
Abstract
Merging functional evidence derived from studies of autism spectrum disorder and attention-deficit/hyperactivity disorder converges in four neural streams of the prefrontal cortex, hence suggesting a model of information processing through four streams: motor through Brodmann area (BA) 8, emotion through BA 9, memory through BA 10, and emotional-related sensory through BA 11. A growing body of functional data has been supporting this model of information processing. Nevertheless, the underlying structural connectivity was only recently unveiled by a population-based high-definition tractography study with data from 1,065 individuals. This update provides a brief overview of recent evidence supporting the anatomofunctional integration of the four streams of the prefrontal cortex and reviews the white matter fiber tracts subserving the four streams.
Introduction
The prefrontal cortex contains Brodmann area 8 (BA 8), BA 9, BA 10, BA 11, BA 12, BA 13, BA 14, BA 24, BA 25, BA 32, BA 44, BA 45, BA 46, and BA 47, and it is thought to be involved in psychological functions such as executive tasks, attention, and memory. The current theoretical model concerns BA 8, BA 9, BA 10, and BA 11, given that they are the most rostral and might be involved in more elementary computations as compared with, say, BA 46, which is clearly involved in working memory.
This region of the brain is often treated as an undifferentiated structure, on one hand, or as a mosaic of psychological faculties, on the other. A third option is to address the landscape of the prefrontal cortex anatomically and functionally, based on the premise that structure and function are inextricably related. This model sees itself as a descendant of Goldman-Rakic’s horizontal division of the prefrontal cortex into spatial cognition (i.e., “where”), with more output-oriented areas in the dorsal prefrontal cortex and object cognition (i.e., “what”), with more input-oriented areas in the ventral prefrontal cortex.
Specifically, starting with autism spectrum disorder (ASD), Ben Shalom (2009) conceived of some core deficits in ASD in terms of four types of integration: medial BA 8, motor integration; medial BA 9, emotion integration; medial BA 10, memory integration; medial BA 11, sensory integration. Ronel (2018) conducted a similar analysis for attention-deficit/hyperactivity disorder (ADHD) in terms of four types of selection/inhibition: lateral BA 8, motor selection/inhibition; lateral BA 9, emotion selection/inhibition; lateral BA 10, memory selection/inhibition; lateral BA 11, sensory selection/inhibition. This connected to a long tradition of discussing ASD in terms of integration and ADHD in terms of selection/inhibition. An example of integration is the integration of the what, when, and where of a memory into a memory episode (event). An example of selection/inhibition is the inhibition of an undesired emotional state during the course of emotional control.
Combining these two ideas, Ben Shalom and Bonneh (2019) suggested a model of the narrow prefrontal cortex in terms of four streams of information (Figure 1): motor (BA 8), emotion (BA 9), memory (BA 10), and perception (BA 11).
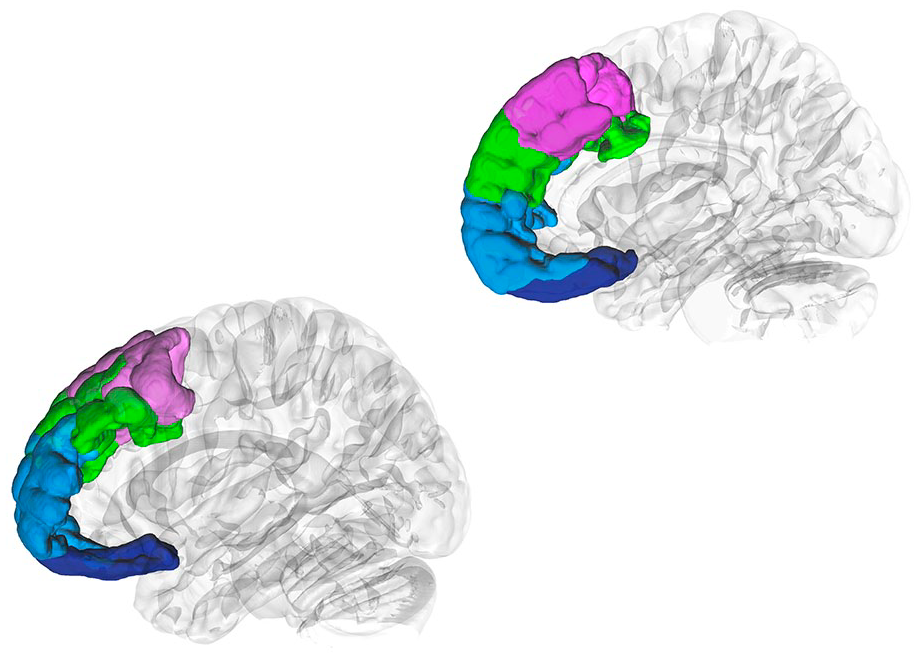
DSI Studio (Yeh 2022) was used to generate three-dimensional regions of interest of BA 8 (pink), BA 9 (green), BA 10 (light blue), and BA 11 (blue) according to the Brodmann atlas (Pijnenburg and others 2021), which were superimposed on left and right hemisphere isosurfaces. BA = Brodmann area.
A Functional Neuroanatomy of the Narrow Prefrontal Cortex
Medial BA 8 and Motor Integration
Medial BA 8 was active during action planning in work by Spiers and Maguire (2007). It was also activated in response to dots representing the joints of an agent performing an action and dots representing individual agents behaving contingently, thus supporting involvement in intentional action representation according to Chaminade and others (2011). In interesting studies, BA 8 has also been associated with decision making under uncertainty (e.g., Volz and others 2003). A plausible interpretation of this activation, however, and consistent with activations in medial BA 10 and medial BA 11, is that it reflects an interaction between prior knowledge and the current motor action—hence, the sensitivity to unexpectedness (Schiffer and others 2014). More recently, one mechanism behind this involvement was suggested to involve a rapid transformation from sensory to action space (Jonikaitis and Zhu 2023).
Lateral BA 8 and Motor Selection/Inhibition
Adelhöfer and others (2019) provided evidence that a prefrontal P2 source localized to lateral BA 8 likely reflects decision processes on action execution using stimulus information. More concretely, Beudel and de Jong (2009) showed that a comparison of free with fixed finger selection in a button press task demonstrated bilateral increases of activation on the lateral surface of the superior frontal gyrus, with a focus of maximum in lateral BA 8. Finally, one possible mechanism for the inhibition side of this selection/inhibition function tasks was a rather selective deactivation of lateral BA 8 in a conjunction of button press NoGO and count NoGO tasks (Nakata and others 2009).
Medial BA 9 and Emotion Integration
An early meta-analysis by Phan and others (2002) concluded that the medial prefrontal cortex, including medial BA 9, is the only brain area that has a general role in emotional processing—that is, showing responsiveness across individual emotions (positive, negative, happiness, fear, anger, sadness, disgust), to different induction methods (visual, auditory, recall/imagery), and in emotional tasks with and without cognitive demand. Two reasons for this involvement might have to do with Smith and Lane’s (2015) model of emotion processing, according to which stage 1 processing involves areas such as the periacqueductal gray and the hypothalamus, which can be described as encoding discrete body features; stage 2 processing involves areas such as the anterior insula, which can be described as encoding whole-body patterns; and stage 3 processing, involving areas such as medial BA 9, which can be described as encoding emotion concepts. In fact, activity in the anterior insula predicts activity in medial BA 9 (Teckentrup and others 2019), and a later meta-analysis (Kober and others 2008) concluded that mediation analyses are consistent with a pathway from medial BA 9 through the periacqueductal gray to the hypothalamus, making the former arguably important in the generation of emotional states.
Lateral BA 9 and Emotion Selection/Inhibition
Lateral BA 9 is involved in emotional control strategies: a careful recent meta-analysis concluded that lateral BA 9 is one of the areas involved in emotional reappraisal (Morawetz and others 2017); a recent small meta-analysis confirmed that it was activated in emotional reinterpretation and distraction (Liu, Wang, and others 2022). A more specific role in emotion selection/inhibition is suggested by the study of Lévesque and others (2003), which found activation in lateral BA 9 when a suppression condition (i.e., voluntarily suppress any emotional reaction in responses to sad film excerpts) was compared with a sad condition (i.e., react normally in response to comparable stimuli). Of note, BOLD signal changes in lateral BA 9 also showed positive correlations with the self-report ratings of sadness in the suppression condition, confirming its role in emotion inhibition.
Medial BA 10 and Memory Integration
Medial BA 10 has been associated with event (episodic) memory for at least two decades (Addis and others 2007), but this does not resolve its specific role within the episodic memory network. Two more recent articles provide evidence for two particular mechanisms, which might be generalizable to more areas of the medial prefrontal/medial orbitofrontal cortex. One is that the more distinct the activation pattern in medial BA 10, the better it is retained (Liu, Shi, and others 2022). The other, perhaps more important, is that in terms of bayesian decision theory, activation in medial BA 10 correlates with the mean of the posterior distribution, a statistic that reflects the integration of prior knowledge and likelihood (Ting and others 2015).
Lateral BA 10 and Memory Selection/Inhibition
One of the earliest pieces of evidence for the involvement of lateral BA 10 in memory selection/inhibition was provided by Lepage and others (2000) in a multistudy analysis of PET data. The authors found that lateral BA 10, on the left side and the right, was involved in a neurocognitive set, or state, in which one mentally holds a segment of one’s personal past in the background of focal attention and treats incoming and online information as “retrieval cues” for selecting particular events in the past. One particular mechanism for this involvement was suggested by Ranganath and others (2000), in which left lateral BA 10 was reliably activated during retrieval trials and activation in this region increased with demands to retrieve perceptually detailed information about studied objects. These notions are largely consistent with Koechlin and Summerfield’s (2007) model of cognitive control, according to which lateral BA 10 is involved in the episodic, especially branching, control of motor action, which involves switching between an ongoing and previously encoded memory episode.
Medial BA 11 and Sensory Integration
Similar to other medial prefrontal areas, there is some evidence that medial BA 11 is involved in combining prior knowledge with current information in the relevant domain. For example, Chaumon and others (2014) showed that the degree of identifiability of low spatial frequency pictures of real-world objects correlated with activity in the medial BA 11. More recently, Hervig and others (2020) found that inactivating the medial orbitofrontal cortex facilitated serial visual reversal learning in rats, which could support a notion that when inactivating the medial orbitofrontal cortex, history does not interfere with representation of current visual stimuli, thus making behavior more readily updatable. Note, however, that as part of the mammalian heritage, BA 11 has robust structural connections to the amygdala, so perhaps a better description of the representations in medial BA 11 is emotionally relevant sensory information (e.g., Tripathi and others 2021).
Lateral BA 11 and Sensory Selection/Inhibition
In general, rodent studies seem to fundamentally complement primate studies (including humans) in that they offer less cognitive sophistication on the functional side but a much better resolution on the structural side, thereby allowing for the cracking of physiologic mechanisms not clearly investigated in primates. An inspection of the human-to-macaque correspondence and macaque-to-rat correspondence in work by Preuss (1995; see Figures 4 and 7, respectively) raises the conjecture that the best human-to-rat correspondence might involve BA 8 to AC (anterior cingulate), BA 9 to PL (prelimbic), BA 10 to IL (infralimbic), and BA 11 to orbital. But a lot remains to be investigated.
Importantly, as far as we can tell, there is little doubt about the correspondence between rodent orbitofrontal cortex and BA 11. Indeed, complementing the results about medial BA 11 and sensory integration, a recent article (Banerjee and others 2020) showed sensory selection/inhibition in the lateral orbitofrontal cortex in mice in a tactile reversal learning task, in which primary somatosensory cortex (S1) activity reflected initial task learning whereas lateral orbitofrontal neurons responded saliently and tran-siently to the rule switch. Structurally, the authors identified direct long-range projections from the lateral orbitofrontal to S1 that can feed this activity back to S1 as a value prediction error. One possible mechanism for this lateral orbitofrontal involvement in updating sensory representation comes from the work of Hirokawa and others (2019), who found two key results relevant in this context. One is that, in general, lateral orbitofrontal neuron types in rats categorically encoded single decision variables—for example, decision confidence. The other and more specific one is that a connectivity-defined cell type—orbitofrontal neurons projecting to, most probably, the ventral striatum—carried a selective and temporally sustained representation of a single decision variable: integrated value. This second finding, in turn, is consistent with the results of Rothenhoefer and others (2017), who found that ventral striatal lesions—which, however, affect other reward-related circuits in addition to corticostriatal projections—caused a specific deficit in rhesus monkeys’ ability to discriminate among images with different values, as opposed to their ability to discriminate among actions with different values, thus suggesting that the lateral BA 11 has a specific role in selecting rewarded stimuli, as opposed to selecting rewarded actions.
Anatomy of the Four Streams
The model of the four streams of the prefrontal cortex proposes four streams of information: motor through BA 8, emotion through BA 9, memory through BA 10, and emotional-related sensory through BA 11. As briefly reviewed here, there is a surge of functional data supporting these four streams within the prefrontal cortex. Nevertheless, the structural connectivity underlying these neural networks had not been fully clarified by the time of suggestion of the original functional model. A recent population-based high-definition tractography study (Skandalakis and others 2023) using an averaged template generated from data of 1,065 healthy humans acquired from the Human Connectome Project sought to elucidate the structural organization of these regions. It documented the structural connectivity of BA 8 with BA 6, BA 9 with the insula, BA 10 with the hippocampus, BA 11 with the temporal pole, and BA 11 with the amygdala (Figure 2). Thus, the four streams of the prefrontal cortex are subserved by a structural neural network encompassing fibers of the anterior part of the superior longitudinal fasciculus I and II, corona radiata, cingulum, frontal aslant tract, and uncinate fasciculus.
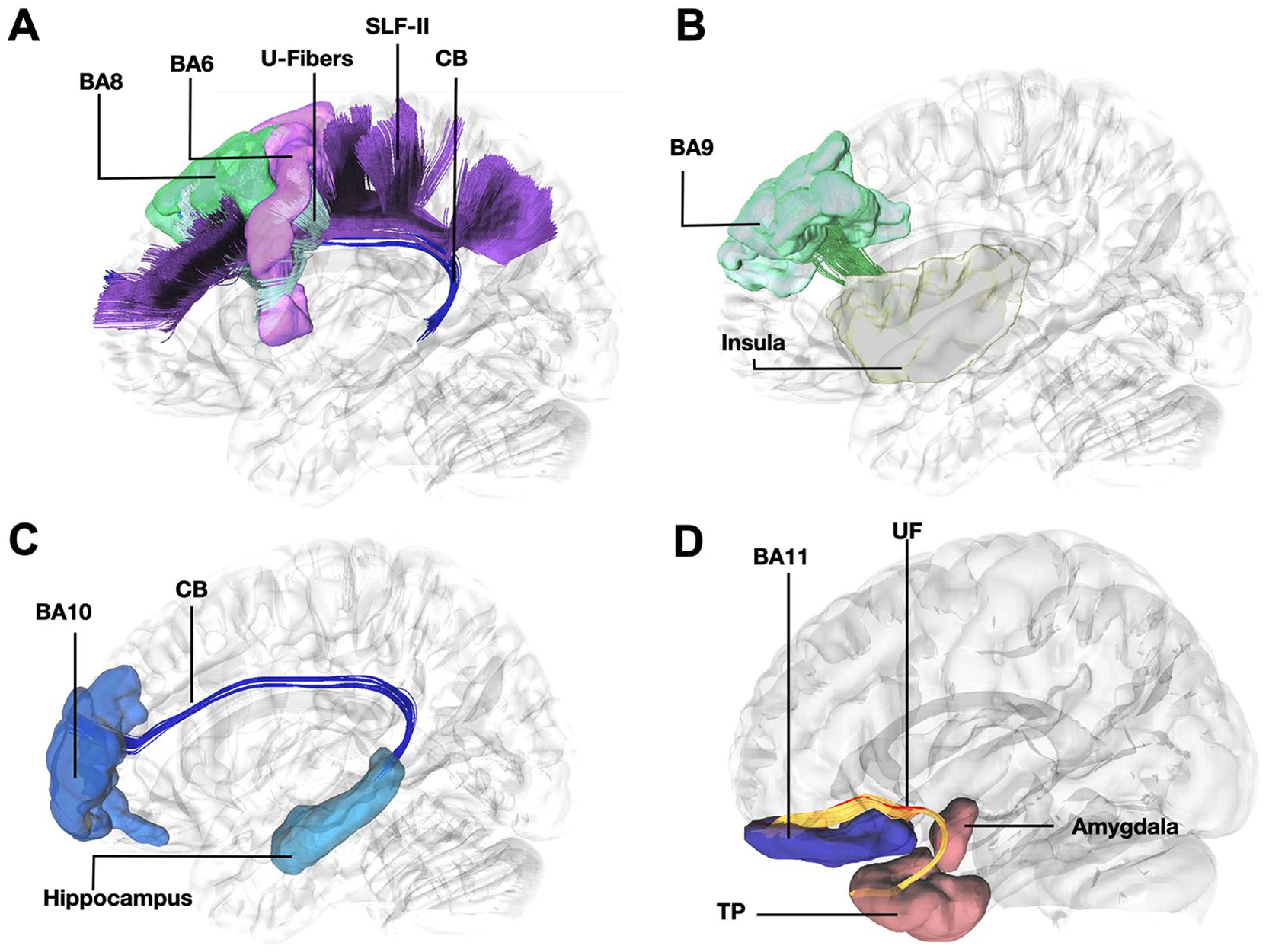
(A) Tractography demonstrating connectivity between BA8 and BA6 through U-fibers, the Frontal Aslant Tract, and the anterior part of the dorsal component of the Superior Longitudinal Fasciculus and Superior Longitudinal Fasciculus-II. light blue = anterior part of the left dorsal component of the superior longitudinal fasciculus, silver = FAT, purple = anterior part of the superior longitudinal fasciculus II purple = BA6, green = BA8, dark blue = cingulum SLF-Ia, anterior part of the left dorsal component of the superior longitudinal fasciculus; SLF-II, Superior Longitudinal Fasciculus-II; CB, cingulum bundle; BA8, Brodmann area 8; BA6, Brodmann area 6. (B) Tractography demonstrating connectivity between BA9 and insula. green = BA9, yellow = insula; BA9, Brodmann area 9. (C) Tractography demonstrating connectivity between between BA10 and hippocampus. dark blue = cingulum. light blue = dorsal hippocampus; BA10, Brodmann area 10; CB, Cingulum Bundle. (D) Tractography demonstrating connectivity of BA11 with amygdala and temporal pole through the uncinate fasciculus. red = fibers of the left uncinate fasciculus interconnecting BA11 with amygdala, yellow = fibers of the left uncinate fasciculus interconnecting BA11 with temporal pole. dark orange = amygdala and temporal pole; BA11, Brodmann area 11; UF, uncinate fasciculus; TP, temporal pole
Conclusions
This article presents functional and structural evidence for a model of the narrow prefrontal cortex (BA 8, 9, 10, and 11) in terms of four streams of information (motor, emotion, memory, and sensory, respectively). The identified functional neuroanatomy of the four streams of the prefrontal cortex will allow comprehensive analysis of these four networks in normal and pathologic brain functioning. For example, this schema might explain a high-order processing mode in normal brain function. In terms of pathologic brain function, this model might contribute to a better understanding of the neural circuits underlying behavioral changes in conditions such as ASD or ADHD. Further work, propelled by functional imaging and related techniques, is likely to build on this new understanding of differential information streaming within the prefrontal cortex.
Footnotes
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.