
Abstract
We present a robot architecture and experiments to investigate some of the roles that pleasure plays in the decision making (action selection) process of an autonomous robot that must survive in its environment. We have conducted three sets of experiments to assess the effect of different types of pleasure—related versus unrelated to the satisfaction of physiological needs—under different environmental circumstances. Our results indicate that pleasure, including pleasure unrelated to need satisfaction, has value for homeostatic management in terms of improved viability and increased flexibility in adaptive behavior.
Keywords
1 Introduction
We present a biologically-inspired robot architecture and experiments to investigate some of the roles that pleasure plays in the decision making or action selection (AS) process of a motivationally autonomous robot that must survive or remain ‘viable’ (Ashby, 1960) in its environment. This study is framed by our long-term interest in the interactions between emotion, motivation and cognition from an embodied perspective, and is a first step towards a more systematic study of the roles of pleasure in such interactions.
Many definitions of pleasure have been provided in the literature, reflecting the fact of its multi-faceted nature, its multiple meanings, and its multiple underlying biological mechanisms, to the extent that some authors talk about ‘pleasures’ rather than ‘pleasure’ as a single notion (Frijda, 2010; Kringelbach & Berridge, 2010), e.g. sensory pleasure, non-sensory linkings, pleasures of achievement, pleasures of gain and relief, social pleasure, activity pleasures, esthetic pleasures. Underlying the different views, however, we can identify a common element of ‘positive affect’, of enjoyment, which is also found in lay, non-technical definitions (e.g. in dictionaries such as the Merriam–Webster or the Oxford). In this paper, we take such a broad, common-sense definition of pleasure as ‘liking’, focusing on two different contexts in which such ‘liking’ can take place: as linked to the satisfaction of survival-related physiological needs, and as a purely hedonic quality not directly linked to need satisfaction. In both cases, we have focused our study on the influence that pleasure has on the perception of external stimuli, and the underlying mechanism we have adopted to model pleasure is a simulation of hormonal modulation of perception. Unlike related work in robotics (Krichmar, 2008, 2012, 2013; Sporns & Alexander, 2002), our simulated hormone constitutes an abstract model aimed to capture the (gross) dynamics of modulation rather than modeling the behavior of specific chemicals underlying pleasure and more generally affective phenomena. Hormonal modulation has been used in robots for other purposes, such as behavior control (Moioli, Vargas, & Husbands, 2009), learning based on value and reward systems (e.g. the 2013 special issue of Frontiers in Neurorobotics on this topic; see (Krichmar and Röhrbein, 2013) for a review of the papers), to model intrinsic motivation (Kaplan & Oudeyer, 2007; Luciw, Kompella, Kazerounian, & Schmidhuber, 2013), energetic autonomy (Lowe et al., 2010; Sauzé & Neal, 2010), and evolutionary and modular robotics (Hamann, Stradner, Schmickl, & Crailsheim, 2010).
Although pleasure is intimately related to affect, and more specifically to emotion, the nature of this relationship remains elusive. Some consider pleasure as an emotion—e.g. one of the earliest forms of emotion that evolved (Panksepp, 1998)—others think that pleasure, even though at the origin of emotions, is not an emotion itself but ‘a constituent quality of certain emotions as well as a trigger for certain emotions’ (Damasio, 1999, p. 76). Firmly grounded in the body’s biological ‘machinery’ at different levels, the term ‘pleasure’ is normally used to denote a subjective quality: a positive hedonic quality, an affective sensation or feeling of pleasantness. To some, pleasure has both sensory and affective elements (Nafe, 1924), whereas for most, pleasure is the affective component (Cabanac, 2010; Kringelbach, 2010), a ‘gloss’ (Frijda, 2010) of sensory processing.
Pleasure is often associated with positive valence. However, the interactions between hedonic feelings, positive affect, and approach behavior can be very complex (Leknes & Tracy, 2010), and their underlying mechanisms are not clear. This link is often explained in terms of the biological ‘utility’ (e.g. adaptive value, evolutionary usefulness) of pleasure, which, from this perspective, would signal that stimuli are beneficial (Cabanac, 1971, 1979; Panksepp, 1998) and foster the acceptance of such stimuli (Frijda, 1986). This view is particularly compatible with models of the body in terms of homeostasis (Leknes & Tracy, 2010), as well as with models of learning inspired by classic and operant conditioning (Rolls, 2014), and is reflected by their focus on the ‘reward’ aspect of pleasure to the neglect of other aspects. This is also the predominant interest of robotic models that take into account pleasure, mostly in the context of reinforcement learning (e.g. Cos, Cañamero, Hayes, & Gillies, 2013; Gadanho & Hallam, 2001; Hiolle & Cañamero, 2009; Kitano, 1995). In previous work, we have investigated the role of pleasure as reward in the maintenance of homeostasis in a reinforcement learning model (Cos et al., 2013). In this paper, we depart from the idea that pleasure is necessarily linked with reward—in the same way as value is not necessarily linked with reward (Krichmar & Röhrbein, 2013)—or with signaling biological usefulness, opening the door to the investigation of the role of other types of pleasure not directly related with the satisfaction of needs (Frijda, 2010), in addition to pleasure stemming from need satisfaction.
Thinking about the link between pleasure and valence leads us to a key unresolved question in affective neuroscience and psychology: how do we go from ‘liking’ something to ‘wanting’ it? This link is, once more, often conceptualized in terms of ‘usefulness’ and ‘reward’ and, in the latter case, in the context of learning. Again, in this paper we depart from this view, in two respects. First, we think that hedonic quality (just ‘liking’, ‘pure pleasure’ unrelated to need satisfaction) might also have an important role in preference behavior and motivation (Frijda, 1986, 2010; Young, 1961). Second, given the involvement of pleasure in basic life regulation in line with its early evolutionary origins (Damasio, 1999; Panksepp, 1998), we think that, to better understand pleasure, we need to consider its roles in the context of ‘simpler’ cognitive functions, and notably in the context of perception, since pleasure involves closely related hedonic and sensory aspects. We have thus investigated the effect of pleasure on the perception of external stimuli and, more specifically, of the stimuli relevant to the satisfaction of homeostatically-controlled internal needs. As we will see, in our model, the modulation of motivation-related perception through pleasure provides a link between ‘liking’ and ‘wanting’, as it changes the ‘attentional effort’ (Sarter, Gehring, & Kozak, 2006) or ‘incentive salience’ (Berridge & Robinson, 1998) of the stimuli. As Pessoa (2013, p. 190) puts it: ‘At the perceptual level, items with affective/motivational content act as if they had increased salience, which improves performance if they are task relevant but impairs it if they are task irrelevant.’
In previous work (Cañamero & Avila-García, 2007), we had used hormonal modulation of the perceptual element of motivation as a function of increased internal deficits and the presence of threats in the environment, i.e. hormone was released signaling that things were not functioning well. In this paper, a simulated modulatory pleasure hormone is released as a function of need satisfaction, signaling an improvement in the interaction with the environment and fosters ‘openness’—increasing the interaction with the environment or continuing an interaction that is going well. We thus follow the principle that pleasure signals well functioning (Panksepp, 1998).
To facilitate systematic analysis of experimental results, we have tested our model of pleasure using what in the AS literature is known as a ‘two-resource problem’ (2RP) (Spier & McFarland, 1997), implementing the simplest decision-making scenario. As its name suggests, in this scenario, an agent (animal or robot) must autonomously decide which of the two resources available in the environment it should consume in a timely fashion in order to satisfy its two survival-related needs successfully. The experiments reported here aim to compare the viability and behavior of robots whose pleasure hormones, released under different circumstances, play different roles. We compare the effects of pleasure that varies as a function of need satisfaction with the effects of pleasure as pure hedonic quality, unrelated to need satisfaction—either independent or added to it. In all cases, the pleasure hormone acts on the ‘subjective assessment’ or ‘assignment of value’ to the perceived stimuli, modifying their incentive salience.
2 Robot’s action selection architecture
Our robot’s architecture (Figure 1) follows design principles of embodied artificial intelligence (Brooks, 1991; Pfeifer, Iida, & Bongard, 2005; Steels, 1993), and builds on our longstanding approach (Cañamero, 1997, 2001) to ground embodied cognition and interaction in ‘core’ affect modeled around a ‘physiology’ of homeostatically-controlled essential variables. For this study, we have used the humanoid robot Nao, since we have developed our pleasure model with the intention to implement it in the autonomous social robot toddler Robin based on a Nao robot, (www.emotion-modeling. info/robin) (Cañamero & Lewis, 2016; Lewis & Cañamero, 2014) that we started developing as part of the EU project ALIZ-E.
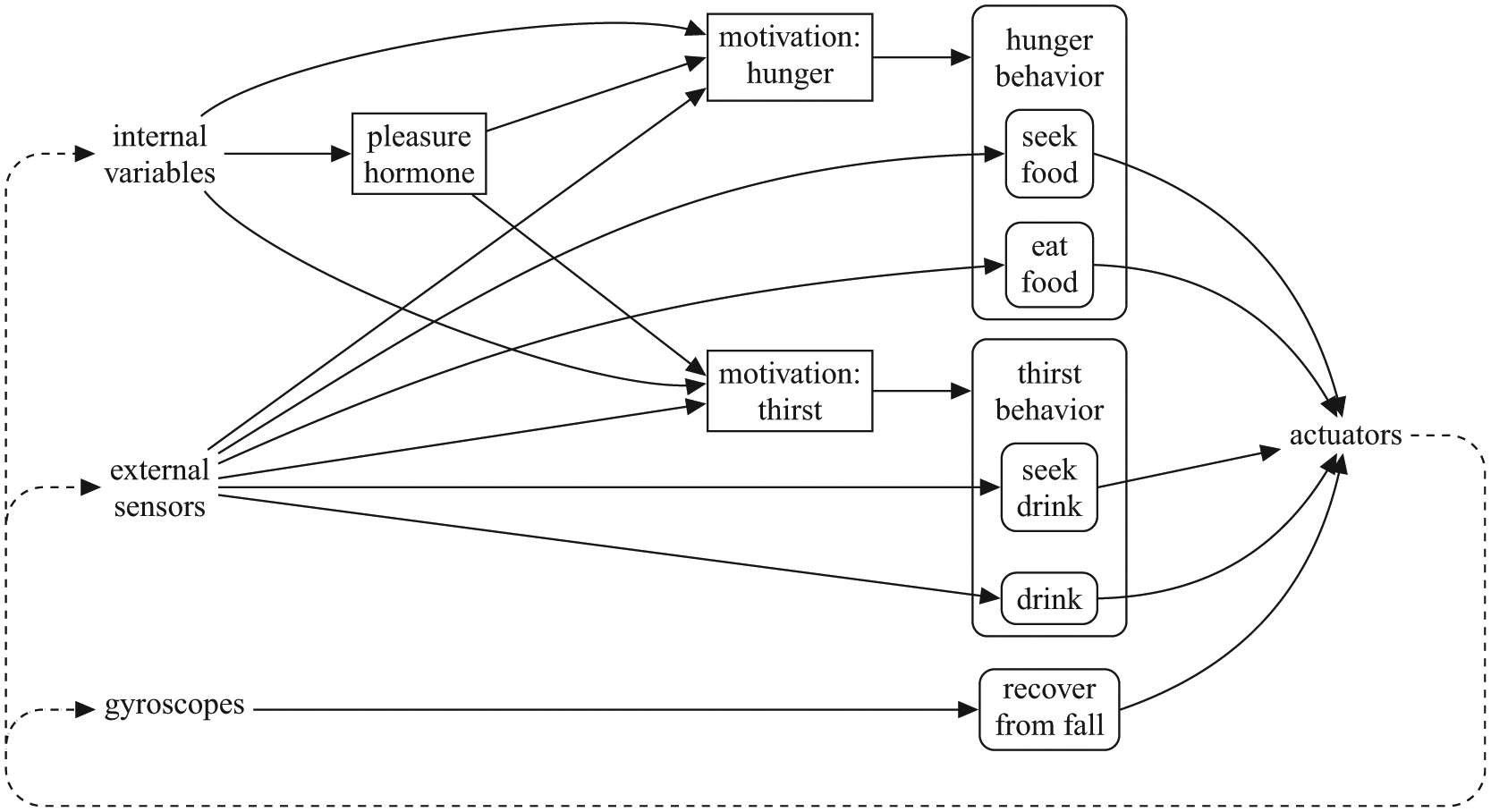
The AS architecture of our robot. Rounded boxes represent behaviors, square boxes represent other internal elements. Composite behaviors are shown in compact form. Some behaviors (‘return-to-neutral’) are omitted to avoid cluttering the diagram.
As a basis for our architecture, we developed a framework in UrbiScript (Baillie, Demaille, Hocquet, Nottale & Tardieu, 2008) consisting of base classes for behaviors, and functions to execute (either synchronously or asynchronously, as required) one or more behaviors of different types, e.g. with different levels of granularity and structure, different constraints, and different durations and temporal dynamics. 1
2.1 Physiology
To implement a simple 2RP, we have given our robot two essential variables, symmetrical in their range of values and dynamics, that need to be kept within permissible limits—lower limit of 0, upper limit of 100—for the robot to remain viable, i.e. survive: energy (replenished by consuming ‘food’) and hydration (replenished by consuming ‘drink’). They are controlled homeostatically, mostly through interaction with the world when behaviors are executed. They have an ideal value or setpoint (in this case coinciding with the upper limit) and the homeostatic control seeks to bring their actual value as close as possible to the ideal value. The difference between ideal and actual values of the variables gives deficit errors (‘deficits’ for short) that provide the robot with internal—and in this case survival-related—needs that its behavior will try to address. Both deficits increase by 0.1 every 250 ms, so if each deficit starts at 20, the robot will die after 3 min 20 s if it does not succeed in finding and consuming both food and drink before this time.
2.2 Perception and actuation
Our robot interacts with its environment constantly. In terms of actuators, it uses its legs to walk, moves its head to visually detect and track objects, and its hand to reach and ‘consume’ food and drink. While interacting, it continuously and asynchronously monitors the environment for percepts potentially relevant to the satisfaction of its needs. We use the standard sensors of Nao, as follows:
one on-board camera to detect the resources (colored plastic balls) on the grounds of their color and size;
sonars to detect obstacles;
contact sensors in the feet to detect collisions;
gyroscopes to detect inclination of the body and falls.
Food (red) and drink (green) resources (plastic balls) are consumed in discrete chunks (‘bites’), each decreasing the corresponding deficit by 10 units. The resources are not depleted when consumed, and therefore each one is a potentially infinite reservoir.
2.3 Motivations
Motivations in animals (including humans) can be defined as ‘inferred internal states postulated to explain the variability of behavioral responses’ (Kandel, Schwartz, & Jessell, 1995, p. 614). In AS, they are modeled as functions that combine the perception of internal deficits and the perception of relevant elements of the environment to provide the robot with urges to action—‘wanting’ to do things—in order to satisfy its needs in the environment in which it is situated. In addition to its longstanding use in the adaptive behavior community (Avila-García & Cañamero, 2005; Cañamero, 1997; Cos, Cañamero, & Hayes, 2010; McFarland & Spier, 1997; Tyrrell, 1993), this notion is broadly used in various disciplines that inform our work, such as animal behavior (Colgan, 1989; Hinde, 1960; Toates, 1995), neuroscience (Kandel et al., 1995; Panksepp, 1998; Pessoa, 2013; Robbins & Everitt, 1999), and psychology (Dai & Sternberg, 2004; Elliot, 2008). In our simple 2RP, each physiological variable has a single motivation associated with it.
As part of the action AS process, motivations are assigned intensity or activation levels that indicate how ‘relevant’ they are, given the robot’s needs and the current external perceptions, as follows
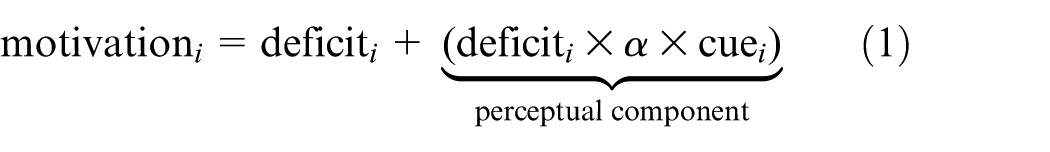
where
As we will see in Section 2.5 and through our experiments, our pleasure mechanism acts on motivations, and hence on the AS process, through modulation of the
2.4 Behaviors
Our architecture has four main or ‘top level’ behaviors: a reflex-like behavior to recover from falls; a ‘return to normal’ behavior that makes the robot adopt a neutral posture when it is not engaged in other activities; and two behavioral subsystems or ‘composite behaviors’ related to the motivations of the robot—hunger and thirst—and that inherit their intensity or activation levels. 2
These motivated behavioral subsystems are composed of a number of smaller behaviors and they are thus ‘action selectors’ themselves, both conceptually and, in our implementation, in an object-oriented sense (Figure 2). The smaller behaviors can, in turn, be composed of yet smaller behaviors, and thus can also be ‘action selectors’. These smaller behaviors can be of two types, as shown in Figure 1—consummatory or appetitive—following the traditional distinction in ethology, neuroscience and AS (Blumberg, 1994; Hinde, 1953; Maes, 1991; McFarland & Spier, 1997; Robbins & Everitt, 1999; Tyrrell, 1993). Consummatory behaviors are goal-achieving and need the presence of a specific incentive stimulus to be executed. Appetitive behaviors are goal-directed search for (or avoidance of) a particular incentive stimulus. In addition to modifying the external environment, the execution of a consummatory behavior has an impact on the level of specific physiological variables; therefore, they are a mechanism to keep the physiological variables viable.
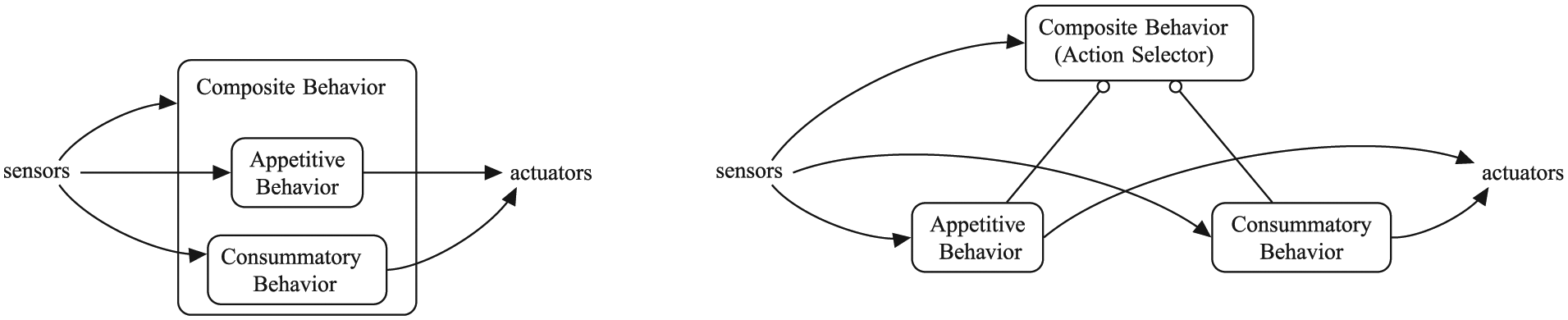
A composite behavior comprising of simpler sub-behaviors—a typical ‘branch’ in our behavior tree. The compact representation, left, as used in Figure 1, is expanded into its tree form, right, which shows the role of the composite behavior as an action selector.
As an example, the hunger behavioral subsystem consists of two immediate sub-behaviors: the consummatory eat food and the appetitive seek food. The latter is in turn composed of four sub-behaviors.
Wander: to walk around the arena when no resource has been detected. This is composed of four sub-behaviors and the interaction of these behaviors, which depend on input from the sensors (e.g. sonars and foot sensors) as well as some random elements, determines the path of the robot.
Visual search: to move the head from side to side. It is continually active, but it is ‘interrupted’ when a food object is detected (when the gaze behavior, which also uses the head, becomes active).
Gaze: at a resource, to turn the head so that the detected food/drink resource that is largest in the camera’s field of view (typically the nearest resource) falls in the center of the visual field.
Approach: to walk towards whatever the robot is facing, turning the body in the appropriate direction. Whenever the approach behavior is active, the gaze behavior will also be active, and the combination of these two behaviors will result in the robot walking towards resources that are detected by the camera.
Each behavior has an activation threshold that determines the level of activation that must be reached for the behavior to be executable, following a process described in Section 2.6. As previously mentioned, multiple behaviors can be executed simultaneously if they do not use the same actuator in a way that makes their simultaneous execution incompatible.
2.5 Modeling pleasure
In the experiments presented in this paper, we have used three ‘types’ of pleasure, all of which act on the perceptual element of motivations (the
Pleasure modeled by a hormone released as a function of the satisfaction of homeostatic needs, building on Cañamero (1997) and Lewis, Hiolle, and Cañamero (2014), that then decays to a background level. This is the model that we describe in this section, and which will be referred to as ‘Modulated
Different fixed values of ‘hormone’ (which we will refer to as ‘fixed values of
Additional hormone (a constant amount) is released linked to the execution of consummatory behaviors of ‘eating’ or ‘drinking’ in Experiment 3 (Section 6). This additional release is unrelated to the satisfaction of needs, and corresponds to the purely hedonic pleasure mentioned in Section 1. We will refer to this as ‘additional release’ or additional pleasure in the experiments (Section 6).
The rest of this section refers exclusively to the first type of pleasure above. In this context, instantaneous changes in the internal homeostatic variables (mathematically, their first derivative) can be thought of as indicators of the current interactions of the agent with the environment. Our hormonal system reflects improvements (thought of as pleasure) or deterioration (thought of as displeasure) in the interaction with the environment, which we model using the second derivative of the deficit of each homeostatic variable. For the sake of making our investigation incremental, in these experiments we only take into consideration pleasure, i.e. improvements in the interaction with the environment. Since we want our hormone to be released when there is a change resulting in a ‘better’ interaction with the environment, we link hormone release to the negative second derivative of any of the homeostatic deficits. 3 For example, if the robot starts consuming food, the energy deficit decreases (resulting in a negative first derivative); since it was previously not consuming food, the second derivative of the energy deficit is also negative, and hence pleasure hormone is released.
Specifically, our model is as follows. On a 500 ms cycle (empirically determined), we store a history of each deficit’s recent past values

and ‘second derivatives’ as

If the second derivative is negative, then the level of the hormone is increased by

where s is a scaling parameter. Because our resource items reduce their respective homeostatic deficits by 10, the value of the
The decay of the hormone to a background level is implemented by updating the hormone level h once per second according to the rule

where b is the background level and k the decay rate. The background level corresponds to a state of equilibrium between the release of the hormone and its natural decay. We use values
In summary, the dynamics of the hormone, illustrated in Figure 3, depends on three factors:
its background level, b;
its decay rate, k;
the relationship between homeostatic variables and hormone release—a linear scaling by s of the second derivative.
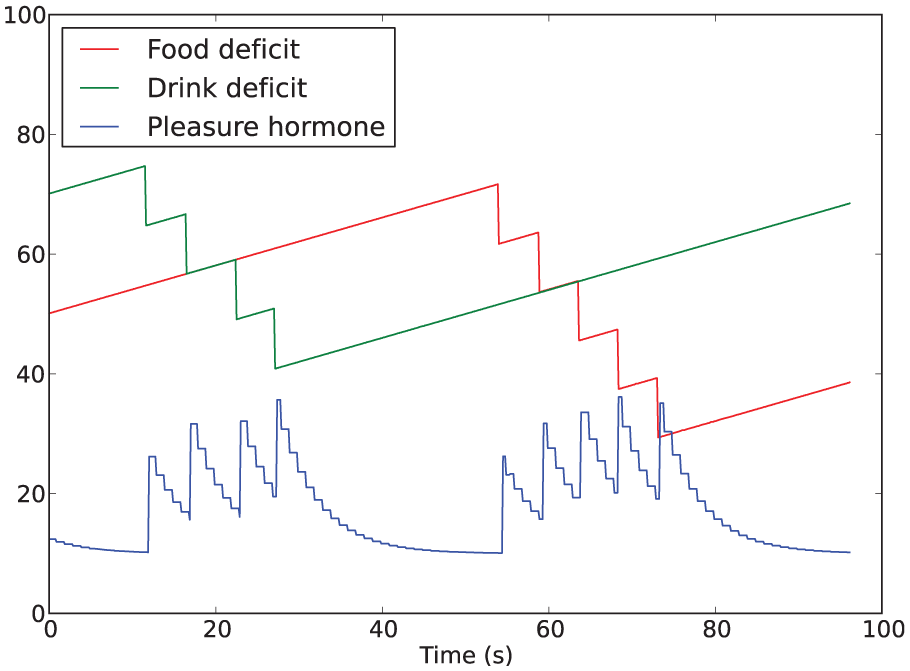
Example of the hormone being released with decreases in the homeostatic deficits and then decaying to the background level.
In those experimental conditions where the hormone level affects
2.6 Action selection process
The AS process involves various elements running on different time scales. Each AS ‘cycle’ is thus not a sequential loop, but a number of asynchronous loops running in parallel. The main elements can be grouped as follows.
The robot is continuously ‘monitoring’ its environment for percepts that might be relevant to need satisfaction and its interaction with the world. Different sampling rates are used for different sensors as appropriate.
The physiology (i.e. the values of the internal variables and the pleasure hormone) is updated.
Motivational intensities are calculated and passed on to the top-level behavioral subsystems.
A behavior selection cycle selects the behavior(s) to be executed every 125 ms, i.e. 8 times per second. This cycle is a sequential loop that can be summarized as follows. Each behavior, starting with the top-level behavioral subsystems: gets the activation levels of its sub-behaviors; sorts the sub-behaviors according to their activation levels, highest to lowest; for each sub-behavior in its sorted list, this behavior (in order): – checks if it is inactive (i.e. if its activation level is below its activation threshold). If so, it skips to the next sub-behavior; if not: – checks if any of the actuators needed for the sub-behavior are already in use by ‘extended behaviors’.
4
If so, it skips to the next sub-behavior; if not: – checks if any of the actuators needed for the sub-behavior have already been used by behaviors already executed in this cycle. If so, it skips to the next sub-behavior; if not: – selects the sub-behavior (which may itself be an action-selector), which is executed. If this sub-behavior is an extended behavior, then it tags those actuators it is using as ‘in-use’.
5
The extended behavior then spawns a separate thread in which the main part of its execution occurs. – after it has been executed, this sub-behavior returns the actuators that it used to the parent behavior, which adds them to the list of actuators already used in this cycle.
3 Experiments’ method and metrics
3.1 Method
To facilitate systematic analysis of results, we decided to test our model of pleasure using a 2RP (Spier and McFarland, 1997). Even this simple problem can give rise to a number of variations with potential consequences for the viability and decision-making behavior of the robot. For this study, we have manipulated the following: the availability of resources (easy/difficult access), their (symmetric or asymmetric) distribution, and how the release of pleasure relates to their consumption—either to their ‘nutritional value’ or simply to the act of consuming.
The arena used to design the environments of all our experiments is a 2
3.2 Metrics
We use the following metrics to assess the robot’s performance and to characterize relevant aspects of its behavior.
3.2.1 Viability indicators: Comfort and discomfort
We use indicators of performance based on the notions of viability and ‘wellbeing’ (Ashby, 1960; Avila-García & Cañamero, 2004) to assess different aspects of how the viability of the physiology is maintained in the interactions of the robot with its environment.
Unlike in previous work, e.g. (Avila-García and Cañamero 2004; and Cos et al., 2013), which builds on the notion of ‘comfort’, here we use the converse notion of ‘discomfort’ that increases and decreases following the deficits (rather than inversely to them), and hence their link between the metrics and the deficits is intuitively easier to see. In addition to using the arithmetic mean of the deficits at time t and the variance of the deficits at time t as measures of ‘overall discomfort’, we introduce a measure of discomfort based on the geometric mean. 6 We define arithmetic and geometric discomfort (and comfort) as:
The arithmetic discomfort at time t,
The geometric comfort at time t,
We prefer to use the geometric (rather than the arithmetic) discomfort, as it has the advantage that the discomfort is at its maximum value (100) if and only if the agent is dead, whereas the arithmetic discomfort would make possible the counter-intuitive situation where a dead agent could have less discomfort than a living agent. In addition, from the well-known relationships of the geometric and arithmetic means, we see that
3.2.2 Behavioral metrics: Persistence and opportunism
These metrics take into account two of the key problems that AS architectures should be able to tackle: persistence and opportunism (Maes, 1995; Tyrrell, 1993). Generally speaking, in the AS literature, persistence is related to the ability of an agent to continue working towards its most relevant ‘goals’, while opportunism has to do with its ability to take advantage of relevant opportunities and contingencies offered by the environment. For an autonomous robot that has to satisfy multiple conflicting survival-related needs, it is crucial not only to choose behaviors that do so in a timely fashion, but also to persist in their execution for long enough to guarantee sufficient satisfaction. Persistence is important to avoid what is known as the ‘dithering’ problem, which occurs when a robot keeps switching between trying to satisfy two needs without satisfying any of them enough to guarantee survival. Closely related to persistence, opportunism is the consumption of a resource that might not be needed at present but is available now and might not be available later. The degree to which a robot should show persistence and opportunism depends on multiple factors; we could generally say that persistence leads to a more ‘conservative’ AS behavior and opportunism to a more ‘risky’ one. In previous work (Avila-García & Cañamero, 2004; Lewis et al., 2014), we showed that persistence and opportunism can also become negative when done in excess, and proposed initial mechanisms inspired from emotions in natural systems and based on hormonal modulation of the perception of external stimuli (the resources), to address these problems.
We first consider definitions of persistence and opportunism that we call ‘persistent’ and ‘opportunistic’periods ofconsumption to distinguish them from our later definitions of periods of attempted persistence and opportunism.
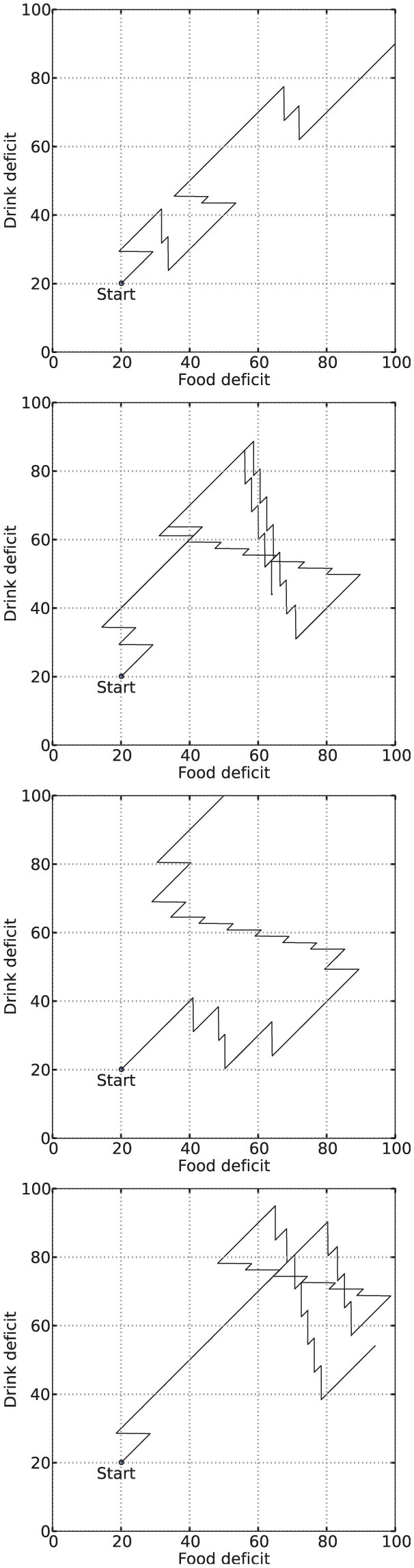
We define a period of persistent consumption as a period where the consumption of a resource that had started when its corresponding deficit was the largest, has continued beyond the crossover point in the ‘deficit’ or ‘physiological’ space (Figure 4). The crossover point is the point at which the largest deficit falls below another deficit and is no longer the largest.
We define a period of opportunistic consumption as a period of consumption that started when the corresponding deficit was not the largest or equal to the largest.
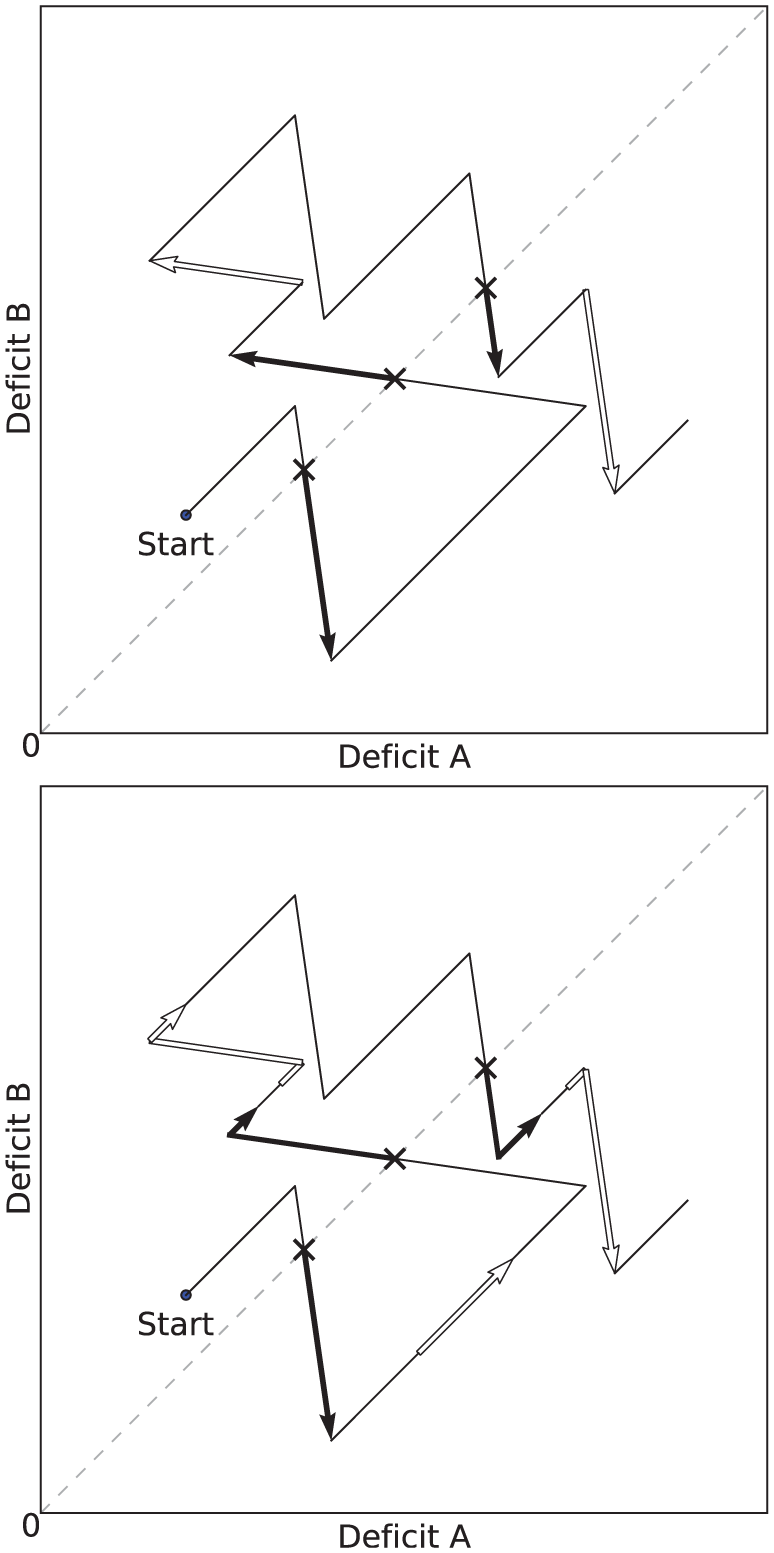
Example plots in a two-resource ‘physiological’ or ‘deficit’ space. The arrows mark periods of persistent/opportunistic consumption (top), and attempted persistence/opportunism (bottom). Both plots show the same evolution of the deficits. From the starting point, the deficits increase as the robot is not consuming any resource. Shortly after starting, the robot begins to consume a resource which decreases Deficit B (the most pressing homeostatic need at this point), while Deficit A continues to increase. During consumption, a first crossover point is reached as both deficits reach the same level (crossover points are marked by a cross), and subsequently a first period of persistence starts. Top: Periods of persistent consumption are shown as filled arrows, and periods of opportunistic consumption by unfilled arrows. Bottom: Periods of attempted persistence are shown as filled arrows, and periods of attempted opportunism by unfilled arrows. In this plot we see that during the long period where both deficits were increasing, there is a period of attempted opportunism—the agent attempted to reduce Deficit B, but it failed, leading to a costly increase in both deficits, and delaying the consumption of a resource to reduce Deficit A.
These definitions of persistence and opportunism only take into account occurrences when resources are successfully consumed. However, one reason for considering persistence and opportunism is that they should occur in a way that balances their possible benefits (consumption of resources) against their costs (time that could otherwise be used seeking resources to satisfy more pressing deficits). Our previous definitions of persistent and opportunistic consumption only include periods where the benefits (successful consumption) are gained, and not periods involving cost (time wasted attempting but failing to access a resource). To take into account these latter periods, we introduce the notions of attempted persistence and opportunism (Figure 4, bottom).
We define a period of attempted opportunism to be one in which all of the following hold: Any of the (consummatory or appetitive) behaviors associated with a resource R are being executed. The deficit associated with resource R is not the largest (or equal largest) of the deficits. Immediately prior to the period, none of the behaviors from (1) were active.
We similarly define a period of attempted persistence to be one in which (1) and (2) above hold, and in addition: 3. Immediately prior to the period, at least one of the behaviors from (1) was active, and the associated deficit was the largest (or equal largest) of the deficits.
4 Experiment 1: Comfortable environment
The first set of experiments were carried out in a simple, unchallenging ‘baseline’ environment in which resources are plentiful, equally distributed and easily accessible. The robot should be able to satisfy its needs so as to normally survive for the entire duration of the runs. In this experiment we are not assessing the role of pleasure in terms of its utility for survival, but rather whether pleasure makes a difference in terms of (a) how viability or wellbeing is maintained and (b) the type of behavior exhibited by the robot.
4.1 Experimental setup
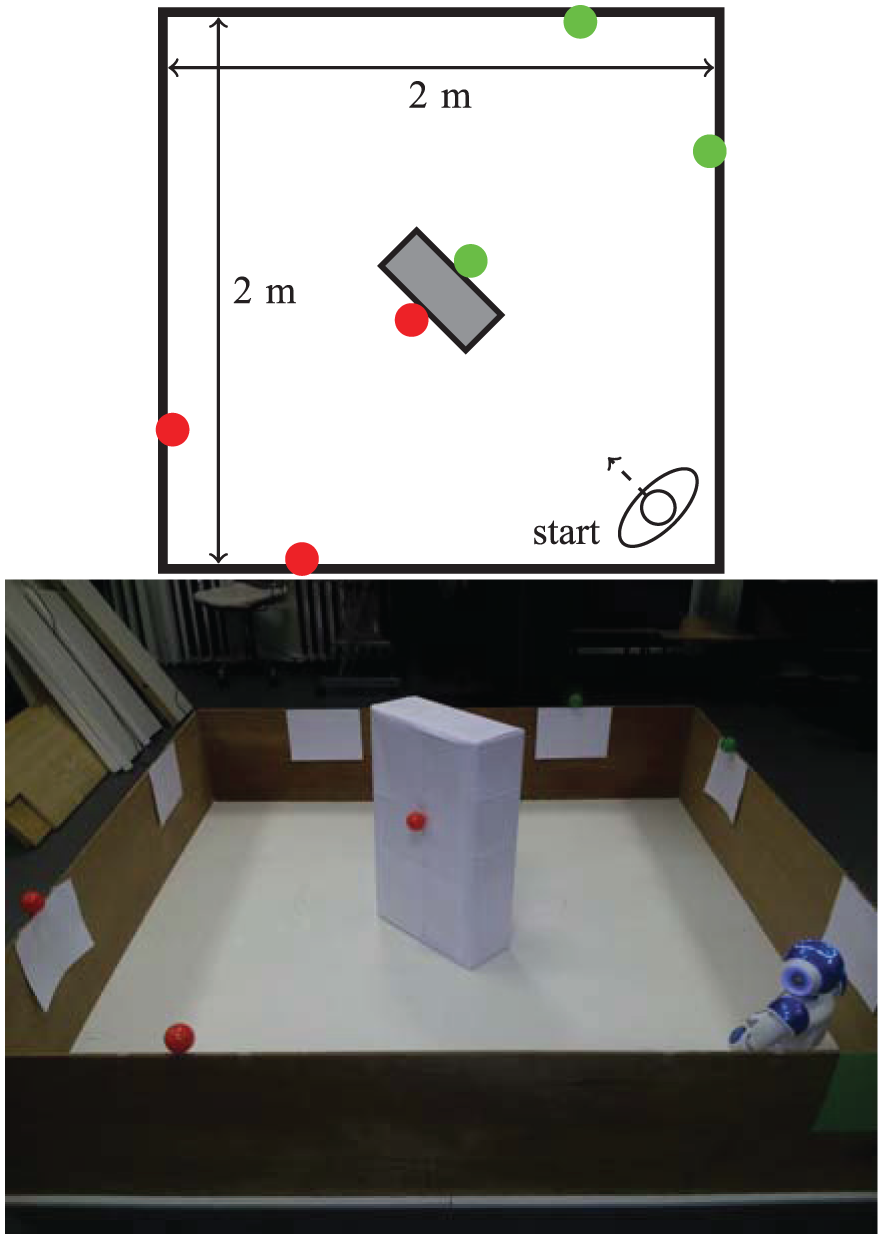
Four red (‘food’) resources and four green (‘drink’) resources are fixed to the top of the walls of the arena, where they are easily visible to the robot’s camera, as shown in Figure 5. This environment is symmetric in the following ways.
The number of items of each resource is identical.
The ‘nutritional value’ of both resources is identical.
The metabolism and physiological changes (e.g. rates of growth and satiation of the deficits, ideal value and fatal limits of the variables) associated with both resources are identical.
The conditions tested (Section 4.2) were identical for each resource. More specifically, the values of
The resources are symmetrically distributed in the environment (Figure 5) as follows: reflection in one diagonal (the diagonal along which the robot is facing at the beginning of each run) would result in the resources swapping (complementary symmetry: red ↔ green); reflection in the other diagonal would leave the resources unchanged (mirror symmetry: red → red, green → green).
The starting position of the robot (in the middle of the arena) affords an equal view of the two resources.
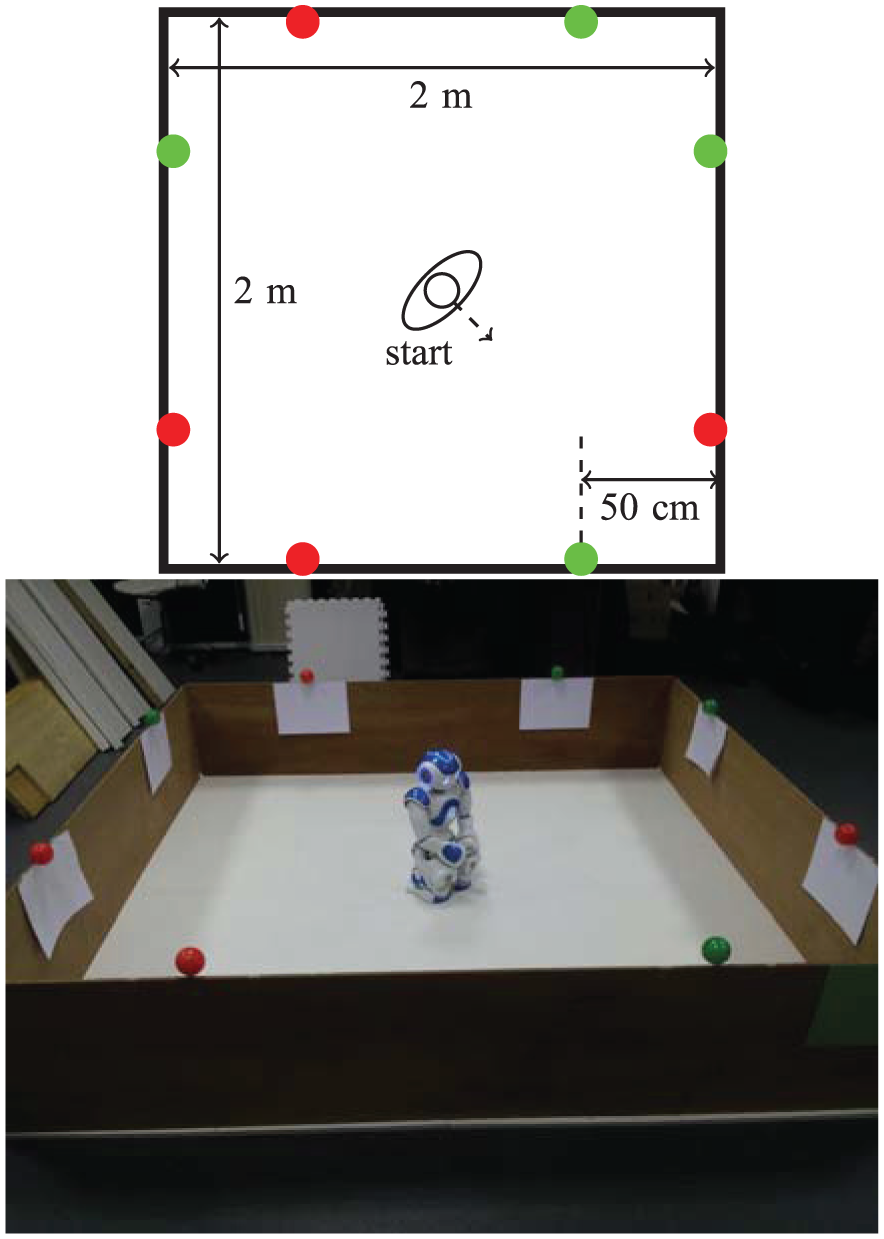
A diagram and photo of the arena used for Experiment 1, showing the robot’s starting position and the location of the resources.
4.2 Experimental conditions tested
We compared four different
Condition 1, ‘Low Fixed
Condition 2, ‘Medium Fixed
Condition 3, ‘High Fixed
Condition 4, ‘Modulated

The particular value 0.01 was chosen so that
The specific LF
We conducted a total of 40 runs—10 runs for each condition. The order of runs was randomized, with runs in each condition spread across the set of runs. This was done by generating 10 random orderings of the numbers 1 to 4. All runs were done in the same artificial light conditions. To avoid potential differences in the functioning of sensors and actuators, breaks (of varying length depending on practicalities, but of at least 10 min) were taken between runs to recharge the battery and to allow the joint motors to cool down. On each run, the robot started at the center of the arena, facing the same corner. Each run lasted either until the robot ‘died’ (one deficit passed the fatal limit), or until 6 min had passed. The data recorded during each run comprise: the values of the deficits, the motivations, the hormone level, the resources detected, and the currently active behaviors.
4.3 Results
Figure 6 shows example deficit–space plots from single runs under each condition. As we can see, increasing the value of
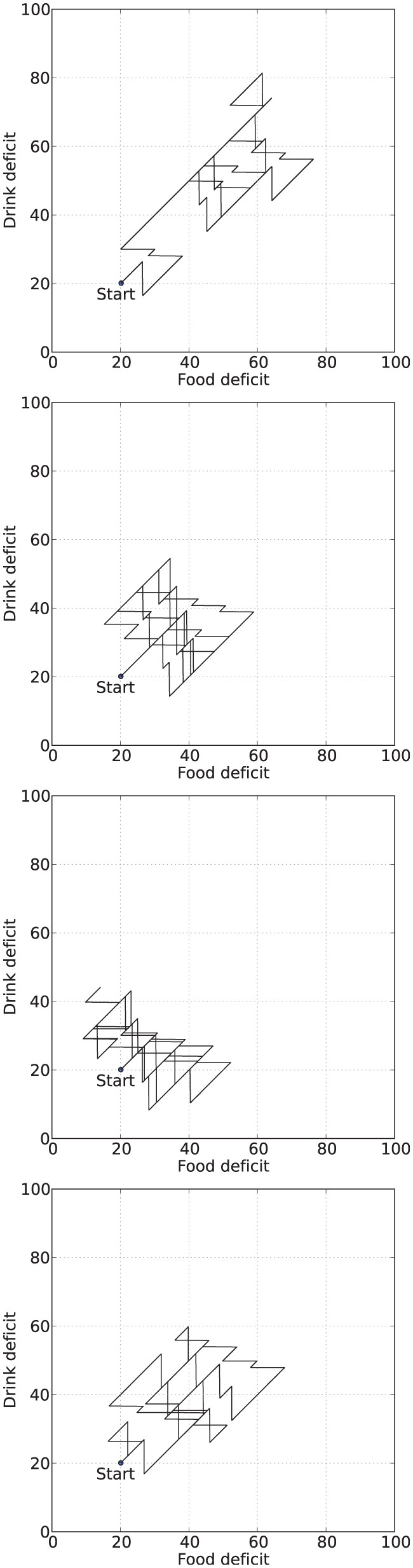
Example deficit–space plots from Experiment 1. From top to bottom:
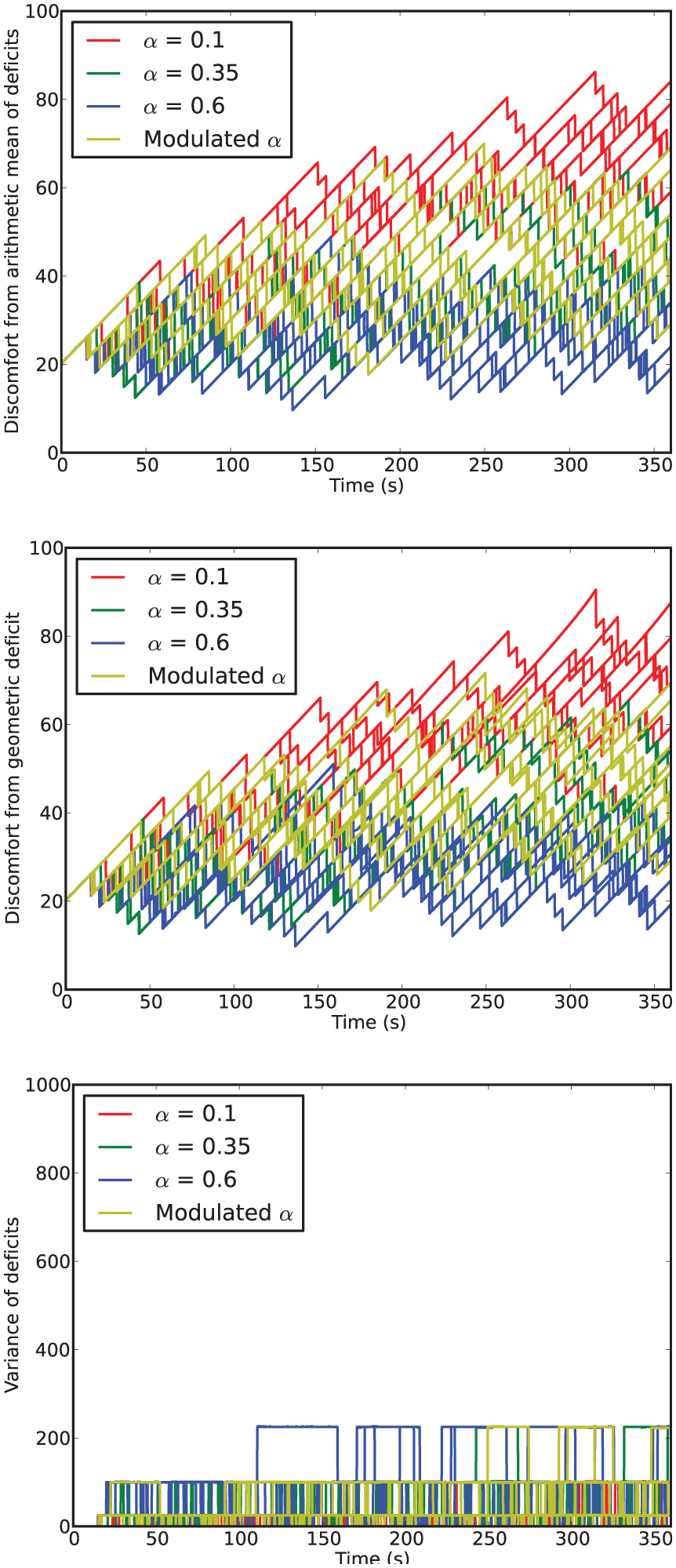
Time series of discomfort metrics from Experiment 1.
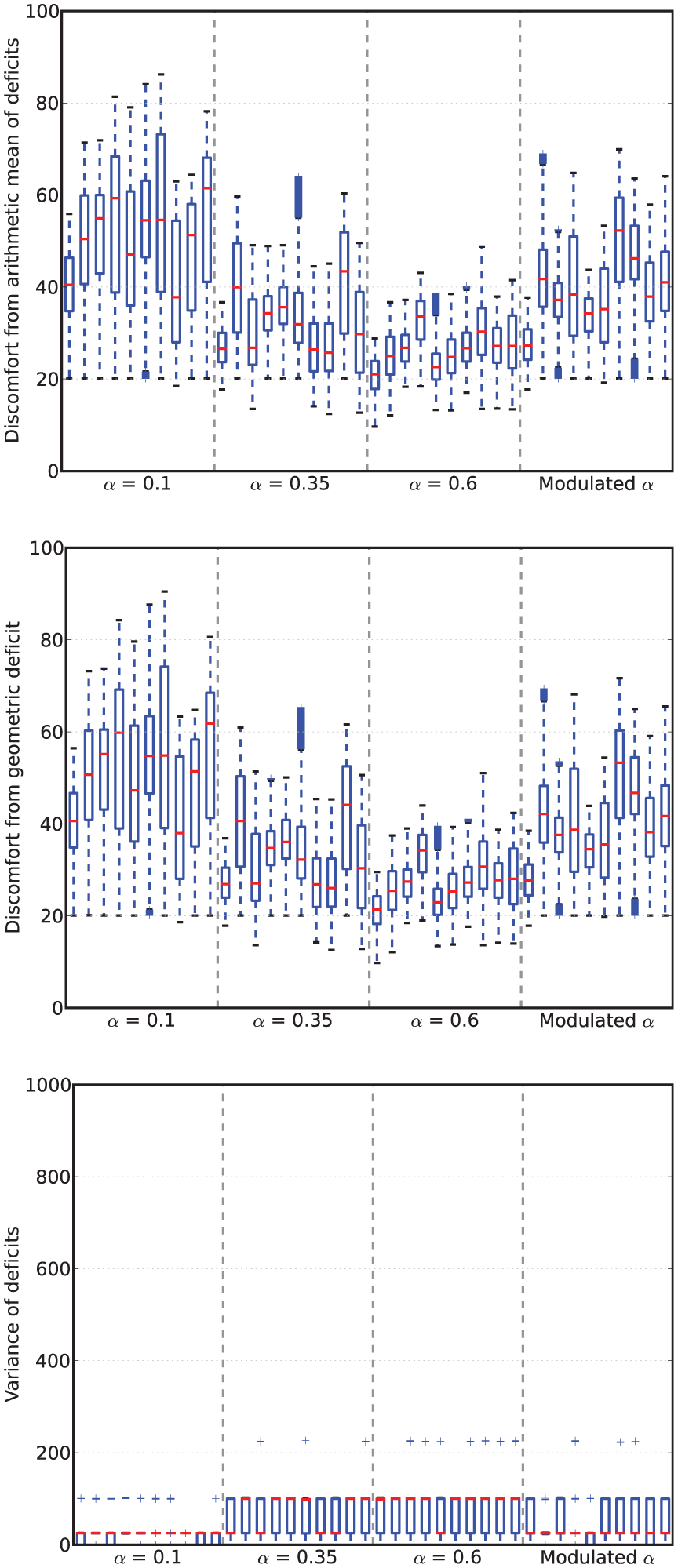
Viability metrics from runs in Experiment 1. In the box-and-whisker plots, the red lines show the medians, the boxes contain the middle quartiles of the data and thus give an indication of the distribution of the deficits over time.
In this environment, the robot survived the full six minutes every run, confirming that it is not a challenging environment. However, there are clear differences between the four conditions.
For fixed values of
Examining the behavior of the robot in terms of persistence and opportunism, we see in Figure 9 that for fixed values of
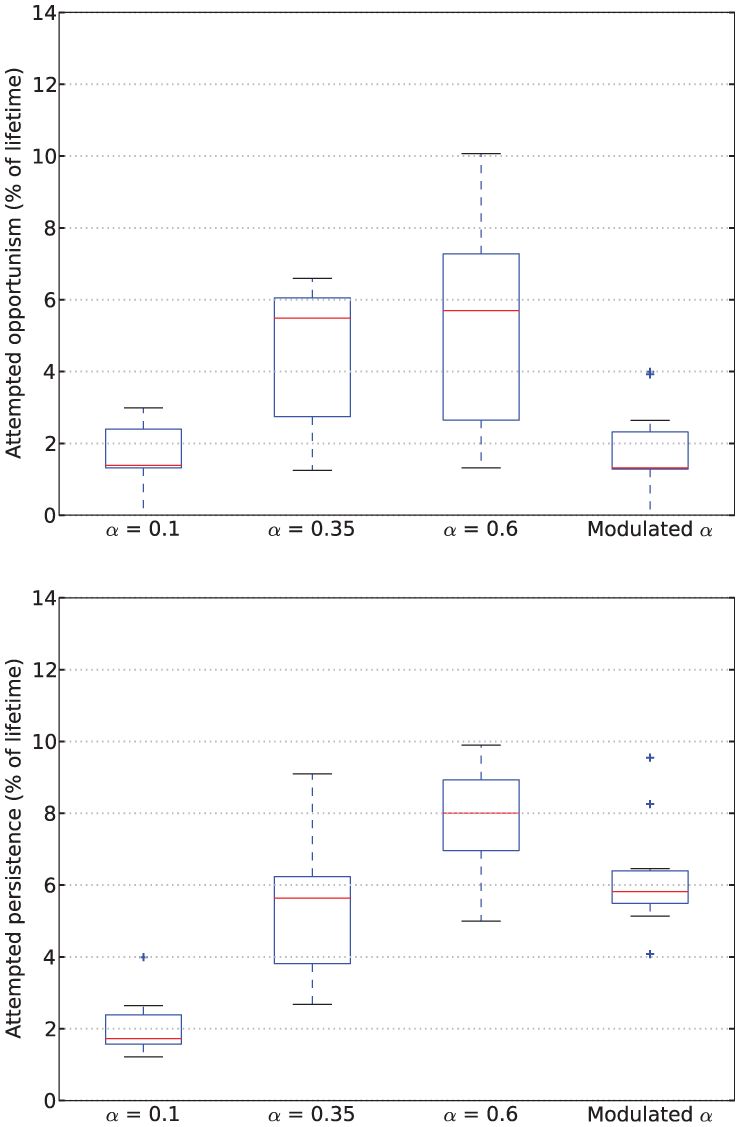
Rates of attempted opportunism (top) and attempted persistence (bottom) from Experiment 1.
Turning to the results for the Modulated
However, looking at persistence and opportunism, we see in Figure 9 a very different picture: it shows that opportunism has rates similar to those of
Comparing the arithmetic and geometric discomforts (Figure 8, top and center), the choice of either metric makes little difference to the overall picture: low and high levels of discomfort remain low and high whichever definition is used. At high levels of discomfort (observed in the LF
5 Experiment 2: Difficult access to resources
In the benign environment of Experiment 1, there was little cost for attending to the current need until satiated, since both types of resources were easily perceived and accessed. In this second experiment, we introduce a cost associated with the access to resources, more precisely with the ability of the robot to perceive the resources.
5.1 Experimental setup
In this environment (Figure 10), the robot might need to wander around the environment before it can detect some of the resources. To achieve this, we have:
placed an obstacle (white cardboard box) in the center to make it more difficult for the robot to detect resources on the other side of the arena;
placed resources so that one area contains only food resources and the other only drink resources.
A diagram and photo of the arena used for Experiment 2.
To consume the resources, the robot needs to move from one section of the arena to the other. In this case, being ‘distracted’ by attempted opportunism would prevent it from doing this efficiently and hence would have a negative impact on viability. On the other hand, the robot still needs to show appropriate levels of persistence in order to consume enough of a resource to counterbalance the growth of the deficit during the exploratory journey from one section of the arena to the other and then back again.
This environment is symmetric in terms of the properties listed in Section 4.1 for the first experiment, although not exactly in the same way. Both environments are identical regarding properties (1)–(4), but the environment in the present experiment differs regarding properties (5) and (6) as follows:
5. Distribution of resources: reflection in one diagonal (along which the robot is facing at the beginning of each run) would result in the resources swapping (complementary symmetry: red ↔ green); reflection in the other diagonal would leave the resources unchanged (mirror symmetry: red → red, green → green).
6. The starting position of the robot (a corner of the arena) affords an equal view of the two areas, and hence resources.
5.2 Experimental conditions tested
We ran the robot in the same Low, Medium and High fixed
5.3 Results
Figure 11 shows deficit–space plots from example runs under each condition. Figure 12 shows the discomfort metrics for each run plotted against time and Figure 13 the distribution of the performance metrics for each run. Note that in cases where the robot died, when calculating mean discomforts over the 6-min run, we set the discomfort of a dead robot to be 100—the maximum value—so that the metrics took account of the death of the robot. Figure 14 shows rates of opportunism and persistence.
Example deficit–space plots from Experiment 2; from top to bottom: sample runs with LF
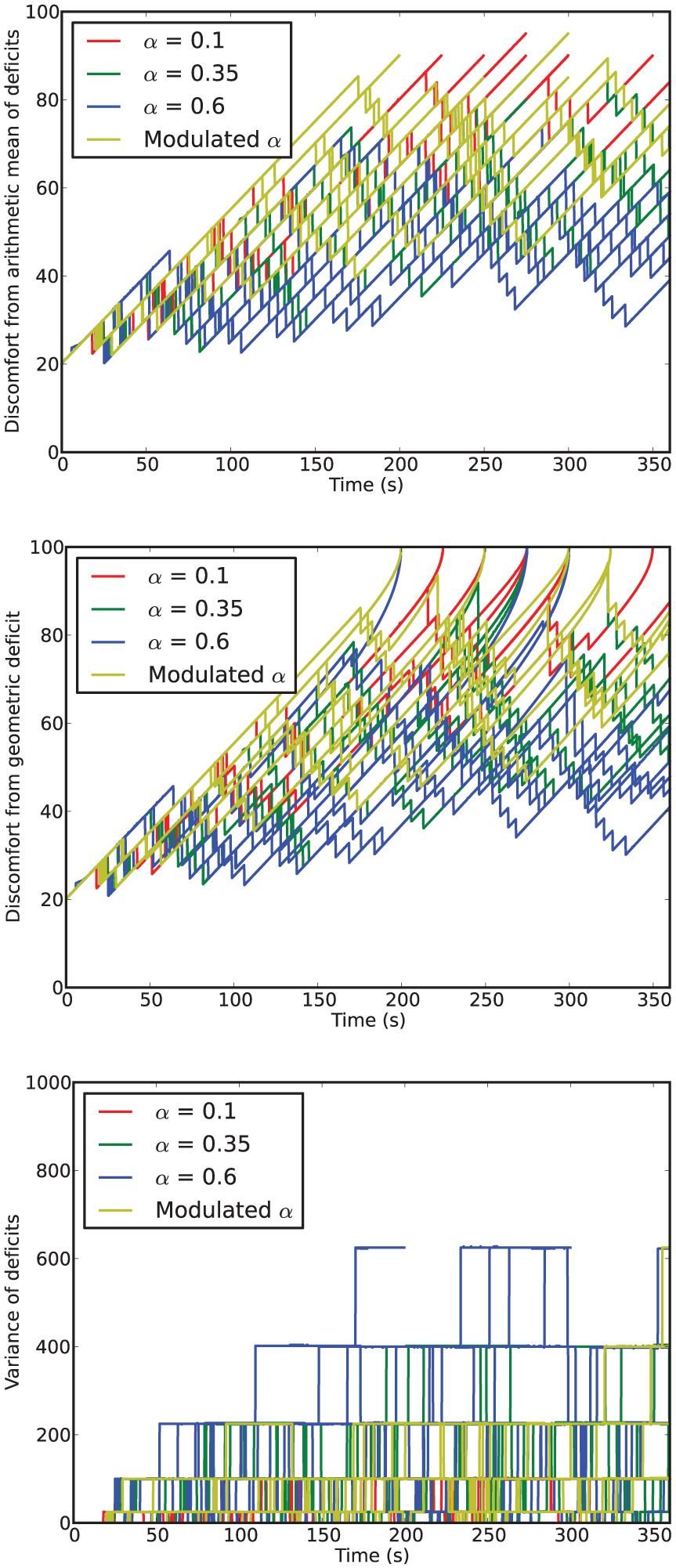
Time series of discomfort metrics from Experiment 2.
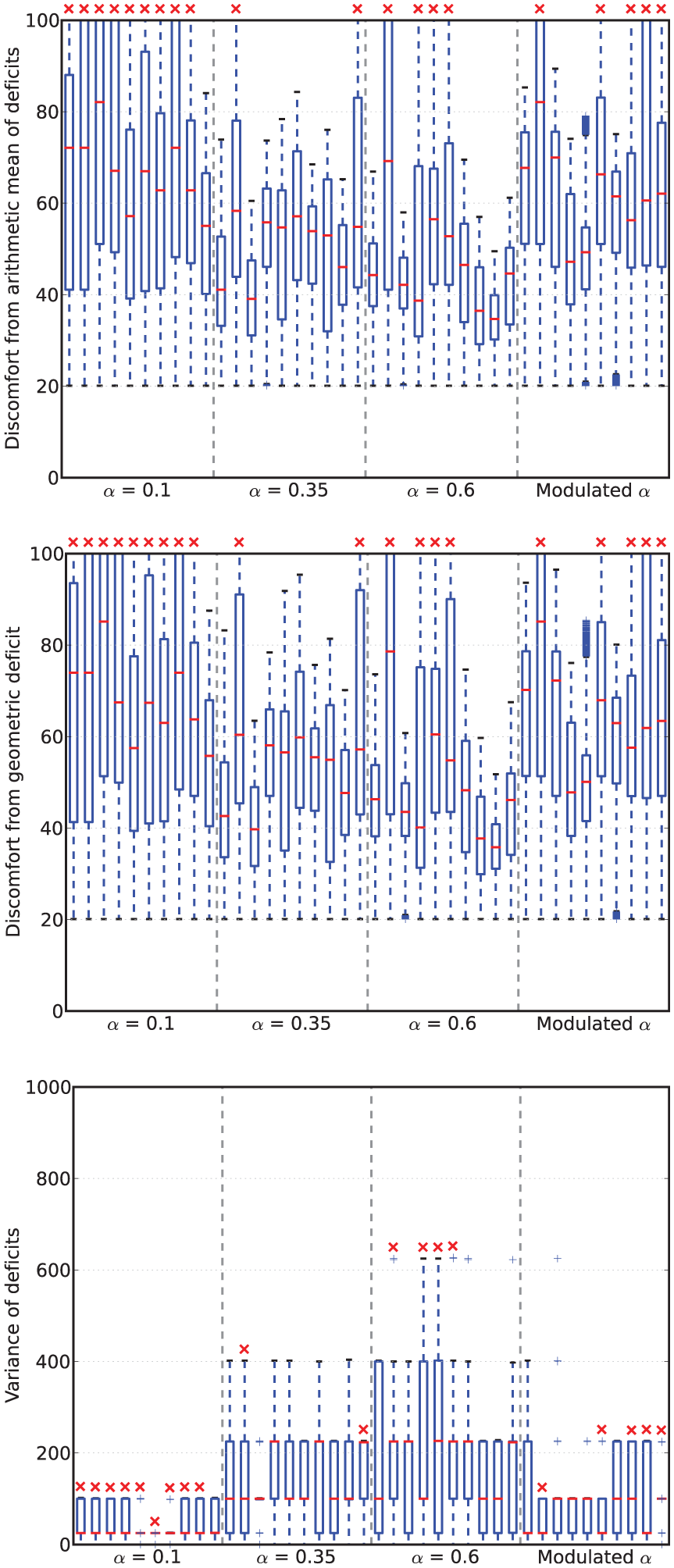
Viability metrics from runs in Experiment 2. In the box-and-whisker plots, the red lines show the medians, the boxes extend to the upper and lower quartiles of the data. The red crosses indicate those runs in which the robot died.
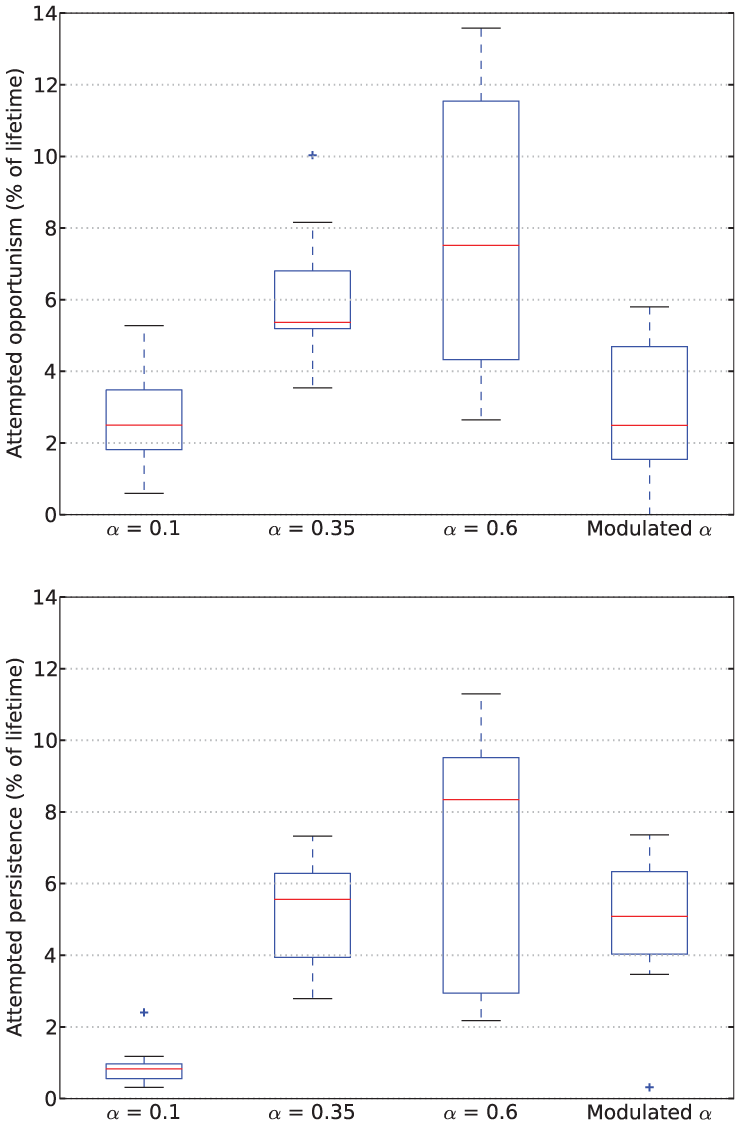
Rates of attempted opportunism (top) and persistence (bottom) from Experiment 2.
In terms of survival, this environment is clearly more challenging than that used in Experiment 1: all but one of the LF
However, the number of deaths does not match the pattern observed in the discomfort. Here, the mean geometric discomforts for increasing
Looking at the variance, we see that it increases with increasing fixed
6 Experiment 3: Introducing asymmetry
In the two previous experiments, we maintained symmetry between the two resources on a number of key features, as mentioned in Sections 4 and 5. In this third experiment, we introduce asymmetry both in the environment and in the role of pleasure.
6.1 Experimental setup
Asymmetry is introduced in the environment (Figure 15) regarding availability of the two types of resource, making one of them abundant—by using six food resources—and the other scarce—only two drink resources are used, and the presence of the box in the middle of the environment ensures that they cannot be perceived from all the locations in the environment.
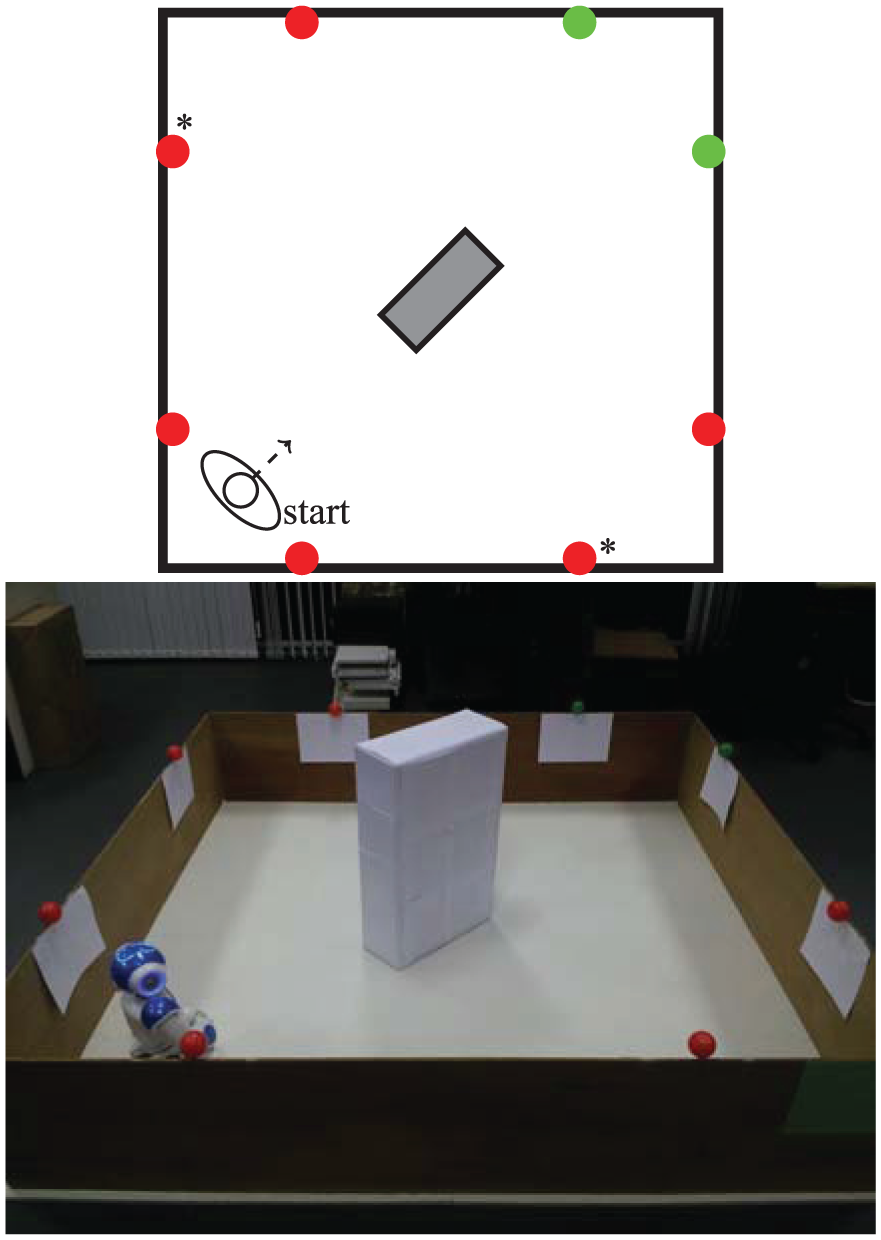
A diagram and photo of the arena for Experiment 3, showing the asymmetric resource layout. To achieve this layout, we modified the environment used in Experiment 1 by replacing two of the food resources with drink resources (marked with asterisks in the diagram). The barrier (the box in the center of the arena) used in Experiment 2 was placed in the center of the arena, so that when seeking a drink resource the robot would sometimes go the long way around. Without the box, the robot would simply go directly to the drink resources, and this would mean that, although there were fewer of them, there would have been little decrease in their availability, making the environment too ‘easy’.
For comparison purposes, in two of the conditions tested (Conditions 1 and 2 in Section 6.2) we use a symmetric environment. This environment was identical to the asymmetric environment, except that the food resources marked with an asterisk in Figure 15 were replaced with drink resources. There were thus four of each resource.
One might intuitively hypothesize that the introduction of asymmetry in the number of resources, whilst maintaining the symmetry in the pleasure obtained from both resources, would lead to the case in which pleasure would be maladaptive since it would make the robot consume even more of the abundant resource and neglect the scarce resource. To investigate whether this would be the case, we also introduced asymmetry in the release of pleasure, making one of the two resources more pleasurable than the other. Asymmetry in the role of pleasure is thus produced by changing the amount and the context in which the pleasure hormone was released for each of the resources.
We made the effects of pleasure from consuming the two resources asymmetric, giving rise to two conditions: more pleasure associated with the consumption of the abundant resource, and more pleasure associated with the consumption of the scarce resource. To better assess the effects of this ‘extra pleasure’, we decided to decouple it from the nutritional value of the resources and hence from the satisfaction of physiological needs. This separation is also supported by recent results in neuroscience (Tellez et al., 2016).
We have thus introduced a second mechanism for the release of the pleasure hormone on top of the first mechanism linked to the changes in the satisfaction of physiological needs: successful execution of the consummatory behavior of one of the resources (consuming either the abundant or the scarce resource, depending on the condition) results in a release of 40 units of the pleasure hormone, independent of any release due to the regulation of the homeostatic variables (Figure 16). This additional trigger for hormone release can be considered analogous to ‘tasting good’, or generally just ‘liking’—sensory pleasure from just the act of consumption, regardless of the physiological benefit or utility of what is consumed (Cabanac, 2010; Frijda, 2010).
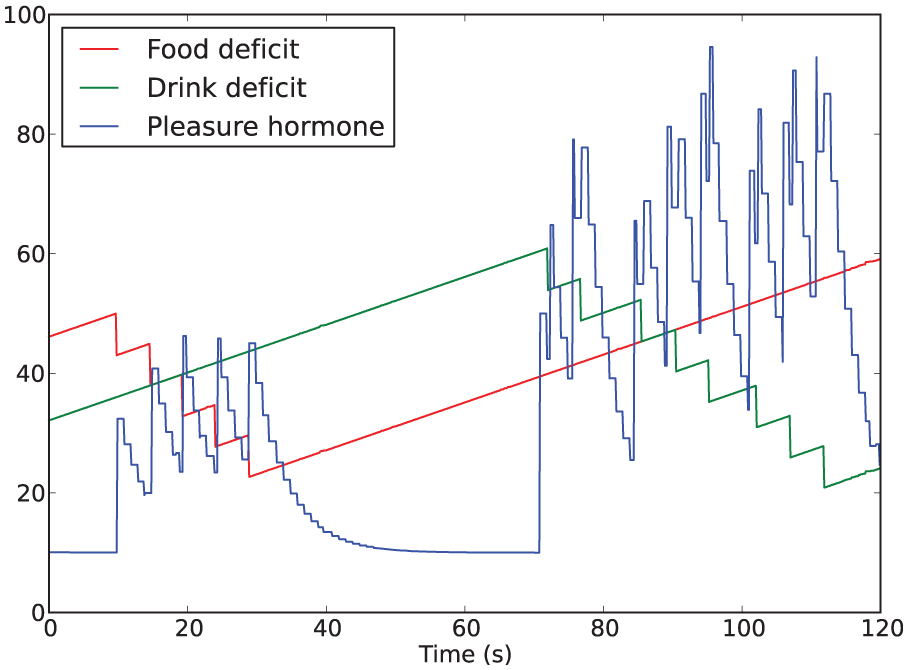
Example of the hormone dynamics from Experiment 3 with hormone also released on execution of the drinking behavior (Condition 4). Consuming drink results in paired spikes showing the two separate releases of hormone, and a larger overall release of hormone.
Considering the environmental properties in Section 4.1, the present environment is identical regarding properties (1) and (3), but differs regarding the remaining properties as follows.
2. The ‘nutritional value’ of both resources is identical in all conditions (though these values are different from those in Experiments 1 and 2).
4. The amount of pleasure hormone released as a result of homeostatic changes due to the consumption of resources is the same for both resources. However, in some conditions, there is an additional release of pleasure hormone associated with the consumption of one type of resource, but not the other.
5. The resources are symmetrically or asymmetrically distributed, depending on the condition. In the symmetric conditions the placement and distribution of resources is the same as in Experiment 1. In the asymmetric conditions, mirror symmetry is used in the axis along which the robot is facing at the beginning of each run. However, since we have different numbers of each resource, distribution is asymmetric on the other axis.
6. The starting position of the robot (a corner of the arena) affords an equal view of the two areas, and hence resources.
6.2 Experimental conditions tested
In this experiment the fixed
Combining the two sets of criteria—asymmetry in the environment and asymmetry in the role of pleasure—we obtain six conditions. These six theoretical conditions in fact amount to five experimental conditions, since in the symmetric environment adding additional asymmetrical hormone for either of the two resource types would give exactly equivalent conditions. Therefore, we report only the five distinct experimental conditions (summarized in Table 1) as follows.
Condition 1, baseline: symmetric environment, symmetric pleasure released only from changes in essential variables.
Condition 2: symmetric environment, asymmetric pleasure. The pleasure hormone is released from essential variables, and there is additional release from consuming drink.
Condition 3: asymmetric environment, symmetric pleasure released only from changes in essential variables.
Condition 4: asymmetric environment, asymmetric pleasure. The pleasure hormone is released from essential variables, and there is additional release from consuming drink—the scarce resource.
Condition 5: asymmetric environment, asymmetric pleasure. The pleasure hormone is released from essential variables, and there is additional release from consuming food—the abundant resource.
The five conditions tested in the Experiment 3.

In order that the robot’s actions have a more fine-grained effect on its internal deficits, which will allow us to see any asymmetries more clearly, we have reduced the ‘nutritional value’ of both resources from 10 to 7 units (established empirically). This means that, in Equation (2),
As in Experiments 1 and 2, we conducted 10 runs in each condition, giving a total of 50 runs.
6.3 Results
Figure 17 shows deficit–space plots from example runs under each condition. Figure 18 shows the discomfort metrics for each run plotted against time and Figure 19 the distribution of the performance metrics for each run. Figure 20 shows rates of opportunism and persistence. Table 2 shows means of the geometric discomfort and of the variance of the deficits over all the runs. See captions for more detailed explanations.
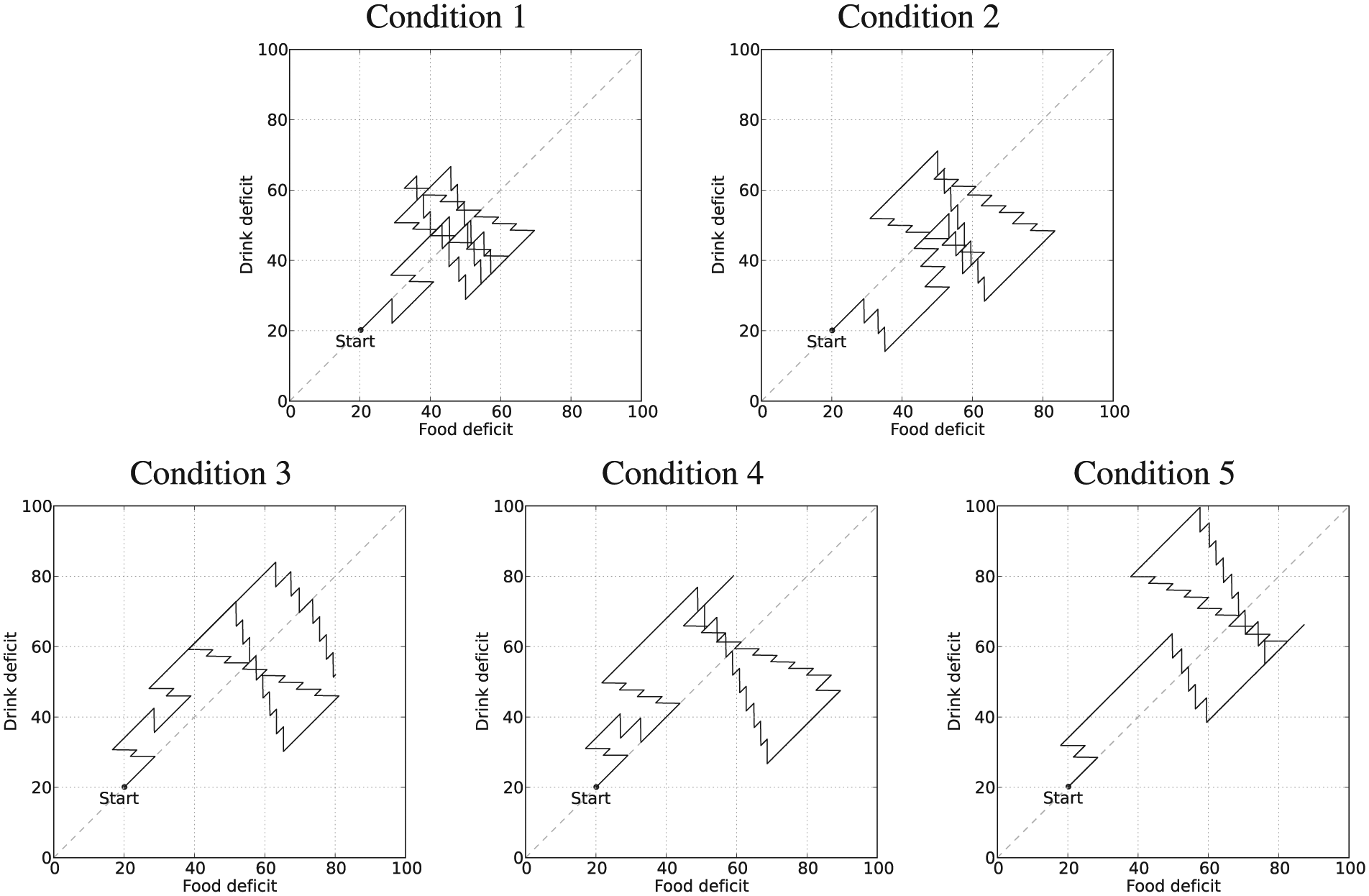
Example deficit–space plots from Experiment 3. Diagonal lines have been added to make the asymmetry more visually clear.
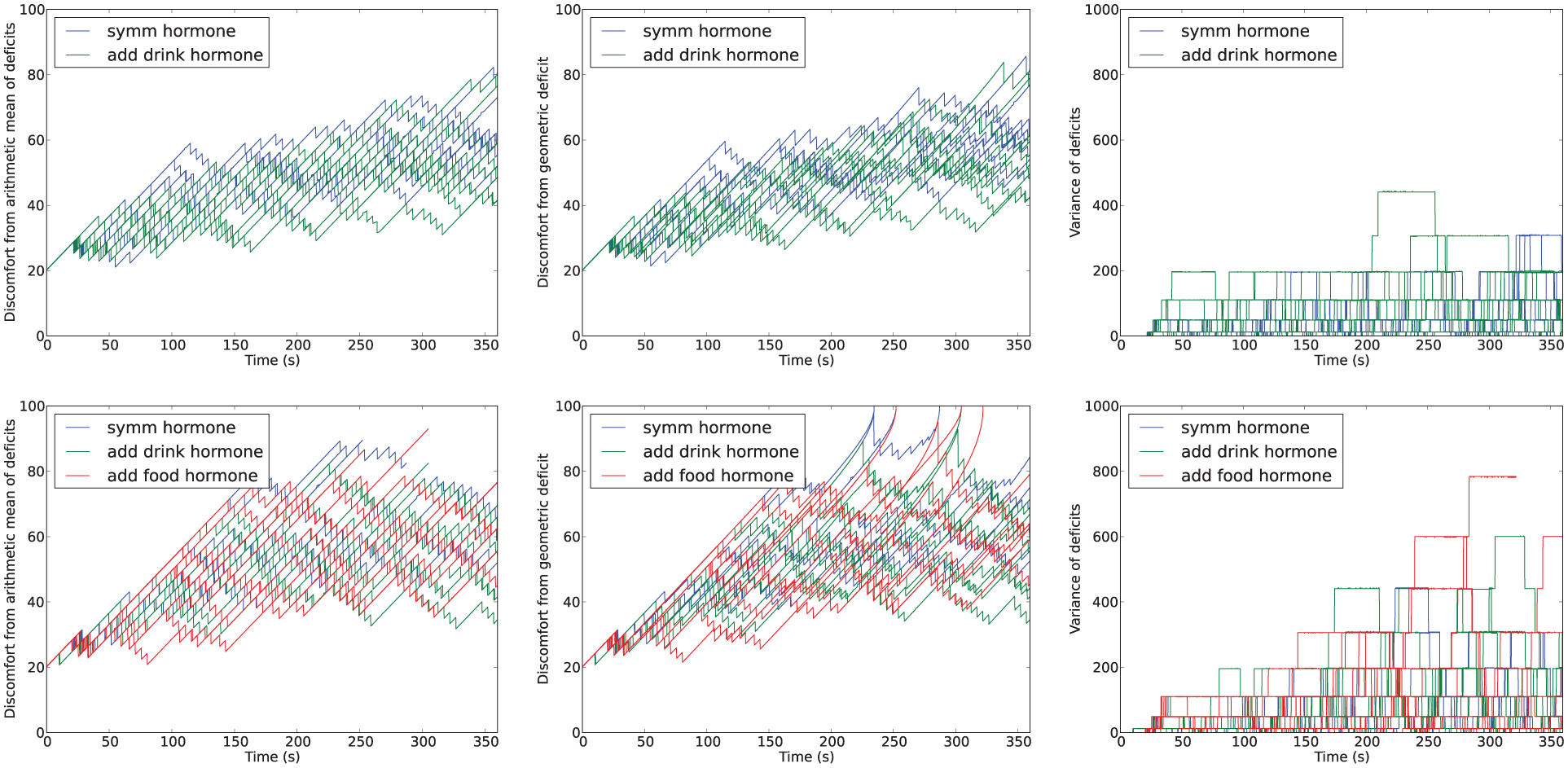
Time-series of metrics from Experiment 3. Top: in the symmetric environment (Conditions 1 and 2, equal availability of each resource); bottom: in the asymmetric environment (Conditions 3, 4 and 5, scarce drink resource).
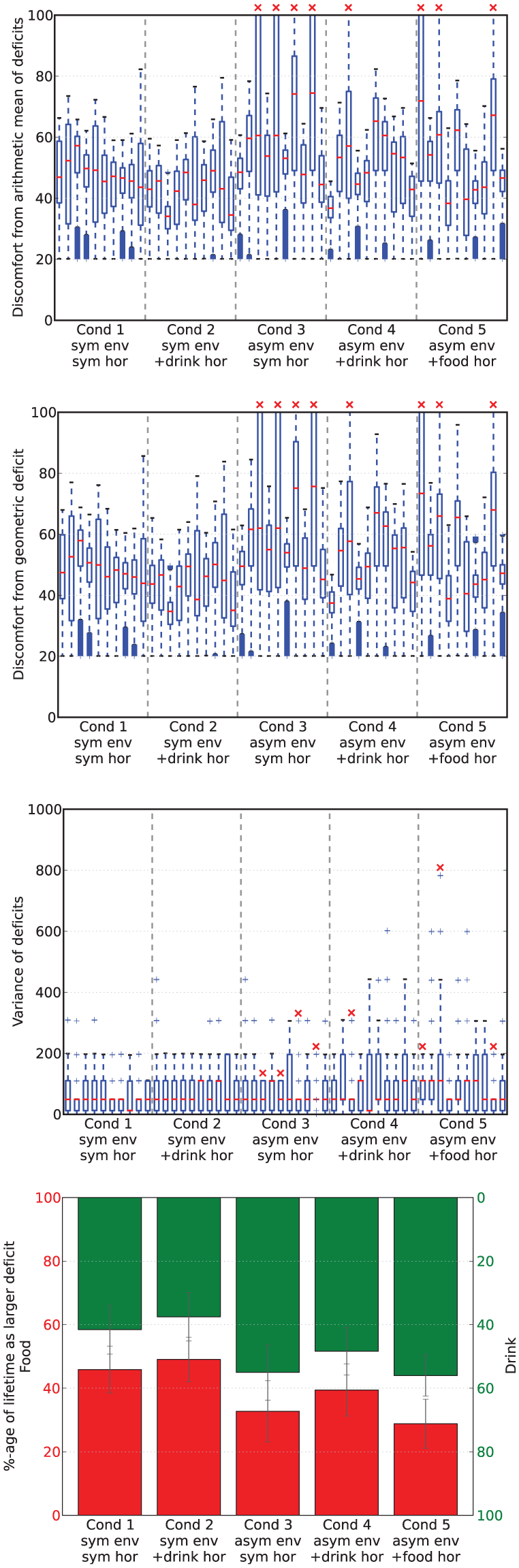
Metrics from runs in Experiment 3. In the box-and-whisker plots, the red lines show the medians while the boxes extend to the upper and lower quartiles of the data. The red crosses indicate those runs in which the robot died. The final plot gives an idea of the balance, in terms of time, between homeostatic deficits in each scenario, showing the percentage of the lifetime in which each of the deficits was larger. The lower bars (in red, read from bottom to top) show the percentage of time that the food deficit was larger, and the upper bars (in green, read from top to bottom) show the percentage of time that the drink deficit was larger.
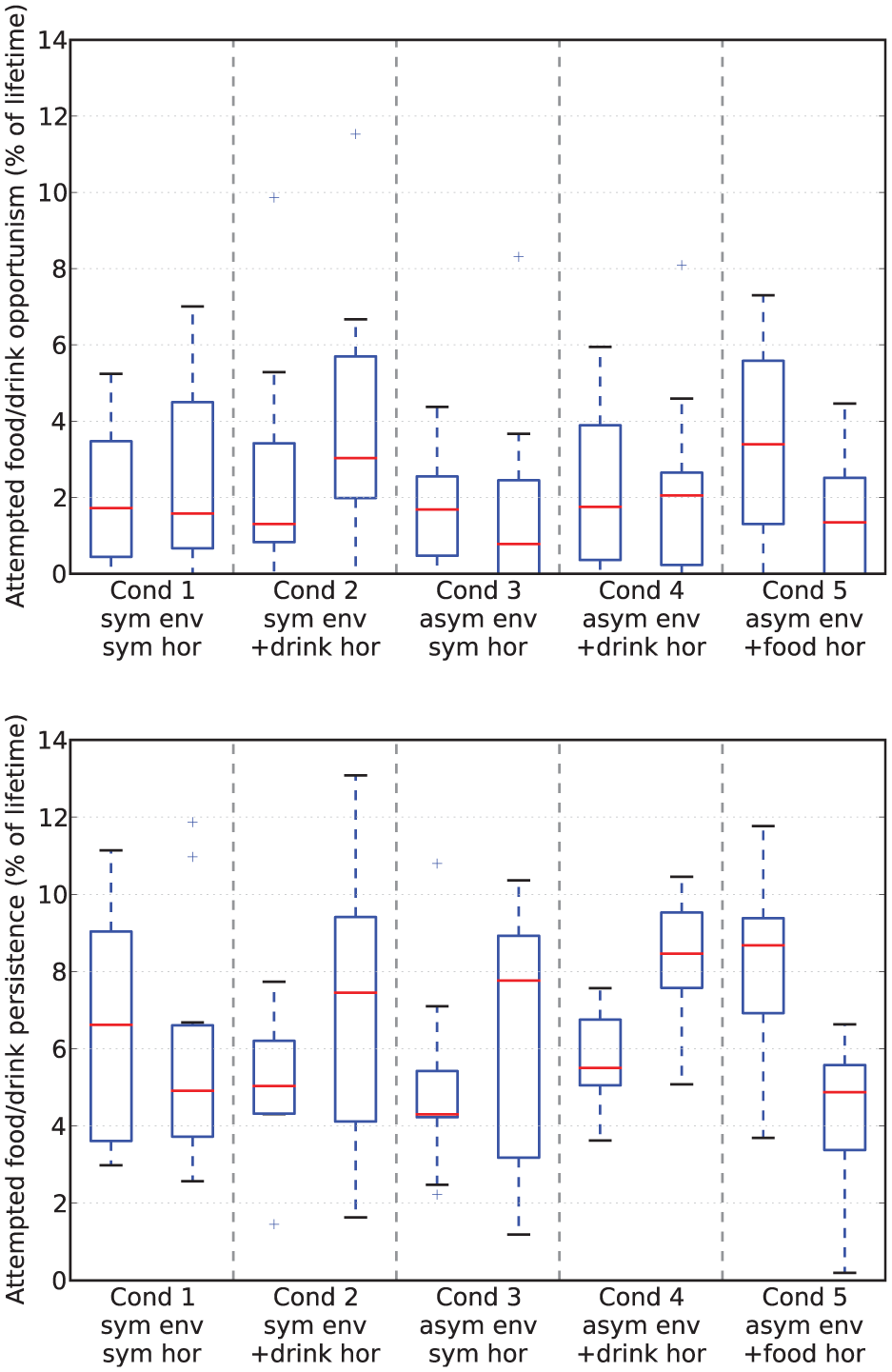
Rates of attempted opportunism (top) and persistence (bottom) from Experiment 3. Due to the asymmetric resource distribution of the environment we have separated opportunism/persistence directed at food (on the left in each pair) and at drink (on the right in each pair). Note that these are not directly comparable to the equivalent values for the previous experiments (the Mo
Means of, respectively, the arithmetic discomfort, the geometric discomfort and the variance of the deficits over all runs in each condition in Experiment 3.

Looking at Figures 19 and 18, we can see that there were no deaths in either of the two conditions for release of the hormone in the symmetric environment. In comparison, the asymmetric environment was clearly more challenging, with four deaths (two from thirst, two from hunger) occurring in the condition in which the hormone release depends only on the homeostatic variables, one death (from hunger) when there was additional hormone release upon drinking (additional hormone release linked to the scarce resource), and three deaths (two from thirst, one from hunger) with additional hormone release upon eating (additional hormone release linked to the abundant resource).
Looking at the rates of attempted persistence and opportunism (Figure 20) we can see that, as in the Mo
Looking at Table 2, we can see that the additional pleasure hormone decreases discomfort (i.e. improves homeostatic management) but increases the variance of the deficits. These changes occur when the extra pleasure comes from eating either of the resources; although we expected this to happen when extra pleasure was associated with eating the scarce resource (drink), we had not anticipated that this would also happen when extra pleasure was associated with eating the plentiful food resource (Experimental Condition 5). Although the increase of variance introduced by the extra pleasure might in principle seem to indicate worsening of homeostatic management, this is not necessarily the case. To have a better understanding of how homeostatic balance is managed, we have also calculated the percentage of time during which each deficit is larger. Results are shown in the bottom plot of Figure 19. For each set of runs, the lower red bar shows the percentage of the lifetime of the robot during which food was the larger deficit, while the upper green bar shows the percentage of time during which the drink deficit was larger. In the symmetric environment with no additional pleasure (Experimental Condition 1, shown by the leftmost pair of bars), the two deficits are well balanced with respect to each other by this metric. The addition of extra pleasure for the drink resource (Experimental Condition 2, shown by the second pair of bars) results in a small shift of this balance so that the drink deficit is more often the smaller of the two deficits. The asymmetric environment, where the drink resource is scarce (Experimental Condition 3, shown by the third pair of bars), clearly shows an asymmetry in the balance of the deficits, with the deficit corresponding to the scarce resource (drink) more often being the larger of the two deficits. Additional pleasure from consuming one resource (Experimental Conditions 4 and 5, shown respectively by the fourth and fifth pair of bars) decreases the amount of time that the deficit corresponding to the more pleasurable resource is larger, as in the symmetric environment. When the extra pleasure is linked to the scarce drink resource (Condition 4, fourth pair) it has the effect of partially reducing the asymmetry caused by the environment; however, when the plentiful resource (food) is more pleasurable, it adds to the asymmetry caused by the environment.
7 Discussion
In the static and unchanging environment with readily available resources used in Experiment 1, if we only consider the viability metrics such as the discomfort (Figure 8), there is little to distinguish the robot with Modulated
However, if we consider its rates of persistence and opportunism (Figure 9), our results show that the robot with modulated
Let us consider how the hormone-related mechanism contributes to this different behavior. No component in our system is explicitly controlling the rates of persistence and opportunism. However, the interaction between the following two elements gives rise to, on average, prolonged interaction with consumed resources.
The hormone acts to modulate the perceptual salience of environmental cues, and therefore their incentive salience related to motivation.
The interaction context in which the hormone is released—successful consumption of a resource to reduce a homeostatic deficit—means that the relevant resource is necessarily present, likely in a situation where it can be readily perceived and consumed.
Our mechanism thus increases attempted persistence but does not increase the likelihood of attempted opportunism. Attempted opportunism more often occurs in contexts where no resource has been consumed recently, and hence the hormone has fallen to its background level; this background level is what has a dominant influence on the level of opportunism. This is seen in the results of Experiments 1 and 2, in which the levels of opportunism for LF
In the more challenging environment with difficult access to resources used in Experiment 2, we observe more interesting differences across the
Note that the Mo
Characterizing the robot’s behavior in this environment in terms of persistence and opportunism in the Mo
As we have already mentioned, with fixed values of
The effects that introducing asymmetry in the environment and in the release of pleasure have on the behavior and viability of the robot (Experiment 3) are clearly visible in our metrics. To describe these effects, let us consider the effects that these types of asymmetry have on an idealized activity cycle, depicted in Figure 21. 8 The activity cycle at the top shows the behavior cycle in a static and symmetric environment, as reflected by the dynamics of the internal deficits. The activity cycle in the center shows the effect of introducing asymmetry in the number of resources of each type: this results in more time spent searching for the scarce resource (corresponding to Deficit B in the figure). Survival is more problematic in this case, as Deficit B comes closer to the fatal limit. The activity cycle at the bottom shows the effect of introducing asymmetric pleasure on top of the asymmetry of the environment. While more time is spent searching for the scarce resource, the whole cycle is shifted away from the fatal limit, showing that this pleasure improves the viability, and hence the survival, of the robot. Note that, to an observer of the behavior of the robot with no access to its internal state, the behavior giving rise to the activity cycles on the center and the right of Figure 21 would appear identical—the robot spends exactly as much time searching and consumes equal amounts of each resource. However, the difference for the robot’s wellbeing is significant as the cycles occur in different places in the deficit space. This difference is also reflected in our persistence metric, the computation of which requires knowledge of the internal deficits.
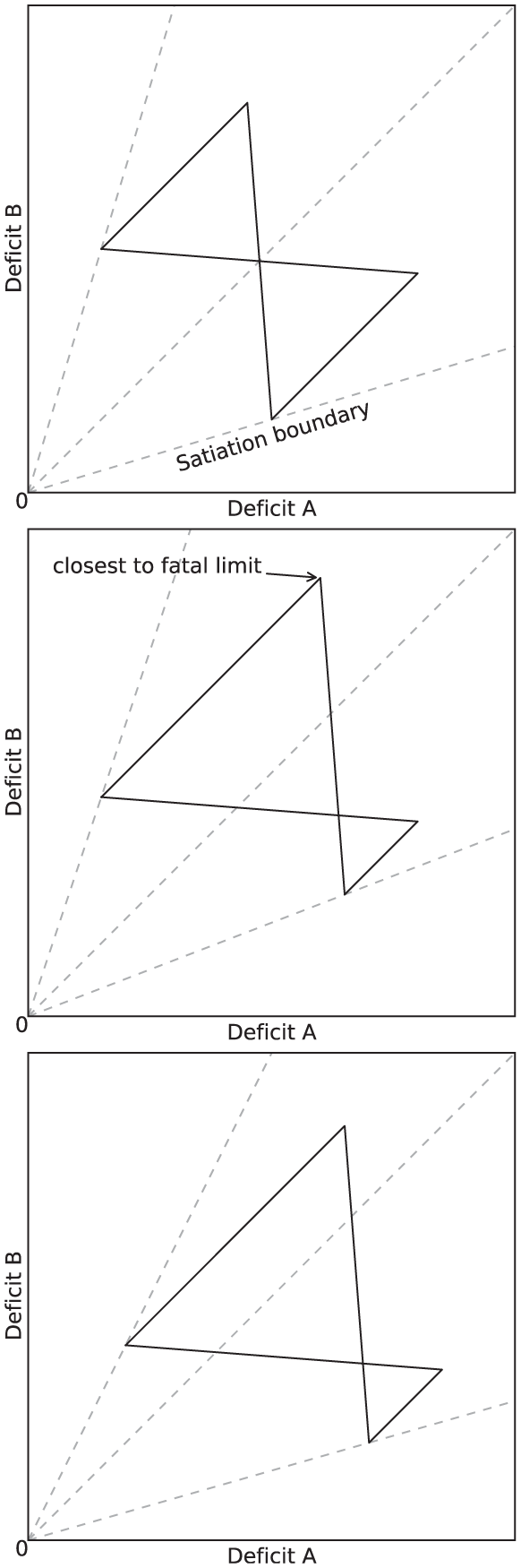
Idealized activity cycles in the deficit space. Top: equal availability of both resources and equal persistence leads to symmetric cycles. Center: Resource B is less available and takes longer to locate, the long search period leading to the agent coming closest to death. Bottom: A larger persistence on Resource B than Resource A shifts the cycle away from the fatal limit; equal amounts of each resource are still consumed, but the agent continues to Resource B further beyond the point where
As we noted in Section 6.3, Table 2 shows that, in each environment, the additional source of pleasure results in: (a) a decrease in (arithmetic and geometric) discomfort (i.e. an increase in comfort) and (b) increased variance between the deficits. These two effects occur even if the extra pleasure happens when consuming the plentiful resource, which intuitively is the ‘wrong’ resource in terms of improving adaptation to the environment. Both effects can be explained by the fact that the additional pleasure increases consumption of the more pleasurable resource, which leads to a large difference between the two deficits; hence, when the less pleasurable resource is eventually consumed, more of it is consumed, as its associated deficit remains the pressing deficit for longer. Due to this, the robot is more frequently in a state where the difference between its deficits is large (increasing variance) and consumes more of both resources (decreasing discomfort by reducing both deficits).
Although increased variance might seem to indicate a worse ‘homeostatic balance’, this is not necessarily the case, since variance is not the only way to conceptualize balance in the system. In order to gain a deeper understanding of the behavior of our robot, we calculated how long each deficit was the larger as a proportion of the lifetime of the robot (Figure 19). We can see that the symmetric environment and symmetric pleasure (Condition 1) leads to roughly equal temporal balance between the two deficits, as would be expected. Adding extra pleasure linked to consumption of one of the symmetric resources (drink, Condition 2) slightly reduces the time for which the deficit associated with that resource is larger. In the asymmetric environment, the temporal balance is shifted by a large amount, and therefore the deficit associated with the scarce resource (drink) is the larger deficit for most of the time. The addition of a second source of pleasure from consuming the scarce resource helps to mostly restore this balance, while added pleasure from consuming the abundant resource slightly increases the imbalance. These shifts in the balance between the two deficits can also be seen in the shifts in the example deficit space plots (Figure 17). Thus the extra ‘asymmetric’ pleasure provides another useful mechanism for adaptation to the environment. In the discussion of Experiment 2, we saw how the pleasure system related to the satisfaction of needs provides a mechanism to control persistence and opportunism that can be used by either the designer or the robot itself to adapt to different environmental circumstances. In the same vein, the additional pleasure introduced in Experiment 3 provides a mechanism to counteract an asymmetry in the temporal balance between the homeostatic variables caused by different environments or changing circumstances.
Let us finally discuss our choice of mechanism to introduce asymmetry in the release of the pleasure hormone. In this experiment, additional sensory pleasure, unrelated to the nutritional value of the resources, was released on successful execution of the consummatory behavior. However, it is worth noting that, in our environment, a similar amount of additional pleasure could have been achieved by increasing s (the scaling factor connecting the second derivative of the deficits with the release of hormone in equation (2)) for just one of the deficits. Since our resources all have the same nutritional value, it would be possible to choose a value for s such that the amount of hormone released was the same as the additional pleasure. It is then worth asking what the difference between these two mechanisms—adding a second source of pleasure or changing the value of the s parameter—is.
In the first place, in an environment where resources could reduce deficits by different amounts, the two systems would differ in that a simple change of s would also scale the difference in the nutrition; therefore, if one resource gave half the nutrition of another, then it would result in half the release of hormone per bite, reducing the persistence in consuming this resource. This is potentially maladaptive in some environments, as we have seen in Experiments 1 and 2. With the additional pleasure mechanism, only one of the sources of pleasure—the pleasure due to the nutritional value—would be halved. The other source of pleasure would change in a way determined by the mechanism used to trigger the release of the pleasure hormone; for example, if the trigger were simply the act of eating, then there would be no change. This separation of the two sources of pleasure presents advantages, such as the ability to discriminate between highly nutritious and less nutritious resources, or the better maintenance of persistence for less nutritious resources, which might be vital in some environments. However, this separation also comes at a cost, as it means that the robot could find a resource that is very pleasurable to eat but has little benefit in terms of the homeostatic variables.
Second, the two mechanisms show a difference in the complexity of the phenomenon that they can model. Changing the scaling factor s corresponds to a very simple pleasure mechanism only linked to nutrition. Compared to this straightforward model, the additional pleasure mechanism offers the potential for more complex roles for pleasure that appear in evolutionarily more complex organisms. It could, for example, allow for a system in which a resource that has not been consumed for some time would give more pleasure than if it had been consumed recently, or for integrating cultural differences in taste-related pleasure.
Third, there is a difference in the temporal dynamics of the two pleasure mechanisms. In our model, consumption of a resource quickly results in a drop in the corresponding homeostatic deficit, and hence nutritional pleasure is an immediate ‘reward’ that can be clearly associated with the ‘eating’ behavior that led to the correction of the deficit. However, in organisms with more complex digestive systems that take time to process food, there is a delay between the eating behavior and the drop in deficit, and hence the nutritional pleasure is delayed (we could talk of a ‘delayed reward’ in terms of reinforcement learning) since it only happens upon digestion of the resource. Changing the scaling factor cannot model these more complex systems, whereas having two sources of pleasure hormone release related to consumption opens the door to keeping an immediate pleasurable reward related to consumption alongside a more delayed pleasurable signal associated with nutrition.
8 Conclusion and future work
We have presented a basic model of pleasure and investigated its effect on the decision making of a motivated autonomous robot. Unlike other work that had looked at pleasure in the context of relatively high-level functions such as learning and memory, we think that pleasure can already play an important role at a more basic level. Therefore, we have focused on lower cognitive functions and investigated the interaction between pleasure, perception and motivation, in particular the incentive salience of survival-related external stimuli. By affecting incentive salience, our pleasure fosters continuing the ongoing interaction, which is one of the main functions attributed to pleasure in the literature. However, we have not taken for granted the other main function often attributed to pleasure: signaling the usefulness of stimuli. We have thus considered two types of pleasure, a well-being related pleasure directly linked to the satisfaction of survival-related needs, and a purely ‘sensory’ pleasure (hedonic quality) unrelated to the satisfaction of needs.
We have framed our study in the context of a classical two-resource AS problem to investigate the effect of these types of pleasure on the viability and decision-making behavior of the robot. We have conducted three sets of experiments varying the following aspects to create increasingly complex AS problems: the availability of (easy or difficult access to) resources and their (symmetric or asymmetric) distribution in the environment, and how the release of pleasure relates to their consumption—either to the ‘nutritional value’ or simply to the act of consuming.
Our results indicate that pleasure, including pleasure unrelated to need satisfaction, has value for homeostatic management in terms of improved viability, as well as in terms of more flexibility in adaptive behavior. Regarding the latter, this is the case specifically in situations where opportunism has a penalty, but increased persistence is beneficial, and where an asymmetry in the availability of resources results in the need to consume each of the resources in different ways in order to achieve good management of homeostasis. Regarding viability, the extent to which the different ‘types’ of pleasure are adaptive or maladaptive depends on the features of the environment and the demands it poses on the task, in addition to the ‘metabolism’ of the robot. In Experiment 1, simply maximizing pleasure (regardless of whether it is related to need satisfaction or not) improved viability. In Experiment 2, constant moderate pleasure (unrelated to need satisfaction) gave the best viability; the pleasure released as a function of need satisfaction was comparable to this by some of the metrics, but additionally more flexible in terms of the behavior of the robot, notably the possibilities it offers to manage persistence and opportunism independently, and hence to display them in the appropriate context. In Experiment 3, in environments with asymmetric availability of resources, the addition of ‘purely sensory’ (not related to need satisfaction) pleasure associated with the scarce resource improved viability.
The work presented here is a first step towards an incremental study of the role of pleasure in AS. The next steps will include the inclusion of resources with different nutritional values, the introduction of dynamic elements in the environment, as well as integrating this model in our social robot Robin.
Footnotes
Authors’ contributions
Both authors contributed equally to this work. LC put forward the initial research hypotheses that were further elaborated jointly. LC and ML designed the experiments jointly. ML implemented the robot architecture, including the details of the pleasure system, conducted the experiments, and carried out quantitative analysis of results; both authors carried out qualitative analysis and contributed equally to the writing of the paper.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by European Commission grant FP7-ICT-248116 (ALIZ-E). The opinions expressed belong solely to the authors.
Notes
About the Authors
![]() .
.