
Abstract
The Charcot–Leyden crystal (CLC) is a major human eosinophil protein that readily crystallizes; these crystals are common in eosinophilic diseases. Although anecdotal existence of these crystals is known in veterinary pathology, definitive reports do not exist, to our knowledge. We identified eosinophilic crystals in a laryngeal myxosarcoma from a 2-y-old, spayed female, Labrador Retriever dog that were tentatively interpreted as CLCs. However, Ziehl–Neelsen acid-fast stain was negative, arguing against CLCs. The crystals stained red with Masson trichrome, precluding collagen. Periodic acid–Schiff and alcian blue were negative. The crystals stained positively with Okajima, and no myoglobin immunoreactivity was detected, supporting their identity as hemoglobin crystals. In the absence of a hematologic abnormality, these crystals were interpreted to be abnormal hemoglobin breakdown products. Protein sequence comparison was pursued to determine whether a protein similar to CLC exists in mammals. Only 3 nonhuman primate species, the Sumatran orangutan (Pongo abelii), rhesus macaque (Macaca mulatta), and cynomolgus monkey (Macaca fascicularis), had a sequence similarity of >80%. Of the crystal-forming residues, 12 of 54 (22%) were different in the Sumatran orangutan and 15 of 54 (28%) were different in the Macaca spp., which may affect the crystallization process. The lack of reports of CLCs in nonhuman species and our results collectively suggest that CLCs are human-specific.
Charcot Leyden crystals (CLCs) are eosinophilic, refractile, non-birefringent, hexagonal bipyramidal structures commonly encountered in human pathology. CLC protein is a major constituent of the primary granules of eosinophils in humans, comprising ~7–10% of total cellular protein. 30 CLC protein expression has been identified in human CD4+ CD25+ Foxp3+ regulatory T (Treg) cells. 7 Initially, CLCs were thought to be lysolecithin acylhydrolase, which has lysophospholipase activity and interacts with the enzyme lysophospholipase. 31 However, lysolecithin acylhydrolase has been renamed galectin-10 17 based on the sequence similarity to galectins and weak lactose-binding activity. 8
In humans, CLCs are found in cytology or histology specimens of eosinophilic inflammation caused by allergic diseases such as asthma, parasitic infections, or any idiopathic eosinophil-rich inflammation in any organ system.3,6 Traditionally, CLCs have been deemed to be a protein found only in humans and nonhuman primates. 9 In domestic species, eosinophilic crystals present in cytology specimens with eosinophils have been interpreted as CLCs. 10 CLCs were also previously believed to be the cause of eosinophilic crystalline pneumonia in mice; however, it is now known that these crystals are composed of a chitinase-like protein, Ym1. 12 Given the rarity of these crystals in research settings, recent publications have deemed these crystals nonexistent, at least in mice.16,23 No work has been reported to prove or disprove the presence of CLCs in veterinary species, to our knowledge. We report herein in silico evidence that suggests that CLCs are unique to humans plus a short review on eosinophilic crystals that was triggered by the identification of eosinophilic crystals in a laryngeal myxosarcoma of a young dog.
A 2-y-old, spayed female, Labrador Retriever dog was admitted to the Cornell University Hospital for Animals (Ithaca, NY) for surgical removal of a laryngeal mass. The patient’s first clinical sign was a diminished bark that appeared 10 mo prior. Over the subsequent months, the bark was lost, and exercise intolerance developed during walks. One month prior to the resection, a whistling or wheezing sound was noted when exercising, which led to the identification of a laryngeal mass on radiographs and via laryngeal exam. On examination at Cornell, this finding was corroborated on computed tomography (CT) scan, which revealed a 1 × 1.3 × 1.9 cm laryngeal mass caudal to the epiglottis with no evidence of lymph node metastasis.
At surgery, the mass was globoid, multinodular, smooth, firm, tan-to-gray, and occupied ~85% of the airway. A single stay suture was placed through the laryngeal mass, and the mass was bluntly dissected until ~90% of the mass had been removed. The resected mass was sent to the Cornell anatomic pathology service. Five soft tissue fragments ranging from 0.5 × 0.5 × 0.3 to 1.5 × 1.5 × 1 cm were received, fixed in 10% buffered formalin for 24 h, processed routinely, embedded in paraffin, and cut at 4-µm thickness. Histologic sections were stained with hematoxylin and eosin (H&E), Ziehl–Neelsen (ZN) acid-fast stain, periodic acid–Schiff (PAS), alcian blue (pH 2.5), Masson trichrome, and Okajima for hemoglobin and myoglobin. Immunohistochemistry for myoglobin was performed (Leica BOND MAX IHC & ISH Staining System, Leica Microsystems, Buffalo Grove, IL). Formalin-fixed, paraffin-embedded sections were deparaffinized, rehydrated, and subsequently blocked with 3% hydrogen peroxide (Leica). Antigen retrieval was performed (Bond Epitope Retrieval Solution 2, Leica) for 30 min. Sections were incubated with anti-myoglobin (rabbit polyclonal; Cell Marque, Rocklin, CA) for 60 min. Slides were then incubated with a polymer (Bond Polymer Refine Detection Kit, Leica) for 30 min. The slides were developed with DAB (3,3’-diaminobenzidine) chromogen and counterstained with hematoxylin. The external and internal positive controls and negative controls were adequate.
Histologic examination revealed a multinodular, poorly demarcated neoplasm expanding the submucosa of the larynx and effacing the normal architecture. The neoplasm was composed of loose, haphazardly arrayed neoplastic spindle cells forming streams and bundles on a background of pale myxomatous matrix (Fig. 1A). Neoplastic cells had indistinct cell borders, a small-to-moderate amount of eosinophilic, elongated, fibrillar cytoplasm, and a round-to-elongated nucleus with vesicular chromatin that lacked a prominent nucleolus. Anisocytosis and anisokaryosis were moderate, and 3 mitotic figures were present in ten 400× fields. Multifocally throughout the neoplasm, accounting for ~30% of the sections examined, were areas of hemorrhage and necrosis with small numbers of neutrophils and lymphocytes. Occasionally embedded in the neoplasm were small deposits of radiating eosinophilic crystalline spicules not associated with hemorrhage or inflammation (Fig. 1A, inset).
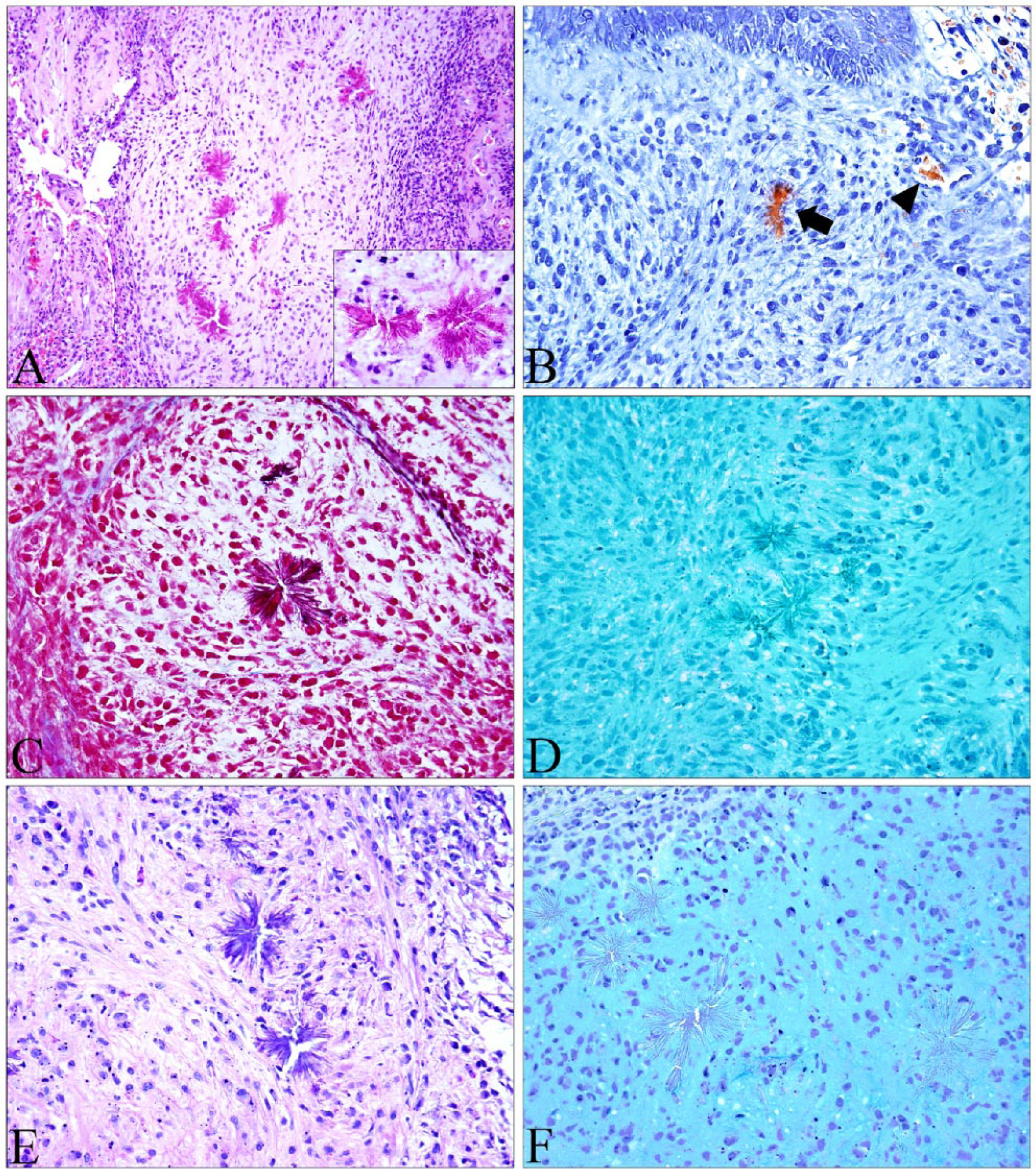
Histologic features and staining properties of eosinophilic crystals within a canine myxosarcoma.
The neoplasm was diagnosed as a myxosarcoma, and the eosinophilic crystals were interpreted as Charcot–Leyden crystals. However, unexpectedly, Okajima highlighted these crystals orange as well as the erythrocytes in blood vessels, a finding more compatible with hemoglobin or myoglobin crystals (Fig. 1B). Immunohistochemical staining for myoglobin was performed and did not immunoreact with the crystals, ruling out myoglobin (data not shown). ZN acid-fast stain has been shown to highlight CLCs in tissue sections, 28 and Masson trichrome stains CLCs red-to-purple. 3 Although the trichrome did stain these crystals deep-red (Fig. 1C), precluding collagen, ZN was negative, suggesting that these are unlikely to be CLCs (Fig. 1D). Alcian blue and PAS confirmed the mass to be a myxosarcoma based on the PAS- (Fig. 1E) and alcian blue– (Fig. 1F) positive myxomatous matrix surrounding the neoplastic cells.
Eosinophilic crystals are rarely encountered in veterinary histopathology with the exception of the mouse and rat. In mice, buildup of Ym1, a chitinase-like protein, in the lung causes eosinophilic crystalline pneumonia and can have significant consequences. 12 In the rat, hemoglobin crystals are fairly common and are thought to be an intermediate breakdown product of hemoglobin.4,15 In our case, the eosinophilic, refractile appearance of the crystals and unusual location sparked investigation into these and other extracellular eosinophilic deposits seen in histopathology. Our investigation included reports of sclerotic collagen, amianthoid fibers, skeinoid fibers, and Charcot–Leyden crystals from various species (Table 1). Rosenthal fibers, seen in gliosis and in brain neoplasms such as pilocytic astrocytomas, and Reinke crystals (seen in Leydig cell tumors of the testis) were excluded based on the anatomic location. Amianthoid fibers are stellate-shaped deposits of crystalline eosinophilic extracellular matrix composed of collagen, most commonly encountered in the palisaded myofibroblastoma in humans.13,26 Skeinoid fibers are eosinophilic stromal fibrillar aggregates seen in neurogenic spindle cell tumor and gastrointestinal stromal tumor in humans. Ultrastructurally, skeinoid fibers are short-spaced collagen and appear as skeins of yarn. 20 Amianthoid fibers, skeinoid fibers, and sclerotic collagen are all composed of collagen and stain blue with Masson trichrome. In our case, the crystals were dark-red to purple when stained with Masson trichrome, which suggested that the crystals were not composed of collagen. In general, keratin, muscle fibers, cytoplasm, and fibrin are red, and collagen, bone, and amyloid are blue with this stain. PAS was applied for potential monoclonal immunoglobulin deposition; however, the crystals in our case were PAS-negative or a light-blue to purple and did not correspond to the underlying matrix present throughout the neoplasm. Alcian blue also confirmed the discordance between the matrix and the crystals. The only positive stain that highlighted these crystals was Okajima, a stain for hemoglobin and myoglobin. Negative immunoreactivity against myoglobin antibody ruled out myoglobin.
Eosinophilic crystalline structures and their staining properties.

Apart from the rat, it is believed that hemoglobin from all other species crystallizes in vivo only if structurally abnormal.5,15 In our case, no hematologic abnormality was observed; however, a blood smear was not performed. Given that these crystals were present within a tumor, the microenvironment of the neoplasm may have predisposed to hemoglobin crystal formation. Interestingly, these crystals were not associated with areas of hemorrhage, although hemorrhage was noted elsewhere in the neoplasm. The significance of these crystals in this patient is unclear and was interpreted to be tumor-related.
Although the neoplasm lacked eosinophils, the crystals were initially interpreted as CLCs, and we speculated that T-regulatory cells present in the neoplastic population could have been the source. However, the histochemical staining did not support the identification of CLCs. Furthermore, the lack of reports of CLCs in the veterinary literature caused us to believe that CLCs may be human-specific. In one study, eosinophils isolated from humans and guinea pigs were monitored to assess crystal formation after cell lysis using light microscopy and electron microscopy. 9 Crystal formation was present in human eosinophils but not in guinea pig eosinophils, and this reaction to injury of the eosinophils of humans and those of the guinea pig was concluded to be characteristic for each of these 2 groups: primates, including humans, and nonprimate mammalian species, including guinea pigs. 9
To further investigate the possibility of CLC formation in nonhuman mammalian species, a protein BLAST search was performed on mammalian protein sequences using default settings of UniProt (http://www.uniprot.org). The query sequence was human galectin-10 (UniProt entry Q05315 v.3), and the target database was mammals. The Sumatran orangutan (Pongo abelii) had a protein with the highest sequence similarity at 87% with conservation of the domain. The rhesus macaque (Macaca mulatta) and cynomolgus monkey (syn. crab-eating macaque, long-tailed macaque; Macaca fascicularis), common primates in research, had ~80% sequence similarity with conservation of the domain. Of the 54 amino acid residues involved in the crystal formation of CLCs, 18 42 amino acids were conserved in the Sumatran orangutan and 39 in the rhesus macaque and cynomolgus monkey (Fig. 2). The differences in the amino acids involved in crystal packing may explain the lack of crystal formation in nonhuman mammalian species. Genome sequencing has demonstrated that the CLC protein is absent in the mouse genome, supporting the lack of reports of this structure in mouse models of human disease in the literature. 16 Similarly, the lack of CLC protein expression in mouse eosinophils has been described. 23
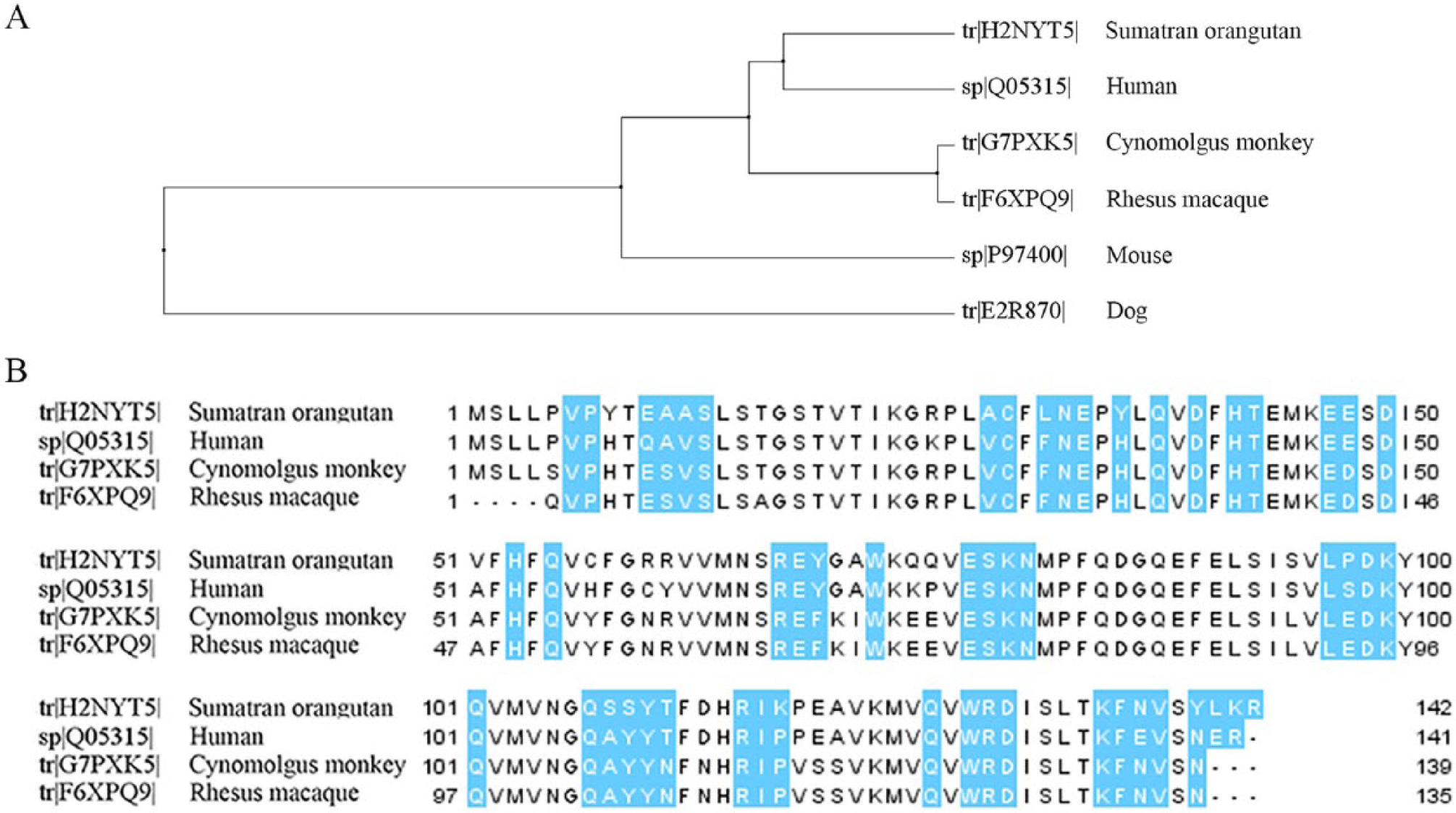
Protein BLAST results using human Charcot–Leyden crystal (CLC) protein.
The presence of eosinophilic crystals in a myxosarcoma in a dog led to research into CLCs in nonhuman mammalian species. The crystals in the myxosarcoma were composed of hemoglobin, and likely represent an intermediate product of erythrocyte breakdown in the tumor microenvironment. Although first considered CLCs, the lack of ZN staining, positive Okajima staining, and negative immunoreactivity against myoglobin antibody collectively support their identity as hemoglobin crystals. Simple in silico tools were able to demonstrate that CLCs may be human-specific. Given these results, caution should be used in interpreting eosinophilic crystals in samples from veterinary species, and histochemical stains should be used to support the identification of these structures.
Footnotes
Acknowledgements
We thank the histopathology technicians at Cornell University College of Veterinary Medicine for assistance with case processing.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.