
Abstract
Hemophilia A (HA) is an inherited X-linked coagulation disorder caused by the deficiency of factor VIII (FVIII). Linkage analysis is a common indirect method for the detection of female carriers in families with HA. In the current study, 173 patients from 30 unrelated families with HA were recruited from the Azeri Turkish population of northwest Iran and analyzed for BclI and HindIII markers by polymerase chain reaction-restriction fragment length polymorphism. We investigated the potential of using these markers for the detection of mutation in carriers through linkage analysis, which would be of tremendous use in prenatal diagnosis. Among the tested women, 47% and 35% were found to be heterozygous for BclI and HindIII polymorphic markers, respectively. The BclI and HindIII markers were informative for the detection of 63% and 17% potential carriers, respectively, demonstrating the effectiveness of the BclI marker for the detection of HA carriers among the Azeri Turkish population.
Introduction
Hemophilia A (HA; Online Mendelian Inheritance in Man: 3067100) is the most common X-linked recessive blood coagulation disorder, with a worldwide prevalence of 1 in 5000 to 10000 male births among all ethnic groups. It is caused by a partial or total deficiency of clotting factor VIII (FVIII), 1 which is essential for the propagation of the intrinsic coagulation pathway. Therefore, clinical presentation and severity of the disease are directly linked to the level of functional FVIII protein in the plasma. 2 The disease has no permanent treatment; treatment at present involves transfusion with exogenous FVIII, which is not only very expensive but also a chief cause of health care-related concern in developing countries, including Iran. Carrier detection and prenatal diagnosis (PND) are alternate ways of controlling the prevalence of this high-cost disease. 3 Molecular analysis techniques, including the direct and indirect analyses of FVIII gene sequence, have increased the detection rate of HA carriers. It is known that about a third of patients with HA acquire new mutations that were not inherited from their mothers. Although the direct detection of mutations may be the best approach for carrier detection and PND, the highly heterogeneous nature of the mutations and the complexity of the FVIII gene (186 kb length comprising 26 exons) make direct analysis of mutations difficult to achieve in low-resource molecular diagnostic laboratories. 2 –4 Due to these difficulties associated with the direct detection of FVIII mutations, indirect allele characterization using DNA markers linked to the FVIII locus appears to be a more practical approach for the detection of HA carriers. 5 The informative potential of individual polymorphic markers, that is, the heterozygosity rate for each marker, should be investigated in the same population for efficient tracking of the chromosome carrying the mutant gene. 6,7
Certain DNA markers for the detection of carrier status at the FVIII locus were documented by World Health Organization in 1993. 8 As a consequence of genetic variation among different populations, a high degree of genetic differences have been found between various ethnic groups. Therefore, there is a need for finding specific markers linked to the FVIII gene in each population. The population of Iran is composed of different ethnic groups, with Azeri Turkish representing one of the largest ethnic groups in the northwest region of Iran. The current study focuses on the detection of carriers in 30 families in this region with a history of HA, and the usefulness of 2 single nucleotide polymorphism (SNP) markers, BclI and HindIII, for linkage analysis (Figure 1). We used 2 polymerase chain reaction (PCR)-restriction fragment length polymorphism (RFLP) strategies for the detection of female carriers in families with HA. The results of this study would be helpful in genetic counseling, carrier detection, and PND of HA, in particular for the Iranian Azeri Turkish population.
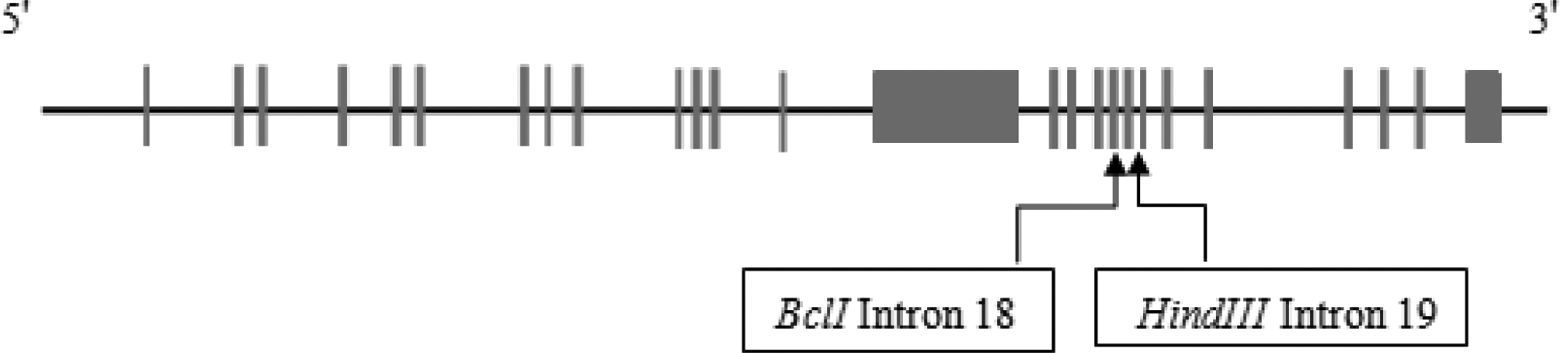
Recognition sites of BclI and HindIII enzymes are placed in introns 18 and 19 of factor VIII (FVIII) gene, respectively, which have been used as intragenic markers for indirect diagnosis of hemophilia A.
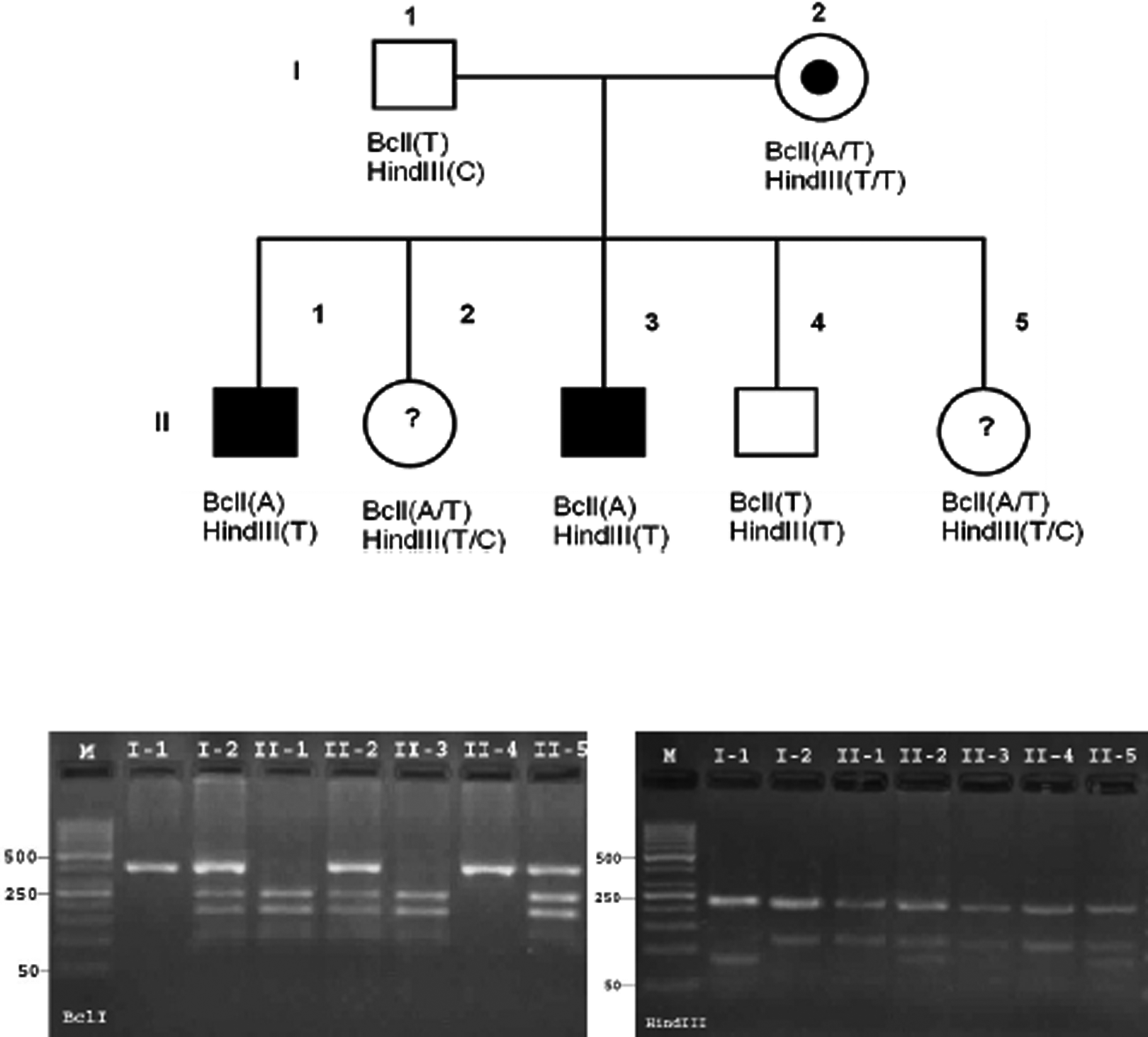
Family pedigree, haplotype analysis, and pictures of their restriction fragment length polymorphism (RFLP) products in a hemophilic A family. Illustrates that BclI marker is informative in this family for mutant chromosome tracking, but HindIII marker is noninformative, because the I-2 is hemozygote for HindIII marker. Mutant associated haplotype is BclI (A)/HindIII (T).
Materials and Methods
Patients
This study included 30 families from the Iranian Azeri Turkish population with at least 1 member diagnosed with HA. A total of 173 individuals, including 34 patients with HA, 104 male and female nonhemophiliac family members, and 35 potential carriers (women), were referred from the Hemophilia Center to the Outpatient Clinic of Tabriz University of Medical Sciences. Following approval of the study by the Ethics Committee of Tabriz University of Medical Sciences, informed consent was obtained from all the family members, and 3 to 5 cm3 of whole blood was collected and stored at 4°C until further processing.
DNA Extraction and Genotyping
Genomic DNA was extracted from whole blood samples using the salting out method. 9 The purity and concentration of the isolated DNA samples were measured using spectrophotometry. The PCR amplification was carried out in a final reaction volume of 25 μL, with the reaction mixture containing ∼100 ng of genomic DNA, 4 pmol/mL of each primer, 2.5 μL of 10× PCR buffer, 0.25 mmol/L of each deoxynucleotide triphosphate, 1.5 mmol/L MgCl2, and 2 units of Taq DNA polymerase (CinnaGene Inc., Iran). The PCR conditions included an initial denaturation at 94°C for 5 minutes, followed by 30 cycles of denaturation at 94°C for 1 minute, annealing at 62°C and 65°C for BclI and HindIII, respectively, for 1 minute, and extension at 72°C for 1 minute, followed by a final extension at 72°C for 5 minutes. Primer sequences were designed based on the reference sequence (RefSeq accession no: NM_000132.3) 10 and are provided in Table 1. The exact location of the SNPs under study were determined using NEBcutter online tool 11 (BioLabs Inc., New England) to ensure that these SNPs are located within the amplified region and to predict the sizes of the digested PCR products. Following PCR, amplification of the target sequence was confirmed using 1% agarose gel electrophoresis. The products were then subjected to digestion using the restriction enzymes HindIII or BclI (MBI Fermentas) according to the manufacturer’s protocol and the products were separated by electrophoresis using 2% agarose gel. The following fragment sizes were predicted on the basis of in silico analysis using the Webcutter tool (1) for BclI RFLP, a 383-bp fragment for the indigestible allele, and 2 fragments of 156 bp and 227 bp for the digestible allele and (2) for HindIII RFLP, a common 214-bp fragment in addition to 2 fragments of 81 bp and 36 bp for the digestible allele and a 117-bp fragment for the indigestible allele.
The Sequences of Deigned Primers for Bcl1 and HindIII Markers.a
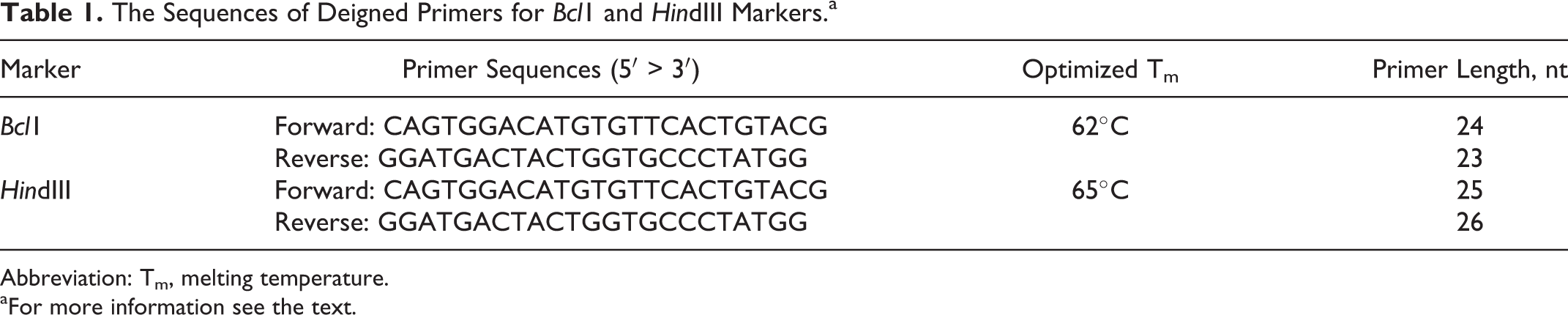
Abbreviation: Tm, melting temperature.
aFor more information see the text.
Statistical Analyses
Allele frequencies for the polymorphic sites were calculated based on the number of X chromosomes and the presence or absence of a restriction site.
The expected heterozygosity rates were calculated using the Hardy-Weinberg equation. A chi-square test was performed to examine differences between the observed and expected heterozygote frequencies. 12 A P value of .05 or less was considered statistically significant. Polymorphism information content (PIC) was calculated according to the method reported by Botstein et al. 13
Results
A total of 173 individuals from 30 families with a history of HA were examined. The results of our analyses of the BclI and HindIII polymorphic sites are provided in Table 2. The allele frequencies of the “+” (digestible by BclI) and “−” (indigestible by BclI) alleles were found to be approximately 31% and 69%, respectively. Of the women tested for this polymorphism, 47% were found to be heterozygous and 63% of the potential carriers were identified using this marker alone.
Allele Frequency and Heterozygosity Rate of BclI and HindIII Markers in Azeri Turkish Population of Iran.
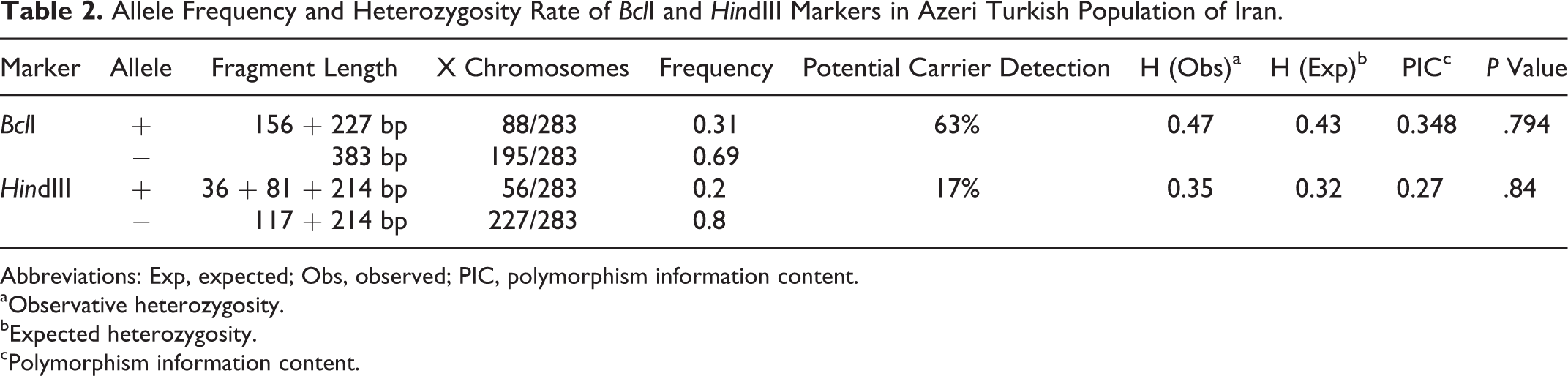
Abbreviations: Exp, expected; Obs, observed; PIC, polymorphism information content.
aObservative heterozygosity.
bExpected heterozygosity.
cPolymorphism information content.
The HindIII marker, on the other hand, showed a lower rate of heterozygosity among the women studied. The allele frequency of the “+” allele (digestible by HindIII) was found to be 19.8%. Approximately 35% of the women tested were heterozygous for this locus and carrier status was identified in 17% of the potential carriers. Using pedigree analysis, we determined 30 mutant and 60 wild-type, nonidentical X chromosomes (Table 3).
Recognized Haplotype Frequencies of 30 Mutant and 60 Nonmutant Nonidentical X Chromosomes.

Of the 4 possible haplotypes defined by these 2 polymorphic sites of FVIII gene (Table 3), the prevalence of haplotype IV or the BclI(−)–HindIII(−) haplotype was found to be the highest, accounting for 25 (41.6%) and 23 (76.7%) of the wild-type or mutant chromosomes, respectively. The observed frequencies of haplotypes I and III were 25% and 13.3%, respectively, for wild-type chromosomes and 0% for the mutant chromosomes. The PIC values for these 2 markers BclI and HindIII are 0.348 and 0.27, respectively.
Discussion
A direct molecular diagnosis of gene defects in HA is often not easy due to the large size, structural complexity, and high frequency of various types of mutations of the FVIII gene; linkage analysis with RFLP is hence considered the most useful approach. The population of Iran is known to comprise different ethnic groups of diverse genetic backgrounds. Therefore, obtaining data about genetic variations in different regions of this country would be valuable for formulating an appropriate strategy for the detection of HA carriers and also for PND.
The present study was conducted to reveal the relative contribution of the 2 polymorphic restriction sites (BclI/intron 18 and HindIII/intron 19), both separately and within haplotypes, for indirect genetic analyses of HA carriers among the Azeri Turkish population of Iran and represents the first such study on this population. The allele frequencies of BclI (T/A) and HindIII (C/T) alleles were 0.69/0.31 and 0.20/0.80, respectively.
Significant differences were detected between the BclI (T/A) allele frequency observed in the current study (0.69/0.31) and those reported in previous studies for various populations, such as Japanese (0.84/0.16; P = .012), 14 caucasian (0.23/0.77; P < .0001), 8,15,16 and Indian (0.43/0.57; P = .0002) 17 but not Brazilian (0.61/0.39; P = .23). 18 In addition, significant differences were also observed in the HindIII (C/T) allele frequencies between the present study (0.20/0.80) and Brazilian (0.42/0.58; P = .0007) 18 or Indian (0.38/0.62; P = .005) 17 populations. In contrast, the HindIII (C/T) allele frequencies that we observed were consistent with those of caucasian (0.26/0.74; P = .3) 8,15,16 and Japanese (0.19/0.81; P = .86) 14 populations. Furthermore, the allele frequencies for these markers in our surveyed population demonstrated significant differences with previously reported results on a sample population composed of 15 families with HA who were referred from different regions of Iran; the frequencies in the previous study were determined to be BclI (T/A) = 0.52/0.48, P = .013 and HindIII (C/T) = 0.48/0.52, P < .0001. 19 The results of the present study also showed that the HindIII polymorphic site was less informative in this population than in the earlier study, also conducted on Iranian population, wherein the heterozygosity of HindIII was reported to be 0.46. 19 This discrepancy in the data could be due to differences in sampling, which was done from different Iranian ethnic groups in the previous study, but only from the Azeri Turkish ethnic group in the current study.
The efficiency of the HindIII polymorphic restriction site was found to be quite low even among the Japanese population, with a heterozygosity rate of 0.3, 14 unlike in the caucasian, Indian, or Brazilian populations with values of 0.38, 8,15,16 0.49, 17 and 0.49, 18 respectively.
The efficiency of BclI RFLP in detecting carriers varied between different ethnic groups: 28% in Japanese, 14 35% in caucasian, 8,15,16 54% in Indian, 17 and 47% in Brazilian 18 populations. In our current investigation, BclI locus displayed a high heterozygosity rate of 47%, consistent with the previous study in Iran also with a reported rate of 47%. 19 Our results support the conclusion that BclI is more informative than other studied markers. Accordingly, BclI-RFLP site was efficacious in detecting 63% of the potential carriers, and HindIII-RFLP site, 18%, although no further advantage of using a second intragenic FVIII polymorphism marker could be observed.
The genetic informative potential was assessed using PIC, which represents the sum of frequency of each possible mating multiplied by the probability that an offspring will be informative. The heterozygosity and PIC values for these 2 markers demonstrate that the BclI polymorphic marker is a better predictor of HA carriers in the Azeri Turkish population than the HindIII marker.
Footnotes
Acknowledgments
The authors would like to thank Research Deputy of Faculty of Medicine affiliated to Tabriz University of Medical Sciences for financial support.
Authors’ Note
This article is a report of a database from dissertation with code: 89/2-7/24 registered in Tabriz University of Medical Sciences, Tabriz, Iran.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.