
Abstract
Platelets play an important role in various thrombotic diseases, including myocardial infarction. Because red wine consumption is inversely associated with death due to ischemic heart diseases, the effects of grape components on platelet function have been extensively investigated. Grape seed extracts (GSEs) reportedly inhibit platelet aggregation; however, the underlying mechanism has not been elucidated. We discovered that GSEs inhibit platelet aggregation induced by collagen and thrombin-receptor agonist peptide and increase basal levels of tyrosine phosphorylation, which was also observed in the presence of a protein tyrosine phosphatase (PTP) inhibitor. An in vitro phosphatase assay indicated that GSE dose dependently inhibited PTP-1B and Src homology 2 domain-containing phosphatase-1 activity, which positively regulates platelet aggregation. We propose that GSEs inhibit platelet aggregation by inhibiting tyrosine phosphatase activity. Moreover, we showed that GSE ingestion inhibited platelet aggregation in mice without enhancing tail bleeding, implying that GSE supplementation might be beneficial to prevention of thrombotic diseases.
Keywords
Introduction
Platelets play a pivotal role in physiological hemostasis and pathological thrombosis such as stroke and myocardial infarction. Platelets interact with exposed collagen fibers at the sites of vessel injury. 1 The collagen receptor glycoprotein VI (GPVI) initiates cellular activation that allows fibrinogen to bind to integrin and triggers the release of adenosine diphosphate (ADP) and thromboxane A2 (TXA2). The released agonists activate additional flowing platelets and mediate thrombus growth by bridging platelets through integrin–fibrinogen interactions. Activated platelet surfaces serve as platforms for clotting factors that accelerate thrombin generation. Thrombin, in turn, directly activates platelets and converts fibrinogen into fibrin, which consolidates platelets into aggregates. Soluble agonists, including ADP, TXA2, and thrombin, induce platelet aggregation through G protein-coupled receptors (GPCRs). On the other hand, GPVI and integrin are tyrosine kinase-linked receptors, both of which utilize Src family tyrosine kinases as upstream signaling molecules. Thrombus formation is regulated by the coordinated action of tyrosine kinase-linked receptor and GPCR.
Red wine was the first polyphenol-rich food reported to have an inverse association with deaths due to ischemic heart disease in industrialized countries. 2,3 French people experience relatively low incidence of coronary heart disease, despite their diets that are relatively rich in saturated fats (French paradox). 3 The high red wine consumption in France has been suggested as a primary factor responsible for this trend. 3 Thus, many studies have focused on the effects that consumption of wine components has on platelet function. Resveratrol and quercetin are well-known, pure polyphenolic compounds contained mainly in grape skins. Resveratrol reportedly inhibits platelet aggregation by inhibiting TXA2 production 4 or generating nitric oxide. 5 Quercetin also reportedly inhibits platelet aggregation by inhibiting protein tyrosine kinase. 6,7 More recently, human studies have shown that polyphenol-rich crude extracts from grape seeds or skins inhibit platelet aggregation that was induced by thrombin-receptor agonist peptide (TRAP) or collagen, 8 –10 and that ingestion of grape seed extract (GSE) inhibits ADP or collagen-induced platelet aggregation in humans. 11,12 However, the mechanism of GSE inhibition of platelet activation has not been fully elucidated to date. In this study, we investigated the effects of GSE from Muscat Bailey A (MBA) or Cabernet Sauvignon (CS) grapes on platelet activation. We discovered that MBA and CS inhibit collagen- and TRAP-induced platelet aggregation by inhibiting protein tyrosine phosphatase (PTP), and we suggest that this may represent a novel mechanism for the inhibitory effects of polyphenols on platelet aggregation. Moreover, we showed that ingestion of MBA and CS inhibits platelet aggregation in mice without prolonged tail bleeding, which implies that GSE supplementation may be beneficial for the prevention of thrombotic diseases such as myocardial and cerebral infarction.
Aim
Although it has been reported that ingestion of GSE inhibits platelet aggregation, its mechanism has not been fully elucidated to date. The aim of this study is to investigate the mechanisms of inhibitory effects of GSE on platelet aggregation.
Materials and Methods
Materials
The TRAP (Ser-Phe-Leu-Leu-Arg-Asn: SFLLRN peptide) was obtained from Bachem Biochemica (Heidelberg, Germany). The PTP inhibitor II (4-methoxyphenacyl Br) was purchased from Calbiochem (Bad Soden, Germany). Mouse anti-phosphotyrosine antibody (4G10), recombinant PTP-1B, and re-combinant Src homology 2 domain-containing phosphatase-1 (SHP-1) were obtained from Millipore (Massachusetts). The Gly-Arg-Gly-Asp-Ser (GRGDS) peptide was purchased from Peptide Institute (Osaka, Japan). Resveratrol, quercetin, and rutin were obtained from Sigma Aldrich (St Louis, Missouri). All other reagents were obtained from sources named previously. 13
Extraction of Grape Seed Components
For extraction of GSE, grape seeds were obtained from grapes grown to commercial maturity in an experimental vineyard of University of Yamanashi. The cultivar included MBA (Bailey × Muscat Humburg, hybrid) and CS (Vitis vinifera). Seed samples were air dried and ground into fine powder with a grinder (Wonder Blender WB-1; Osaka Chemical Co, Osaka, Japan). The powder (300 g) was deoiled with hexane and then extracted 3 times with 50% aqueous ethanol (1000 mL). The extract was partitioned 3 times with an equal volume of ethyl acetate. The ethyl acetate layer was collected, evaporated under vacuum, and then lyophilized. The resulting lyophilizate was weighed and then used as GSE. The recovery of ethyl acetate extracts from MBA and CS seeds was 0.8% and 2.0%, respectively.
Platelet Preparation and Platelet Aggregation
Venous blood was collected from healthy, drug-free volunteers into 10% sodium citrate tubes. This study was approved by the ethical committees of the University of Yamanashi. Informed consent was obtained according to the Declaration of Helsinki. Washed human platelets (2 × 108/mL) were obtained as described previously 13 and preincubated with the indicated concentrations of resveratrol, rutin, quercetin, MBA, CS, or PTP inhibitor II for 5 minutes at 37°C. Finally, platelets were stimulated with the indicated concentrations of collagen or TRAP. Platelet aggregation was monitored by measuring light transmission for 5 minutes with an AG-10 aggregation analyzer (Kowa, Tokyo, Japan).
Platelet Biochemistry
Washed platelets (5 × 108/mL) were pretreated for 5 minutes with 1 mmol/L GRGDS peptide and 10 μg/mL indomethacin at 37°C to inhibit the secondary effects of platelet aggregation or TXA2 release. Next, the platelets were incubated with dimethyl sulfoxide (DMSO) or 50 μg/mL quercetin, resveratrol, MBA, CS, or 10 μmol/L PTP inhibitor II for 5 minutes at 37°C. Platelets were stimulated with 20 μg/mL collagen for 5 minutes. Reactions were terminated by the addition of 4 × sodium dodecyl sulfate (SDS) sample buffer, and proteins were separated using 8% SDS polyacrylamide gel electrophoresis (SDS-PAGE), electrotransferred, and subjected to Western blot analysis with anti-phosphotyrosine antibody as described previously.13
In Vitro PTP Assay
A PTP-1B microtiter assay was performed using p-nitrophenyl phosphatase (pNPP) hydrolysis according to the manufacturer’s instructions (Millipore). Briefly, 10 μL of recombinant PTP-1B (1.5 μg) was preincubated with DMSO, or the indicated concentrations of MBA or CS in assay buffer (25 mmol/L 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid [HEPES]), 50 mmol/L NaCl, 5 mmol/L dithiothreitol, 2.5 mmol/L EDTA, pH 7.2) with BSA (100 μg/mL) for 10 minutes in a 96-well microtiter plate. The reactions were initiated by the addition of 20 μL pNPP (50 mmol/L) and 80 μL assay buffer at 37°C and stopped after 10 minutes by the addition of 2 mol/L Na2CO3. The PTP-1B activity was determined by measuring the absorbance at 405 nm in 50-μL diluted aliquots with a Bio-Rad Novapath microplate reader (Bio-Rad Laboratories, Inc., CA, USA). The assay for SHP-1 activity was conducted according to the manufacturer’s instruction (Millipore) with 6,8-difuoro-7-hydroxy-4-methylcoumarin (DIFMU) as a substrate. The SHP-1 (1 ng) was incubated with DMSO or the indicated concentrations of MBA or CS, phosphorylated DIFMU was then added, and the mixture was incubated for 60 minutes at 22°C. The SHP-1 activity was determined by measuring the fluorescence intensity with a SPECTRAmax GEMINI microplate fluorescence reader (Molecular Devices Japan, Tokyo, Japan; excitation 358 nm, emission 455 nm).
Animals, Murine Platelet Aggregation, and Tail Bleeding
Eight-week-old male C57BL/6NJcl mice were purchased from CREA Japan, Inc (Tokyo, Japan). All procedures were approved by the Animal Experiment Committee, University of Yamanashi. The MBA or CS (200 mg/kg) dissolved in 200 μL saline or the same volume of saline was administered to mice orally with a plastic feeding needle. At 1 and 3 hours after ingestion, blood samples were collected from mice. Washed platelets (2 × 108/mL) were prepared, and platelet aggregation was monitored as described previously.13 These washed platelets (2 × 108/mL) were pretreated for 5 minutes with 1 mmol/L GRGDS peptide and 10 μg/mL indomethacin at 37°C. Platelets were stimulated with or without 20 μg/mL collagen for 5 minutes. Reactions were terminated by the addition of 4× SDS sample buffer. Proteins were separated using 4% to 12% SDS-PAGE, electrotransferred, and subjected to Western blot analysis with anti-phosphotyrosine antibody. Throughout the experiments, mice were anesthetized with sevoflurane via a face mask 90 minutes after ingestion. Fifteen minutes following the administration of anesthesia, the 3-mm tip of the tail was cutoff with a sharp razor blade; the volume of blood loss was measured for 10 minutes.
Statistics
Statistical significance was evaluated by Student t test. In each case, P values <.05 were taken as the minimum to indicate statistical significance.
Results
Grape Seed Extracts From MBA and CS Inhibit Collagen- and TRAP-Induced Platelet Aggregation in a Dose-Dependent Manner
Grape seed extracts obtained from MBA and CS grapes were compared to pure polyphenolic compounds, resveratrol, quercetin, and rutin for their effects on platelet aggregation. Both collagen- and TRAP-induced types of platelet aggregation were reduced by approximately 50% in the presence of either 10 μg/mL MBA or CS (Figure 1A and B). Resveratrol and quercetin, both of which are polyphenols obtained from purified grapes, inhibited collagen-induced platelet aggregation by 50% at concentrations of 0.38 or 2.8 μg/mL, respectively, whereas concentrations that were 5 times greater were required for the inhibition of TRAP-induced aggregation (Figure 1A and B). Thus, the inhibitory profile of pure polyphenolic compounds is distinct from that of MBA and CS. In addition, the concentrations of resveratrol and quercetin in MBA and CS were below the level of detection (data not shown). Therefore, the inhibitory effects of MBA and CS cannot be explained by resveratrol and quercetin. In marked contrast, rutin, which is a polyphenolic compound isolated from buckwheat, showed no inhibitory effects on platelet aggregation even with the maximum concentrations of polyphenols (50 μg/mL) used in this study (Figure 1A and B). Rutin, resveratrol, quercetin, MBA, and CS possess antioxidant activity, but rutin did not inhibit platelet aggregation, which suggests that the inhibitory effects of the other 2 pure polyphenols and MBA and CS cannot be fully explained by their antioxidant activity.
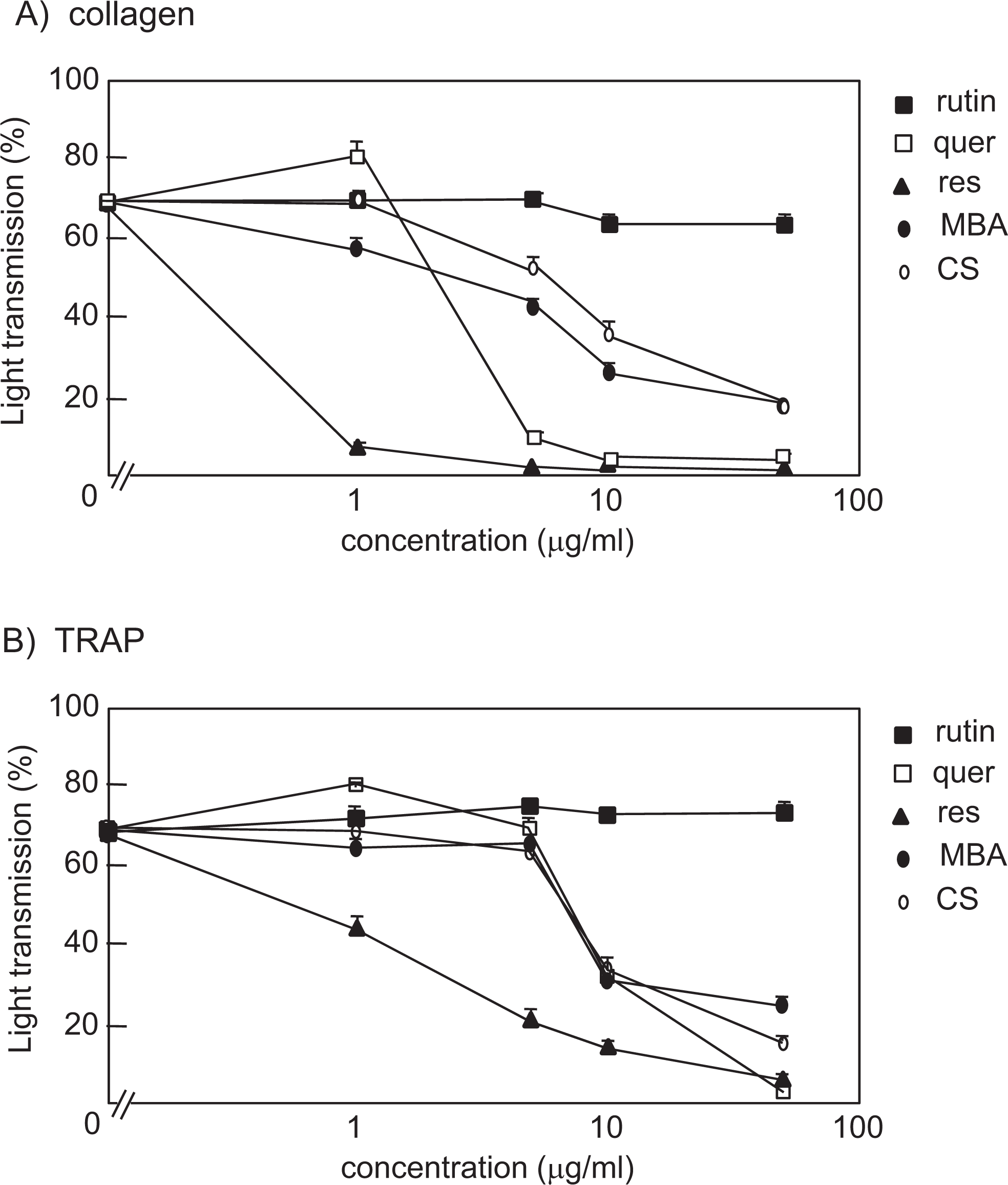
Muscat Bailey A (MBA) and Cabernet Sauvignon (CS) cause dose-dependent inhibition of platelet aggregation induced by collagen or thrombin-receptor agonist peptide (TRAP). Washed platelets were pretreated for 5 minutes with 0, 1, 5, 10, or 50 µg/mL of rutin, quercetin, resveratrol, MBA, or CS. They were stimulated with 1 µg/mL of collagen (A) or 30 µmol/L of TRAP (B). Platelet aggregation was measured for 5 minutes by light transmission. Data are expressed as means of maximal light transmission ± standard error (n = 5-10, average from 3 experiments).
Muscat Bailey A- and CS-Enhanced Protein Tyrosine Phosphorylation in Unstimulated Platelets and in Collagen-Stimulated Platelets
We investigated potential mechanisms for the inhibitory effects imparted by MBA and CS on platelet aggregation. Collagen induces platelet activation that is dependent on the Src family of tyrosine kinases and stimulates a marked increase in tyrosine-phosphorylated protein production,14,15 whereas thrombin stimulates only minor tyrosine phosphorylation. Therefore, we first investigated GSE for its effects on protein tyrosine phosphorylation during collagen-induced platelet aggregation. After collagen stimulation, increased tyrosine phosphorylation was observed on multiple signaling molecules; tyrosine phosphorylation was greatly inhibited in the presence of quercetin, which is consistent with previous reports (Figure 2A).6,7 Resveratrol had no effects on protein tyrosine phosphorylation (Figure 2A). In marked contrast, both MBA and CS caused increases in basal levels of tyrosine phosphorylation in multiple proteins (Figure 2A).
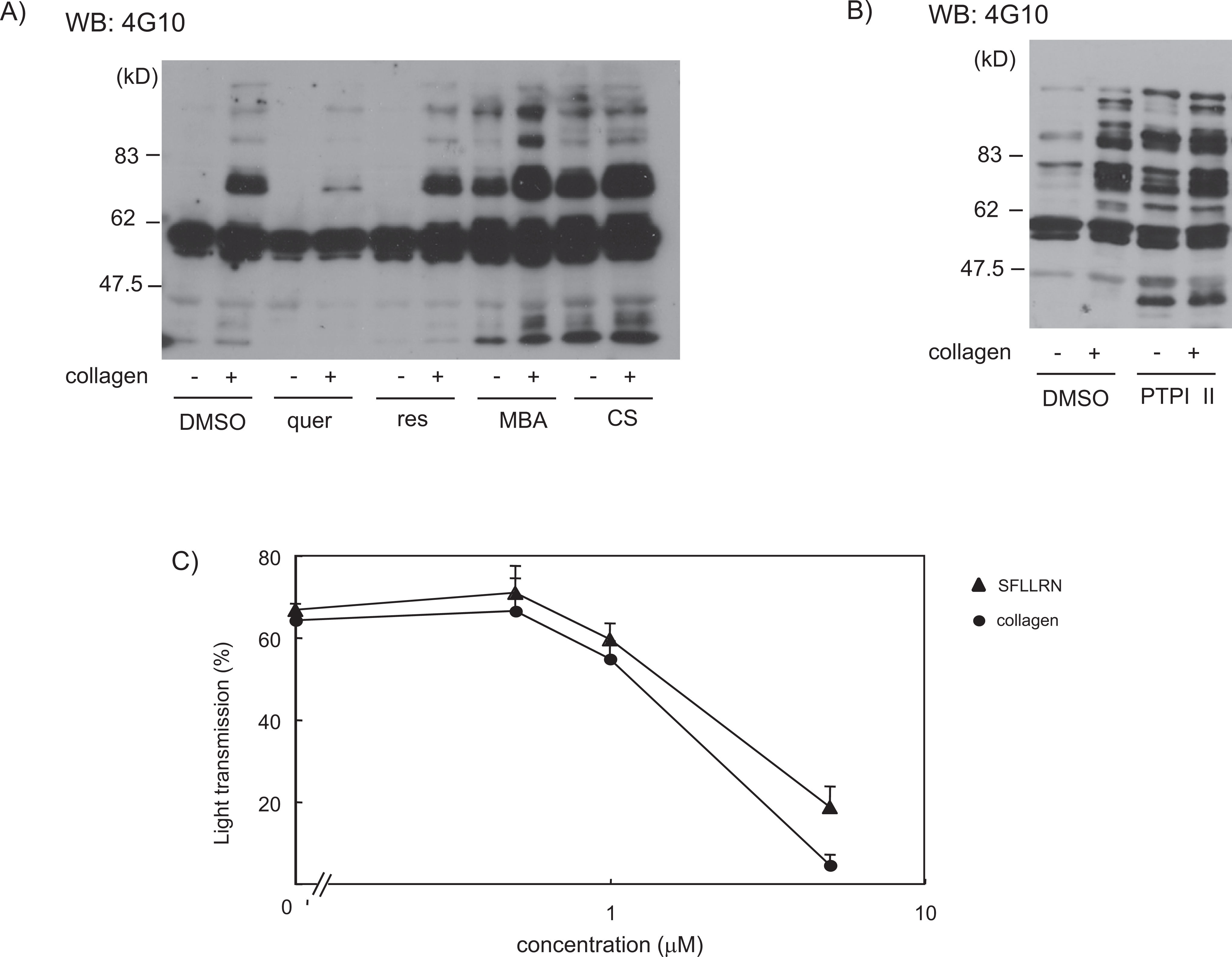
Muscat Bailey A (MBA), Cabernet Sauvignon (CS), and PTP inhibitor-II increase basal levels of tyrosine phosphorylation in platelets. Washed platelets were pretreated with 1 mmol/L GRGDS and 10 µmol/L indomethacin to inhibit secondary effects of platelet aggregation and TXA2 release; then they were incubated for 5 minutes at 37°C with DMSO or 50 µg/mL quercetin, resveratrol, MBA, or CS (A) or 10 µmol/L PTP inhibitor-II (B). Next, they were stimulated with 20 µg/mL collagen for 5 minutes. Reactions were terminated by the addition of 4× SDS sample buffer, and the proteins were separated by 8% SDS-PAGE, electrotransferred, and subjected to Western blot with anti-phosphotyrosine antibody (4G10). The data represent 3 experiments. C, Washed platelets were pretreated for 5 minutes with 0, 0.5, 1, or 5 µmol/L PTP inhibitor-II. They were stimulated with 1 µg/mL collagen or 30 µmol/L TRAP. Platelet aggregation was measured for 5 minutes by light transmission. Data are expressed as the mean of maximal light transmission ± standard error (n = 5 from 3 experiments). PTP indictaes protein tyrosine phosphatase; MBA, Muscat Bailey A; CS, Cabernet Sauvignon; DMSO, dimethyl sulfoxide; GRGDS, Gly-Arg-Gly-Asp-Ser; SDS-PAGE, sodium dodecyl sulfate polyacrylamide gel electrophoresis.
It is well known that agonist stimulation activates protein tyrosine kinases in platelets, whereas some PTPs are active in the resting (nonstimulated) state. Therefore, we hypothesized that MBA and CS inhibit platelet aggregation by inhibiting the activity of PTP. To investigate this hypothesis, we compared MBA and CS with PTP inhibitor II, which is a commercially available inhibitor of tyrosine phosphatases (eg, SHP-1), for their effects on protein tyrosine phosphorylation and platelet aggregation. We discovered that PTP inhibitor-II enhances basal levels of tyrosine phosphorylation, which is similar to the results that manifest in the presence of MBA and CS. Preincubation with PTP inhibitor-II led to almost equal inhibition of collagen- or TRAP-induced platelet aggregation in a dose-dependent manner (Figure .2C), which was also the case for MBA and CS (Figure 1). These findings suggest that MBA and CS inhibit the activity of PTPs, which regulate tyrosine kinase activity in platelets.
Muscat Bailey A and CS Inhibited In Vitro Activity of Tyrosine Phosphatases PTP1B and SHP-1
We used an in vitro tyrosine phosphatase assay to prove directly that MBA and CS inhibit tyrosine phosphatase activity. Cytosolic PTP-1B lies upstream of Src and is essential for integrin αIIbβ3-mediated Src activation. 16 The SHP-1 is another cytosolic tyrosine phosphatase that is expressed in platelets; it positively regulates GPVI-mediated platelet activation. 17 An in vitro assay on PTP-1B activity showed that preincubation of MBA or CS with recombinant PTP1B inhibits phosphatase activity in a dose-dependent manner (Figure 3A). Recombinant SHP-1 activity was also inhibited in a dose-dependent manner in the presence of MBA or CS (Figure 3B). All these findings support the hypothesis that MBA and CS inhibit platelet aggregation by inhibiting PTPs.
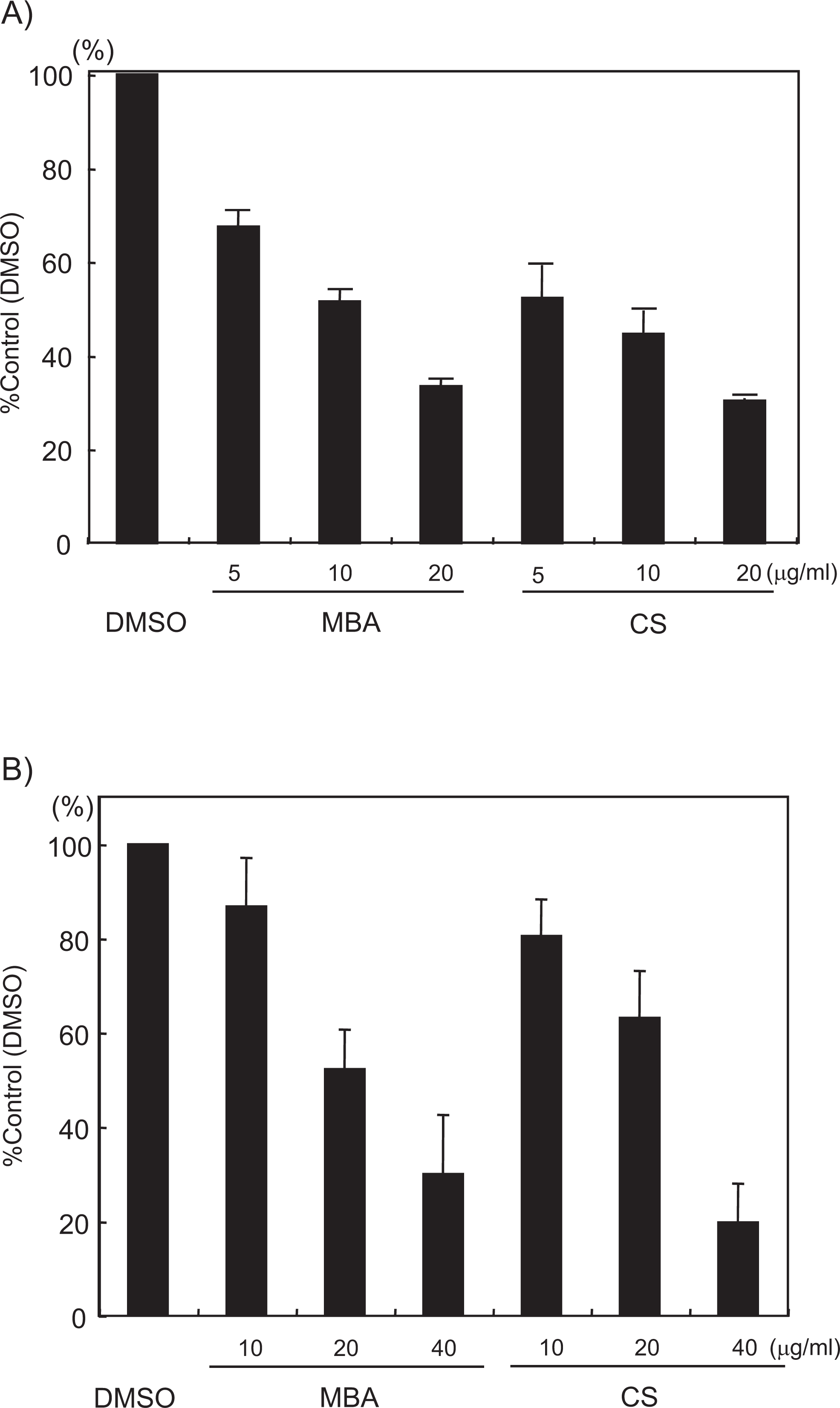
Muscat Bailey A (MBA) and CS cause dose-dependent inhibition of PTP-1B and SHP-1 activity. (A) Recombinant PTP-1B was preincubated with DMSO or the indicated concentrations of MBA or CS, and then incubated with pNPP for 10 minutes at 37°C. PTP-1B activity was determined by measuring pNPP absorbance on a microplate reader. (B) SHP-1 was incubated with DMSO or the indicated concentrations of MBA or CS. Then, phosphorylated DIFMU was added and incubated for 60 minutes at 22°C. SHP-1 activity was determined by measuring the fluorescence intensity of DIFMU using a microplate fluorescence reader. Data are expressed as the mean of percent production ± standard error (n = at least 6, average from 2–3 experiments). PTP, protein tyrosine phosphatase; MBA, Muscat Bailey A; CS, Cabernet Sauvignon; pNPP, p-nitrophenyl phosphatase; DIFMU, 6,8-difluoro-7-hydroxy-4-methylcoumarin; SHP-1, Src homology 2 domain-containing phosphatase-1; DMSO, dimethyl sulfoxide.
Muscat Bailey A and CS Ingestion Inhibited Collagen-Induced Platelet Aggregation, But Did Not Enhance Tail Bleeding in Mice
Although 2 previous studies have shown GSE ingestion inhibits collagen- or ADP-induced platelet aggregation in humans,11,12 the question of whether GSE supplementation increases bleeding tendencies has not yet been investigated. Therefore, we examined whether oral administration of MBA or CS enhances tail bleeding or inhibits platelet aggregation in mice. Blood samples were collected from mice 1 and 3 hours after ingestion of MBA or CS (200 mg/kg) or saline, and collagen-induced platelet aggregation was monitored with an aggregometer. The TRAP was omitted from the mouse experiments because proteinase-activated receptor-1 (PAR-1), the target of TRAP, is not expressed on murine platelets. Both MBA and CS were shown to inhibit collagen-induced platelet aggregation significantly by 1 hour after administration, whereas by 3 hours, the inhibitory effect of MBA or CS was not statistically significant (Figure 4A). We investigated whether oral ingestion of GSEs inhibits protein tyrosine phosphorylation in mice. Protein tyrosine phosphorylation in resting and stimulated platelets was minimally increased 1 hour after oral ingestion of GSE compared to saline-ingested mice (data not shown) as is the case of in vitro incubation of washed human platelets with GSEs (Figure 2A). However, the increase in phosphorylation observed in oral ingestion in mice is minimal compared to in vitro incubation of platelets with GSEs (Figure 2A).
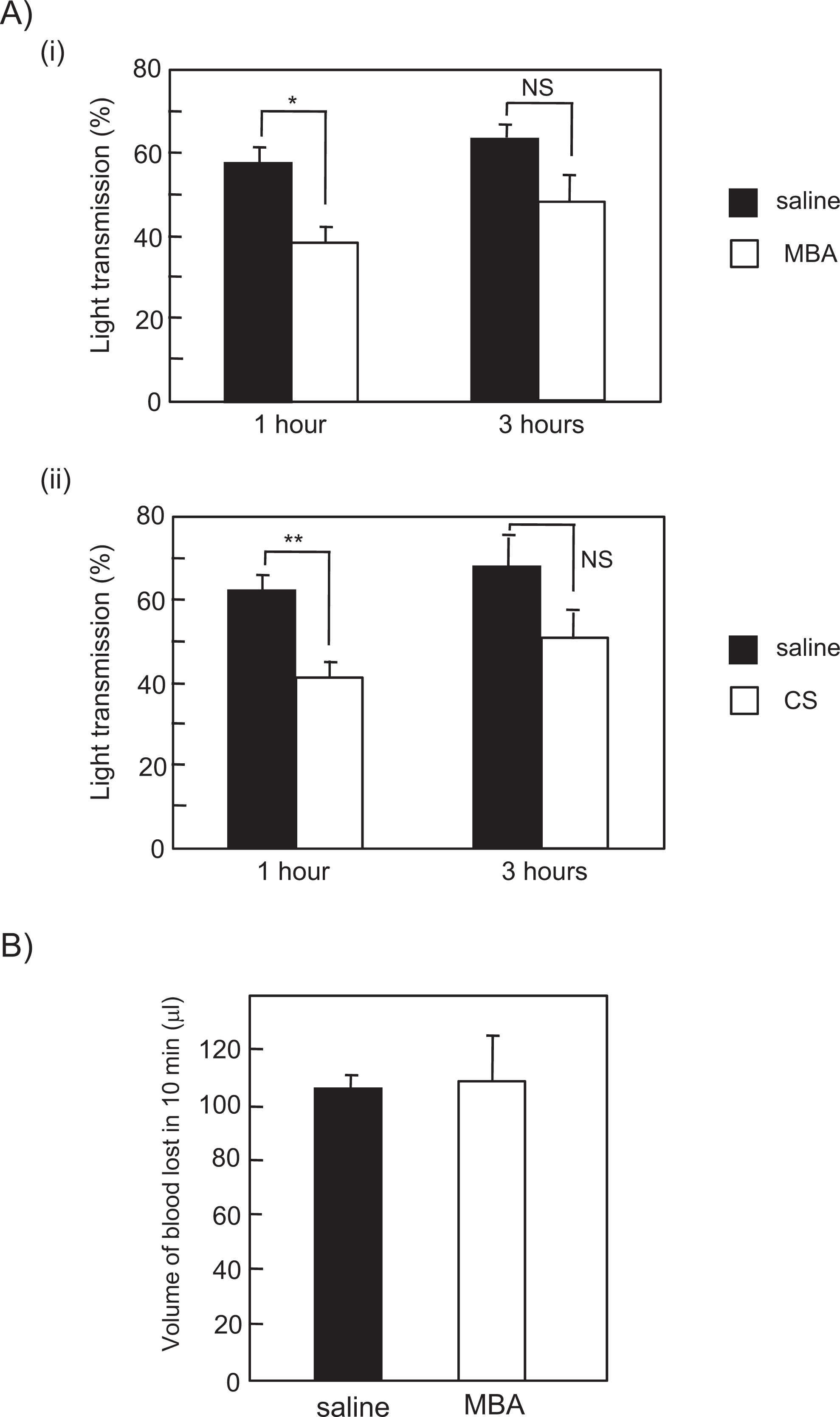
Ingestion of Muscat Bailey A (MBA) and Cabernet Sauvignon (CS) inhibited platelet aggregation in mice without enhancing tail bleeding. A, (i) MBA (200 mg/kg), (ii) CS (200 mg/kg), or saline were administered orally to mice with a plastic feeding needle. At 1 and 3 hours post-ingestion, blood samples were collected from mice, and the platelets were washed and prepared. Platelet aggregation induced by collagen (2 µg/mL) was monitored by light transmission. Data are expressed as the mean of maximal light transmission ± standard error (n = 6, average from 2–3 experiments). B, Ninety minutes after ingestion, mice were anaesthetized with isofluorane through a face mask. Fifteen minutes after the anesthesia was administered, the 3-mm tip of the tail was cutoff with a sharp razor blade, and the volume of blood loss was measured for 10 minutes. Data are expressed as the mean ± standard error (n = 3 mice).
Ninety minutes following oral administration, the volume of blood loss from a small tail incision was measured for 10 minutes. No significant differences were evident between mice fed MBA/CS and saline (Figure 4B), which suggests that ingestion of MBA or CS inhibits platelet activation but not bleeding tendency. The GSE supplementation may be beneficial for the prevention of thrombotic diseases such as myocardial and cerebral infarction.
Discussion
In the present study, we proposed that MBA and CS inhibit platelet aggregation by inhibiting PTP. Here, we showed that (1) MBA and CS cause dose-dependent inhibition of platelet aggregation that is mediated by GPVI and PAR-1, (2) MBA/CS and PTP inhibitor II increase basal levels of tyrosine phosphorylation in platelets, and (3) MBA and CS inhibit in vitro activation of PTP-1B and SHP-1. These observations support the hypothesis that MBA and CS inhibit platelet aggregation by inhibiting PTP activity. Purified polyphenols from grapes or GSEs have been investigated intensively for their effects on platelet aggregation and to understand the mechanisms that underlie the “French paradox.” Quercetin reportedly inhibits platelet aggregation by either inhibiting protein tyrosine phosphorylation6,7 or enhancing platelet nitric oxide production. 18 Resveratrol reportedly inhibits platelet aggregation by either inhibiting TXA2 production 4 or generating nitric oxide. 5 Crude GSEs were also reported to inhibit superoxide release and enhance nitric oxide generation. 9 However, no reports have described how polyphenols or GSEs inhibit PTP activity.
In platelets, several PTPs play important roles in platelet activation by activating the Src family of kinases. Cytosolic PTP-1B is essential for integrin αIIbβ3-mediated Src activation,16,19 whereas cytosolic SHP-1 positively regulates Src and GPVI-mediated platelet activation. 17
Because MBA and CS inhibited the activity of PTP1B and SHP-1 in vitro (Figure 3), we suspect that MBA and CS have greater inhibitory effects on GPVI-mediated platelet aggregation than GPCR-mediated platelet aggregation. However, MBA and CS caused almost equal inhibition of platelet aggregation that was mediated by GPVI or PAR-1. Recently, the receptor-like PTP CD148 has been reported to positively regulate the downstream activation of signals of both tyrosine kinase-linked receptor and GPCR. 20 It is possible that MBA and CS also inhibit CD148 activity. Consistent with this notion, we showed that PTP inhibitor-II, a chemical inhibitor that mainly acts on SHP-1 and other tyrosine phosphatases, inhibits collagen- and TRAP-induced platelet aggregation (Figure 2C) and increases basal levels of tyrosine phosphorylation (Figure 2B).
We suggest that some components other than resveratrol and quercetin are responsible for the inhibitory effects of MBA and CS, but we have not yet determined which components in MBA and CS inhibit PTP activity. Multiple components in GSEs may inhibit the phosphatase activity synergistically. In fact, polyphenols quercetin and catechin have been reported to act synergistically in the reduction of platelet activation by inhibiting NADPH oxidase activation. 18 With regard to the polyphenols lower in molecular weight, an increasing body of evidence shows that they are absorbed and metabolized rapidly, which results in a pervasive presence of metabolites in circulation. 21 Moreover, most studies have shown that the metabolite quantity for any polyphenol reaches its peak in the plasma within 2 to 4 hours after consumption. 21 In the present study, ingestion of MBA and CS by mice inhibited platelet aggregation 1 hour after administration, but the effect was not significant at 3 hours, which suggests that certain components of MBA and CS, but not their metabolites, inhibit platelet aggregation. We observed increase in protein tyrosine phosphorylation of resting and stimulated platelets in GSE-ingested mice, compared to saline-ingested mice (data not shown), suggesting that oral ingestion of GSEs may also inhibit the activity of PTP. However, the increase in phosphorylation observed in mice orally administered with GSEs is only minimal compared to in vitro incubation of human platelets with GSEs (Figure 2A). This may be partly because of poor absorption of GSEs, rapid degradation of GSEs, or species difference (humans and mice).
We also showed that ingestion of MBA and CS by mice inhibits platelet aggregation without enhancing tail bleeding probably because the antiplatelet effect is minimal. The inhibitory effects of MBA and CS on platelet aggregation were short acting and had no effects on tail bleeding, both of which point to the potential merit of using MBA or CS as a supplement. We suggest that GSE supplementation may be beneficial in the primary prevention of thrombotic diseases such as myocardial and cerebral infarction.
Footnotes
Acknowledgments
We express our gratitude to Ms Chiaki Komatsu, Ms Haruka Abe, Mr Hisaichiro Nakazawa, and Mr Toshiaki Shirai for their excellent technical assistance.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by a Grant-in-Aid from the Ministry of Education, Culture, Sports, Science, and Technology, Japan.