
Abstract
Species identification of 200 beta-hemolysin–producing canine staphylococcal isolates was performed using a recently described polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) method (based on MboI restriction of a pta gene fragment), supplemented with biochemical testing and sequencing of housekeeping genes. The PCR-RFLP method misclassified a small fraction (approximately 1%) of the Staphylococcus pseudintermedius population as a result of heterogeneity in the MboI restriction site. A potentially novel species within the Staphylococcus intermedius group (SIG) was found, having closest similarity to S. intermedius based on sequence comparison to the genes sodA, pta, hsp60, tuf, and full-length 16S ribosomal DNA, thus demonstrating further species diversity within the SIG.
Keywords
Staphylococcus pseudintermedius, one of the major pathogens involved in canine infections, can be difficult to identify to the species level. 1,3,8 In particular, separation of S. pseudintermedius from the other members of the Staphylococcus intermedius group (SIG; i.e., Staphylococcus intermedius and Staphylococcus delphini) can be challenging. 1,3,8 Members of the SIG share a high average nucleotide identity (ANI) at the 16S ribosomal DNA (rDNA) gene sequence, and identification to the species level has largely been dependent on sequencing of suitable housekeeping genes. 1,8
Species identification and polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) patterns of the 200 beta-hemolysin–producing canine staphylococcal strains investigated.
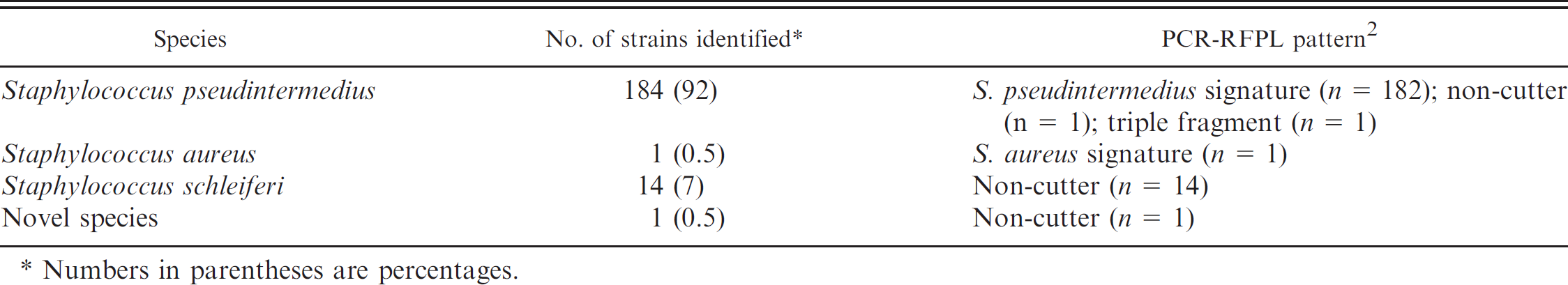
Numbers in parentheses are percentages.
During the last few years, methicillin-resistant S. pseudintermedius (MRSP) in dogs has been reported with increased frequency. Transmission of methicillin-resistant staphylococci between humans and animals has been described, 10 demonstrating a potential zoonotic and public health problem associated with canine staphylococci.
Differentiation between S. pseudintermedius and Staphylococcus aureus also needs attention, as both bacterial species are coagulase and DNase positive and often have a similar appearance on blood agar. It has recently been reported that cefoxitin may be an unsuitable screening marker for methicillin resistance in S. pseudintermedius, 9 emphasizing the need for correct species identification in clinical laboratories that handle samples of both human and animal origin. In a recent proficiency testing of Norwegian hospital laboratories, most of the participating laboratories struggled with the detection of methicillin resistance and species identification of an included MRSP strain (Per Sandven, Norwegian Institute of Public Health, personal communication, 2010).
A rapid and simple polymerase chain reaction–restriction fragment length polymorphism (PCR-RFLP) method for identification of S. pseudintermedius was published in 2009. 2 This method is based on MboI restriction of a pta gene fragment. In the present study, the PCR-RFLP method was used for the species identification of 200 beta-hemolysin–producing Staphylococcus spp. strains associated with canine infections. Complete species identification of all 200 strains was also carried out with the use of supplementary biochemical testing and sequencing of suitable housekeeping genes.
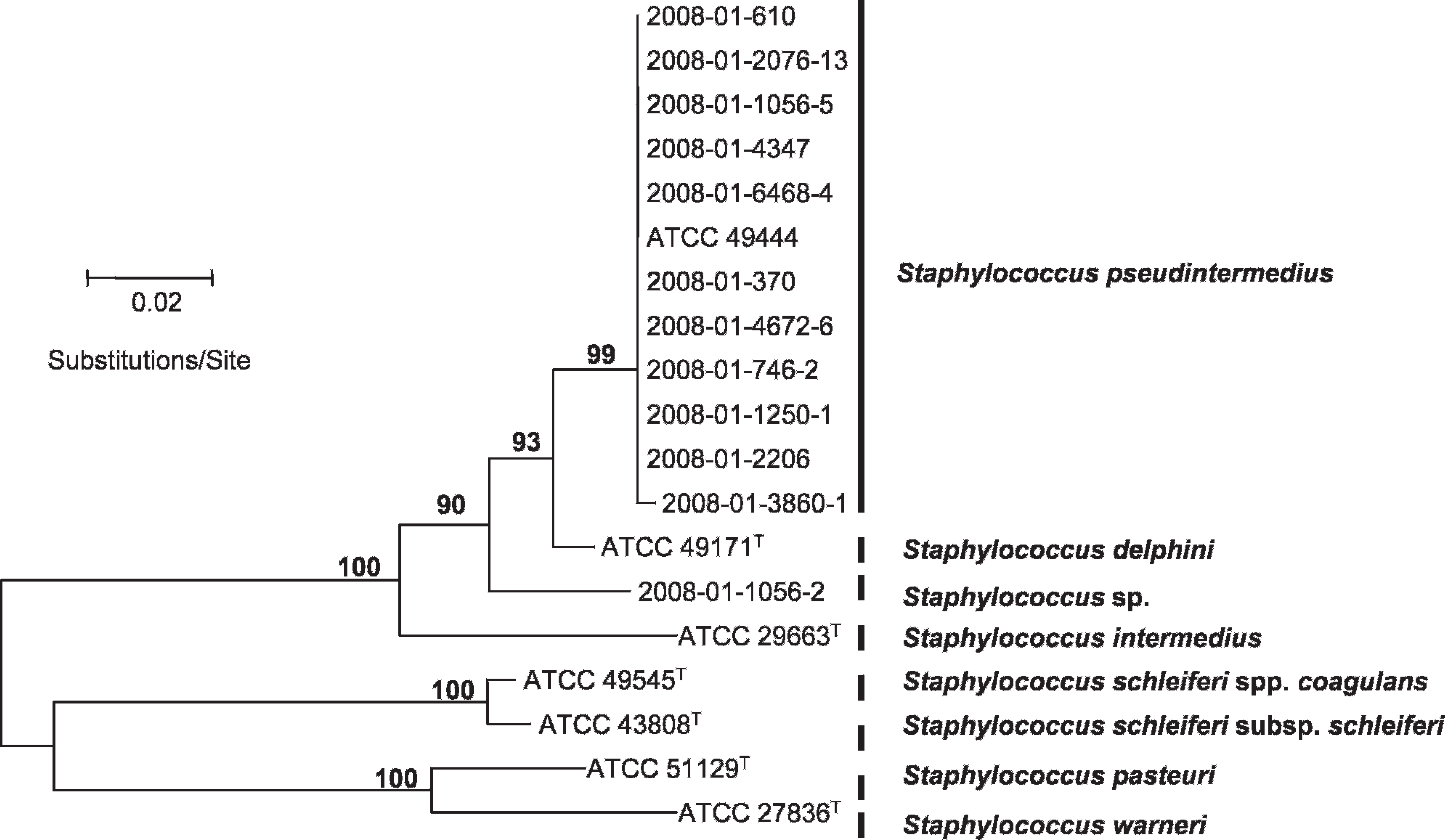
Neighbor-joining and bootstrapping analysis of the sodA gene sequence data; total length of the alignment was 356 characters. Bar = 0.02 substitutions per site. Only bootstrap values larger than 75 are shown.
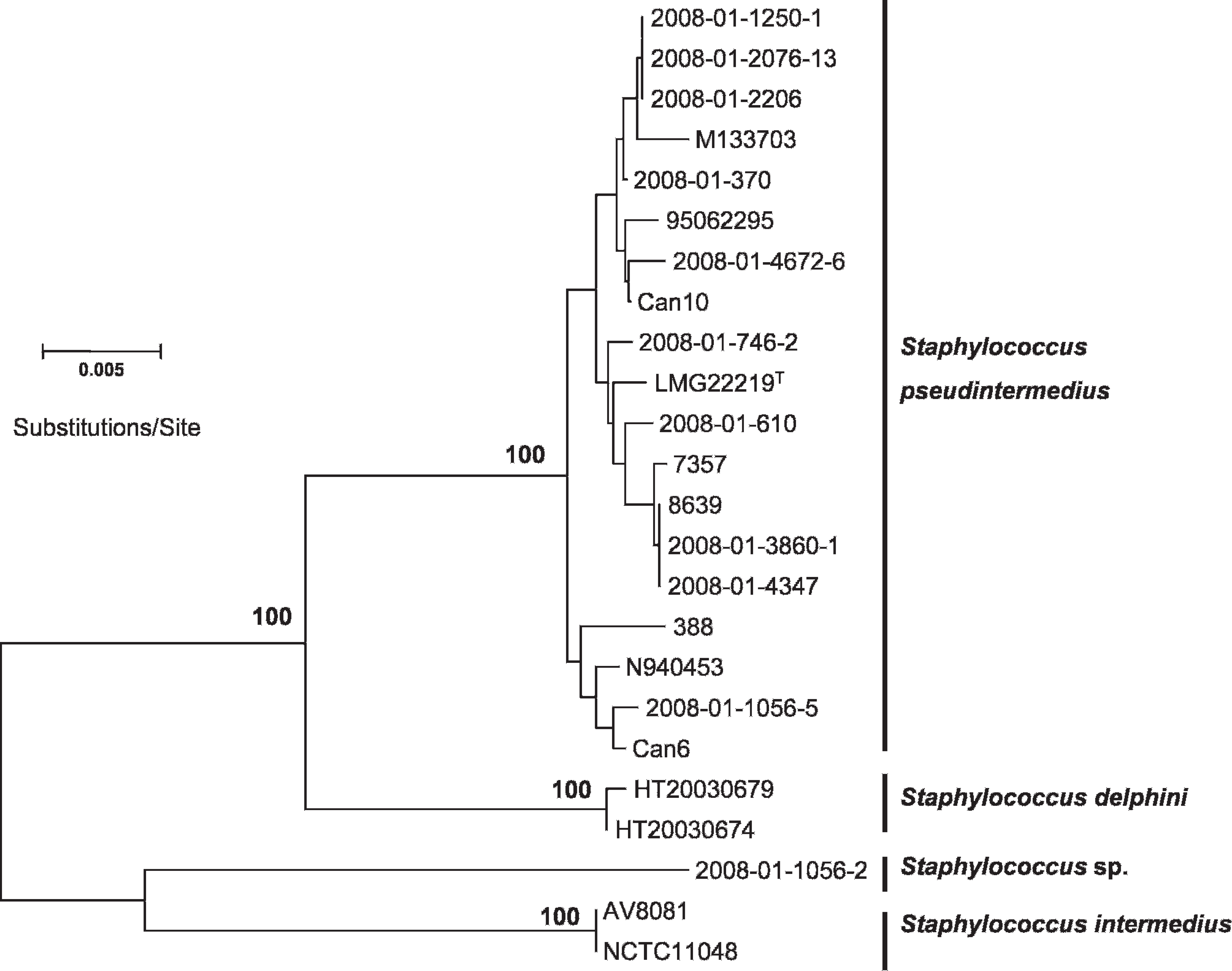
Neighbor-joining and bootstrapping analysis of concatenated alignment consisting of the pta, hsp60, and tuf gene sequences. Total length of the alignment was 1,279 characters. Bar = 0.005 substitutions per site. Only bootstrap values larger than 75 are shown.
The investigated strains were originally included in the Norwegian monitoring program for antimicrobial resistance in the veterinary sector in 2008 (http://www.vetinst.no/eng/Research/Publications/Norm-Norm-Vet-Report/Norm-Norm-Vet-report-2008). Inclusion of isolates was based on positive catalase, arginine dihydrolase, beta-galactosidase activity, sensitivity to acriflavine, and beta-hemolysin production. These inclusion criteria were not sufficient for complete species identification. The isolates were associated with clinical submissions in dogs living in 5 different regions of Norway.
The PCR-RFLP method a , b identified 182 S. pseudintermedius profiles, one S. aureus, 16 non-cutters, and a unique triple fragmented pattern of 47, 111, and 162 base pairs (bp 2 ; Table 1). Fourteen of the 16 non-cutters were identified as Staphylococcus schleiferi subspecies schleiferi or subspecies coagulans 5 based on coagulase, d trehalose, c maltose, c and mannitol c testing and partial 16S rDNA sequencing. e The finding of S. aureus was confirmed by further biochemical testing and testing for resistance to colistin. 3,5 , f
Three isolates, the 2 remaining non-cutters (2008-01-4672-6, 2008-01-1056-2), and the isolate with a unique restriction pattern (2008-01-6468-4) were subjected to further species identification by sequencing of the sodA gene. 7 In addition, sequencing of the genes pta, hsp60, tuf, agrD, and 16S rDNA 1 was carried out. For comparison, 9 randomly selected S. pseudintermedius isolates from the collection of 182 isolates were subjected to the same investigations.
Sequencing of the pta gene fragment revealed G→A or G→T substitution in the MboI site of the 2 non-cutters, and 2 MboI sites were detected in the PCR product from the isolate with a unique restriction pattern. Reference sequences obtained from GenBank were included in alignment studies using BioEdit software. 4 Phylogenetic analyses were carried out with the MEGA4 software with the neighbor-joining method, using the Maximum Composite Likelihood model. 6 Analyses based upon the individual housekeeping genes generated highly similar, but not identical, phylogenetic trees (data not shown). Phylogenetic analyses based on individual and concatenated alignments clearly identified one of the non-cutters and the isolate with the unique restriction pattern as S. pseudintermedius (2008-01-4672-6, 2008-01-6468-4). This finding demonstrates heterogeneity in the MboI site used for S. pseudintermedius species identification. Table 1 presents an overview of identified species and PCR-RFLP patterns of the 200 beta-hemolysin–producing canine staphylococcal strains included in the study.
Finally, the remaining non-cutter (2008-01-1056-2) did not show close relatedness to any of the reference sequences obtained from GenBank. All phylogenetic trees demonstrated this isolate to be a novel member of the SIG (Figs. 1, 2). Polymerase chain reaction with agrD-specific primers failed to generate amplicons of expected size. Sequencing of the 16S rDNA gene (1,535 bp) confirmed a close relationship to the SIG, with an ANI of 99.3% to the S. pseudintermedius type strain LMG 22219T. Concatenated ANI (Fig. 2) identified closest similarity to S. intermedius (95.9%).
The strain was alpha-hemolysin producing on blood agar g containing 5% bovine blood, but was alpha- and beta-hemolysin producing on blood agar containing ovine erythrocytes. This was in contrast to the S. pseudintermedius isolates exhibiting “double” hemolysis due to production of both alpha- and beta-hemolysin on bovine blood agar. The strain was isolated from a dog with an external ear infection. The epidemiological importance of this species is unclear, as it represents a single finding. However, this species may be underrepresented in the current sampling, as either ovine or bovine blood agar was used in connection with the initial selection of isolates. Searching for isolates belonging to this potentially novel species could include the use of both bovine and ovine blood agar, as the appearance on the different agar types could be used as a marker for this species.
Phylogenetic analyses showed that some of the strains were closely related (i.e., 2008-01-1250-1, 2008-01-2206, 2008-01-2076-13 and 2008-01-3860-1, 2008-01-4347; Fig. 2). These strains originated from dogs in geographically distinct regions of Norway, separated by up to 1,000 km, indicating the existence of successful clonal S. pseudintermedius lineages in the Norwegian dog population.
The phylogenetic relationship of all of the 182 isolates demonstrating a S. pseudintermedius PCR-RFLP signature has not been addressed, although 9 of the isolates consistently grouped with S. pseudintermedius. Sequencing of the sodA gene revealed the lowest ANI of the members of the SIG, representing a robust method for identification of species belonging to the SIG. Nevertheless, the PCR-RFLP method should be considered to be a valuable contribution to the species identification of S. pseudintermedius, despite the fact that the method erroneously excludes a small fraction of the population as a result of heterogeneity in the MboI restriction site. The sequences determined in the current study were submitted to GenBank under accession numbers GU057857–GU057895 and GU576850–GU576872.
Acknowledgements
Part of this work was presented in NORM/NORM-VET 2008: Usage of antimicrobial agents and occurrence of antimicrobial resistance in Norway (ISSN: 1502-2307).
Footnotes
a.
Taq DNA polymerase, Qiagen GmbH, Hilden, Germany.
b.
MboI Restriction Enzyme, Fermentas International Inc., Ontario, Canada.
d.
BBL™ Coagulase Plasma, Rabbit with EDTA; BD Diagnostics, Sparks, MD.
c.
BD Diagnostics, Sparks, MD.
e.
Big Dye® Terminator v3.1 Cycle Sequencing Kit, Applied Biosystems Inc., Foster City, CA.
f.
Neo-Sensitabs™, Rosco Diagnostica A/S, Taastrup, Denmark.
g.
Blood Agar Base No. 2, Oxoid Ltd., Basingstoke, Hampshire, United Kingdom.