derived Beating EBs and Beating CCs
Immunofluorescence analysis on day 10 of differentiation showed that fractions of the cTnT-positive cells from mESC-derived beating EBs and beating CCs contained BrdU-positive nuclei, indicating the presence of proliferating cardiomyocytes ( Fig. 2A ). In beating EBs, 15.5% of cTnT-positive cardiomyocytes were BrdU positive, whereas in beating CCs, 22.6% of cTnT-positive cells were BrdU positive ( Fig. 2B ).
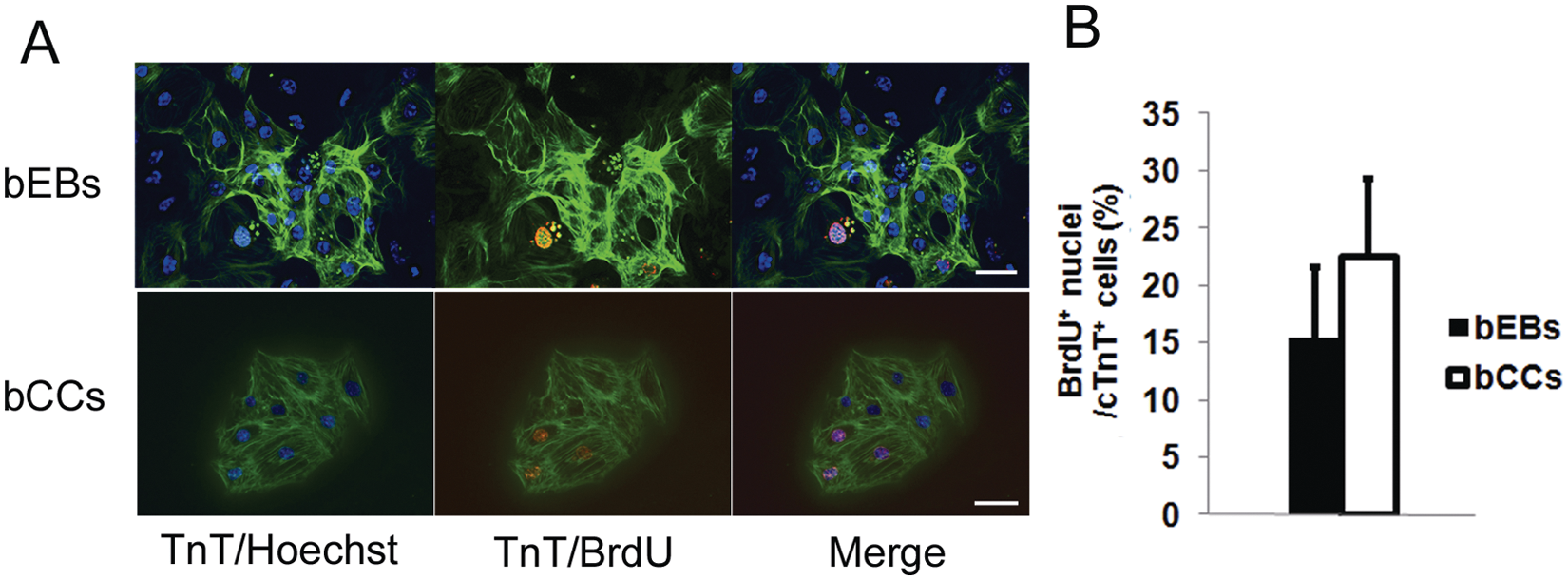
Proliferative activity of cardiomyocytes in mouse embryonic stem cell (mESC)–derived beating embryoid bodies (bEBs) or beating cell clusters (bCCs) on day 10 of differentiation. (
Differentiation, Purification, and Proliferative Activity of Human iPSC-Derived Cardiomyocytes
Beating EBs were observed after 12 to 14 days of differentiation. Dissociation of a beating EB with a thin glass pipette on day14 of differentiation generated several beating CCs, and this was similar to that seen with mESC-derived beating EBs ( Fig. 3A ). The immunofluorescence staining for cTnT, MHC, and MLC2 showed that the cells in beating CCs possessed cardiac myofibrils ( Fig. 3B ). The percentage of cTnT-positive cells in the beating CCs was 91.6% and significantly higher than that in beating EBs (67.1%) ( Fig. 3C ). As for the proliferative activity, a fraction of the cTnT-positive cells in the beating EBs and beating CCs contained BrdU-positive nuclei on day 20 of differentiation ( Fig. 3D ). The percentage of BrdU-positive cardiomyocytes was 3.3% in beating EBs and significantly higher in beating CCs at 10.4% ( Fig. 3E ).
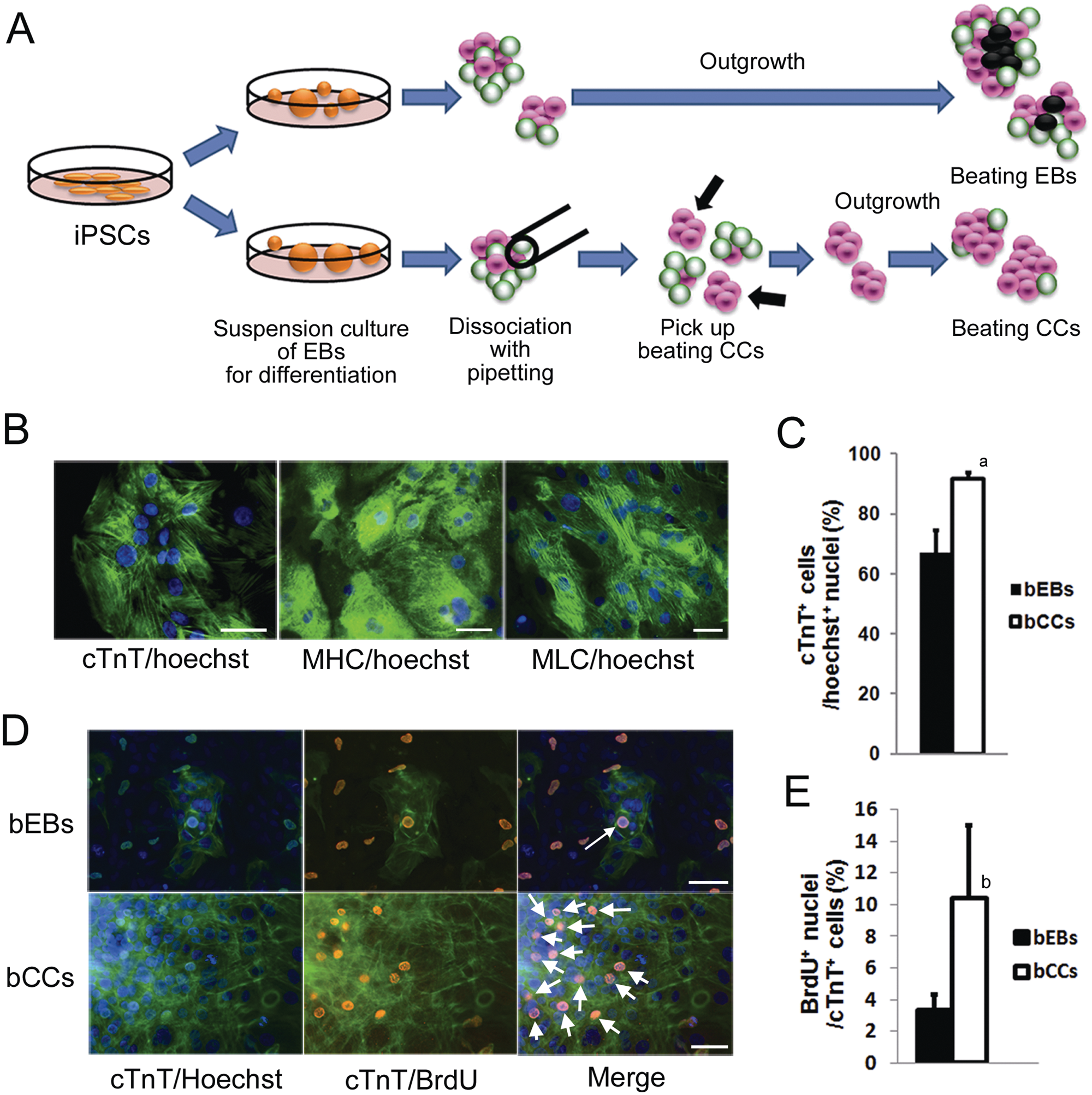
Purification of cardiomyocytes from human-induced pluripotent stem cells (hiPSCs). (
Doxorubicin Induces Myofilament Disarray in mESC- and hiPSC-Derived Cardiomyocytes
In the control mES-CMs, the cTnT immunofluorescence was localized in cardiac filaments along the long axes of cells ( Fig. 4A ). After the treatment of mES-CMs with 0.5 µM doxorubicin for 24 h, the cells became round in shape, and the myofilaments showed a “moth-eaten” disorganized appearance with a relative paucity of cTnT staining ( Fig. 4B ). In the hiPS-CMs, cTnT-positive cardiac filaments were also observed along the long axes of the nontreated cells ( Fig. 4C ), and treatment with doxorubicin caused structural changes of myofibrillar disarray ( Fig. 4D ). These changes were quite similar to those in the mES-CMs.
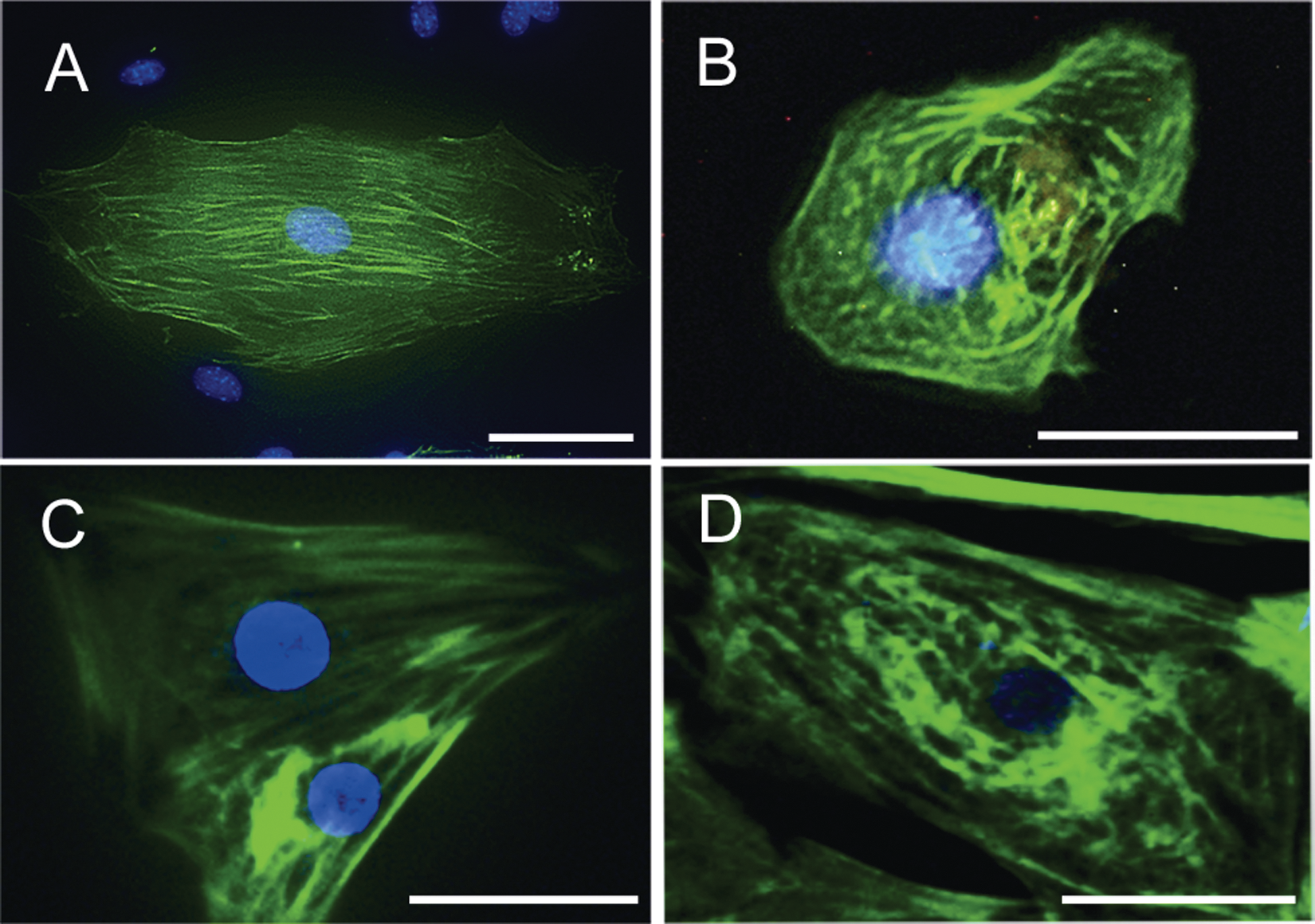
Effect of doxorubicin on morphology of cardiomyocytes derived from mouse embryonic stem cells (mESCs) and human-induced pluripotent stem cells (hiPSCs) on days 10 and 20 of differentiation, respectively. (
Discussion
In the previous studies, manual dissection of ES/iPS-derived beating foci or genetic manipulation with various combinations of cardiomyocy-tespecific reporters was used to obtain highly purified ES/iPS-CMs,9-15 and the purity of ES/iPS-CMs isolated using genetic manipulation techniques was reported to be 83% to 99% and was almost equivalent to that obtained in this study. However, these methods require genetic modification of the cells or expensive equipment and are technically complex. In addition, the proliferative activity was not shown in these reports, and the purification method that uses a fluorescence-activated cell sorter to isolate cardiomyocytes might result in damage to the sorted cells due to laser emission and the process of single-cell sorting. In contrast, the purification method established in the present study does not need any especial equipment, causes no obvious damage to cells, and is much simpler for the isolation of mES- or hiPS-CM-rich beating CCs from beating EBs. Although the size of mES/hiPS-CM-rich beating CCs was smaller than beating EBs, the yield of mES-CMs isolated from beating CCs was 1.6 times higher than that from beating EBs in a 96-well plate. Intriguingly, the proliferative activities of cardiomyocytes in beating CCs versus the corresponding undissociated EBs were higher by 1.5 times (22.6% vs. 15.5%). On the other hand, the yield of hiPS-CMs from a beating EB could not be calculated since colonies of hiPSCs did not dissociate into single cells in suspension culture to form EBs in the present differentiation method, and therefore, the number of hiPSCs could not be accurately determined in each 90-mm dish. However, we found that the proliferative activities of cardiomyocytes in beating CCs versus the corresponding undissociated EBs were also higher by 3.2 times (10.4% vs. 3.3%) for hiPS-CMs. Therefore, the pipetting purification method appears to improve the proliferative activity of mES/hiPS-CMs and might therefore increase the eventual yield of mES-CMs.
Although the mechanism of emerging mES/hiPS-CM-rich beating CCs from beating EBs by the purification method is unclear, one of the reasons may be that clusters of cardiac linage cells are included in EBs. In our previous study, we demonstrated that clusters of cardiac actin–positive cells were localized in frozen sections of beating EBs produced from 1 × 104 mES cells on day 7. 7 This suggests that the dissociation by pipetting might result in the separation of these clusters of cardiac linage cells from EBs. In addition, the size of beating CCs might also be an important factor for generation of a high density of cardiomyocytes in them. It has been reported that mESC-derived EBs, whose diameter was 100 to 200 µm on day 3 of differentiation, may mimic the early stages of development in vivo, and after extended differentiation, many of the mEBs develop beating outgrowths. 19 Mohr et al. 20 reported that the percentage of cardiomyocytes in small hESC-derived beating EBs whose diameters were 88 µm was higher than that of large hESC-derived beating EBs whose diameters were 350 µm. In addition, many cells located inside the large EBs are found to be apoptotic, indicating that most of those ESCs could not differentiate into healthy cells.20,21 In the present study, the diameter of beating EBs was about 500 µm, and that of the beating CCs dissociated with pipetting was about 100 µm. In addition, the number of dead cells stained with trypan blue was larger in beating EBs than in beating CCs on day 10 of differentiation (data not shown). This suggests that the size of beating CCs is better suited for cardiogenesis, and the dissociation of beating EBs might prevent mES-CMs or hiPS-CMs from cell death by enabling more efficient exchanges of O2 and CO2 and nutrients and waste.
The number of beating CCs obtained from mESC-derived beating EBs at day 6 or 7 was significantly higher than that at day 5. Miller et al. 22 demonstrated the onset of robust Nkx2-5 expression in the cardiac progenitor cell population on day 6 of culture after the differentiation from mESCs. Therefore, it might be important to dissociate beating EBs at the time of the emergence of the cardiac progenitor cell population, and the dissociation might improve the conditions for the growth of a cardiac progenitor cell population in beating EBs.
Although previous reports have shown the applicability of ES-CMs and iPS-CMs to the assessment of drug-induced cardiac toxicity by electrophysiological methods,6,23,24 there are few reports describing drug-induced morphological toxic changes. In the present study, we tested the effect of a cardiotoxic drug, doxorubicin, on the cardiomyocytes isolated from mESC- and hiPSC-derived beating CCs to evaluate their utilities for toxicity assessment. We found that doxorubicin-induced morphological changes such as myofibril disarray in both mES-CMs and hiPS-CMs. This is the first report to our knowledge that has demonstrated such an effect. These observations are similar to the myofibrillar disarray shown in patients and animals,25,26 as well as rat primary cardiomyocytes treated with anthracyclines, including doxorubicin (0.1–0.5 µM). 18
In conclusion, the manual pipetting method with a thin glass pipette allowed us to obtain cardiomyocyte-rich beating CCs from mESC-derived beating EBs and to purify cardiomyocytes by a simple technique. In addition, we demonstrated that this method was also useful for hiPSC-derived beating EBs. Both mES-CMs and hiPS-CMs in beating CCs had higher proliferative activity compared with that in undissociated EBs. Moreover, we showed that a cardiotoxic drug induced myocardial toxicity in the mES-CMs and hiPS-CMs. Further development of this technique by constructing an automatic pipetting device for dissociation of beating EBs produced from ESCs or iPSCs and investigation of the molecular mechanisms underlying the present purification method would accelerate the development of larger scale preparation of functional cardiomyocytes for drug screening.
Footnotes
Acknowledgements
Human iPS cell line 253G1 was kindly provided by Kyoto University under a material transfer agreement. We thank Dr. Hirofumi Nagai for encouragement, Drs. Nobuyuki Nishida and Makoto Miyamoto (Drug Safety Research Laboratories, Takeda Pharmaceutical Company Limited) and Satoshi Yamamoto (Biology Research Laboratories, Takeda Pharmaceutical Company Limited) for their helpful suggestions, and Shingo Ookubo, Ryotaro Hori, Maya Morita, and Yoshiko Okai (Drug Safety Research Laboratories, Takeda Pharmaceutical Company Limited) for their excellent technical assistance.
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
The authors received no financial support for the research, authorship, and/or publication of this article.