
Abstract
Four brown pelicans (Pelecanus occidentalis) housed at a rehabilitation facility were found dead after a 3-day history of muscle weakness and after being fed for about 2 weeks from a recent shipment of fish. The birds had pale streaking of the skeletal and heart muscles. Microscopically, the skeletal muscle, and to a lesser extent the cardiac muscle, had severe myocyte degeneration and necrosis characterized by microvacuolation with loss of cross-striations, condensation of cytoplasm, fragmentation, mineralization, and inflammatory cell infiltrates consisting of multinucleated cells, macrophages, and few heterophils. The findings were consistent with myopathy, and a nutritional myopathy caused by eating rancid fish was suspected. Immunohistochemical staining revealed abundant immunoreactive copper zinc superoxide dismutase and manganese superoxide dismutase either as diffuse homogeneous precipitates or granular aggregates in the cytoplasm of affected cells. Immunoreactivity was directly related to degree of cellular damage as estimated by light microscopic examination. We suggest that the lack of protection, despite upregulation of superoxide dismutase, is most likely attributable to supersaturation of oxidants beyond the capacity of superoxide dismutases to scavenge.
Myopathy characterized by degeneration, necrosis, and inflammation is frequently encountered in veterinary medicine. Numerous etiologies have been identified, including vitamin E and selenium deficiencies, ionophore toxicity, various plant toxicities, and inherited genetic defects involving cytoskeletal proteins. 12 Further, exertional or capture stress is well documented as causing severe myopathy in domestic 12 , 14 and wild animals 7 , 8 , 16 , 18 and often ends in death. These etiologies may vary in their microscopic presentation based on the stage of muscle degeneration or regeneration (monophasic vs. multiphasic lesions), which tends to be a direct reflection of exposure to the offending agent. 12
Selenium and vitamin E are important antioxidants intracellularly and within cell membranes, and they protect cells from oxidative stress-induced injury. Vitamin E deficiency may result from dietary deficiency, ingestion of high levels of polyunsaturated fatty acids, 1 ingestion of propionic acid treated grain, 3 or malabsorption diseases of the gastrointestinal tract. 19 The result is oxidant-mediated cell damage, which may manifest in a number of disease entities involving different tissues, such as white muscle disease, mulberry heart disease, encephalomalacia, and steatosis. 4 , 12 The reasons for these diverse presentations are poorly understood; however, the most common manifestation of vitamin E deficiency is myopathy, which has been documented in mammals and birds. 12 , 17
Superoxide dismutases (SODs) are a group of enzymes that efficiently degrade superoxide radicals. They are hypothesized to play a significant role in preventing oxidant-mediated cell damage. Three specific enzymes, mitochondrial manganese SOD (MnSOD), intracellular copper-zinc SOD (CuZnSOD) and extracellular SOD, have been described based on their specific distributions and functions. 9 Immunohistochemical staining for these enzymes has been done to determine their activity in various disease states. 11 , 13 , 20
In January 2005, a group of 10 captive, nonreleasable, brown pelicans (Pelecanus occidentalis) displayed clinical signs of muscle weakness while remaining bright and alert and having normal appetites. There was no history of recent capture stress. One bird died and fresh and fixed sections of skeletal muscle (multiple locations), heart, liver, kidney, testicle, and lung were submitted. Approximately 8 days later and 3 days after the onset of clinical signs, 3 additional birds (1 adult female, 1 adult male and 1 subadult male) were found dead and submitted for necropsy. The rehabilitation facility reported a recent shipment of fish that was the primary food source for the birds for about 1 week before initial onset of clinical illness. These fish were not fed to other animals in the facility. This report describes the gross and microscopic changes found in these birds, along with patterns of immunohistochemical evidence for expression of antioxidant enzymes in the affected muscles.
Whole birds and submitted tissues were examined grossly. Whole birds were necropsied and 2 sets of tissues, including muscle, brain, heart, lung, liver, kidney, skin, spleen, gastrointestinal tract, and reproductive organs, were collected. One set of tissues was frozen (-80°C) for possible future testing, pending histologic examination. The other set of tissues was collected in 10% buffered formalin, routinely processed, and embedded in paraffin; then, 5-μm sections were cut, placed on glass slides, and stained with hematoxylin and eosin for light microscopic examination. Tissue sections submitted from the first pelican were similarly processed.
Immunohistochemical staining using commercially available polyclonal antibodies a against CuZnSOD and MnSOD was performed. Sections were cut from the tissue blocks (as already described), placed on positively charged glass slides, deparaffinized, rehydrated, and treated for 15 minutes with 3% hydrogen peroxide to inhibit endogenous peroxidase. This was followed by incubation with the respective primary sheep antibody at room temperature for 1 hour. After washing with phosphate-buffered saline (PBS, pH 7.4), the sections were incubated with biotinylated anti-immunoglobulin. The sections were again washed with PBS and treated with avidin-biotin complex reagent. b The immune complexes were visualized as a brown product using diaminobenzidine as chromogen. The slides were counterstained with Mayer hematoxylin. Parallel sections were simultaneously stained following the protocol described but omitting the primary antibodies; these served as the negative controls. Sections of canine heart, used to optimize the protocol, served as a positive control. These antibodies have not previously been tested in avian species. Therefore, sections of turkey heart and skeletal muscle were used in parallel and served as a positive avian control.
All 4 pelicans had similar changes. The whole birds had healed amputated wings and were in fair flesh; the males had fish in their gastrointestinal tracts. There was generalized pallor of the body musculature. The pectoral muscle had expanded bands of pallor surrounded by normal-appearing muscle (Fig. 1A). Muscles in the legs and wings also had similar pale bands. Linear multifocal pale slender streaks were randomly dispersed in the myocardium (Fig. 1B).
Microscopic changes in the muscles were severe and diffuse. Loss of cross-striations, myocyte micro- and macrovesiculation, myofiber fragmentation, and necrosis were widespread (Fig. 1C). Virtually no fiber was spared. Multicentric mineralization, stellate cell proliferation, influx of histiocytic cells and multinucleated giant cells were frequently present, altering the integrity of the muscle. Perimysial fibrosis, myofiber regeneration, and segmental loss of myocytes were common. The myocardial changes were similar to skeletal muscle changes but were less intense.
Intracytoplasmic dark brown granular material was stained by the MnSOD polyclonal antibody (Fig. 1D). Myocytes, macrophages, multinucleated giant cells, fibro-blasts, and endothelial cells within the affected muscles were positive for MnSOD. The expression was directly proportional to the degree of myocyte injury. The CuZnSOD expression level was more intense and homogeneous intracellularly (Fig. 1E). Although the level of CuZnSOD expression was directly related to the degree of damage, immune-complexes in the form of dark brown patches were more intense peripherally in the myocytes.
Histiocytes and giant cells within the affected muscles had diffuse strong CuZnSOD immunorecognition in the cytoplasm.
Both cardiac and skeletal muscles are sensitive to oxidative-stress-induced damage. All 4 pelicans in the present study likely were exposed to the same etiologic factor that led to the generalized myopathy. In a recent report, rancid diet with high peroxide contents has been ascribed as the potential causative factor for muscular dystrophy. 17 Rancid diet decreases the digestibility of nutrient and thus precipitates myopathy and death. 5 , 6 Oxidative phosphorylation constantly generates reactive oxygen species (ROS) that are deleterious to cells and are scavenged by a group of antioxidant molecules. These ROS are involved in peroxidation of membrane lipid, leading to cell injury and death. Antioxidant vitamins, particularly vitamin E, are engaged intracellularly and at the plasma membrane level to terminate ROS-induced cellular damage. Like vitamin E, selenium also exerts an antioxidant action and protects from muscular dystrophy. However, the relative importance of vitamin E and selenium in preventing myopathy is not clearly understood. Although intake of vitamin E can minimize the requirement of selenium and vice versa, the 2 nutrients are not completely complementary. In pastured cattle, nutritional myopathy can be prevented by the addition of selenium but calves fed a cod liver oil-enriched diet are not protected and develop a myopathy. 4
In the present study, blood samples or feed samples were not available for laboratory analysis. The fish found in the stomachs of the whole birds were not from the suspected rancid shipment, as a new shipment was acquired and fed once diagnosis was made on the initial submission. However, it seems very likely that the birds had access to rancid fish and succumbed because of a myopathy, as has been reported in the literature. 5 , 17 Further, the remaining live birds were supplemented with the antioxidant vitamin E, which resulted in recovery and no additional mortalities, similar to a previously reported case. 17
Superoxide dismutases are a unique class of intracellular antioxidants that convert superoxide radicals to hydrogen peroxide. High expression of CuZnSOD and MnSOD in the myocytes in the present study is a typical response to oxidants. In spite of high expression, both CuZnSOD and MnSOD failed to avert the cellular damage. The most likely explanation is that the overwhelming quantity of oxidants was beyond the ability of the antioxidant enzymes to scavenge, thus precipitating cell death. Additionally, the rate of oxidant generation likely was so rapid that it was beyond the physiologic functional capacity of these enzymes. A less plausible explanation is that, similar to what has been previously reported, 2 , 10 , 15 antioxidant enzymes alone may not always be enough to confer protection. This study demonstrates severe generalized myopathy with expression of abundant antioxidant enzymes in the damaged muscle tissue in pelicans. To elucidate the mechanism of myodegeneration in oxidant induced myopathy, systematic studies using known oxidants are necessary.
Acknowledgements. We wish to thank the staff of The University of Georgia, College of Veterinary Medicine, Tifton Veterinary Diagnostic and Investigational Laboratory, especially Lisa Whittington, for assistance in tissue processing.
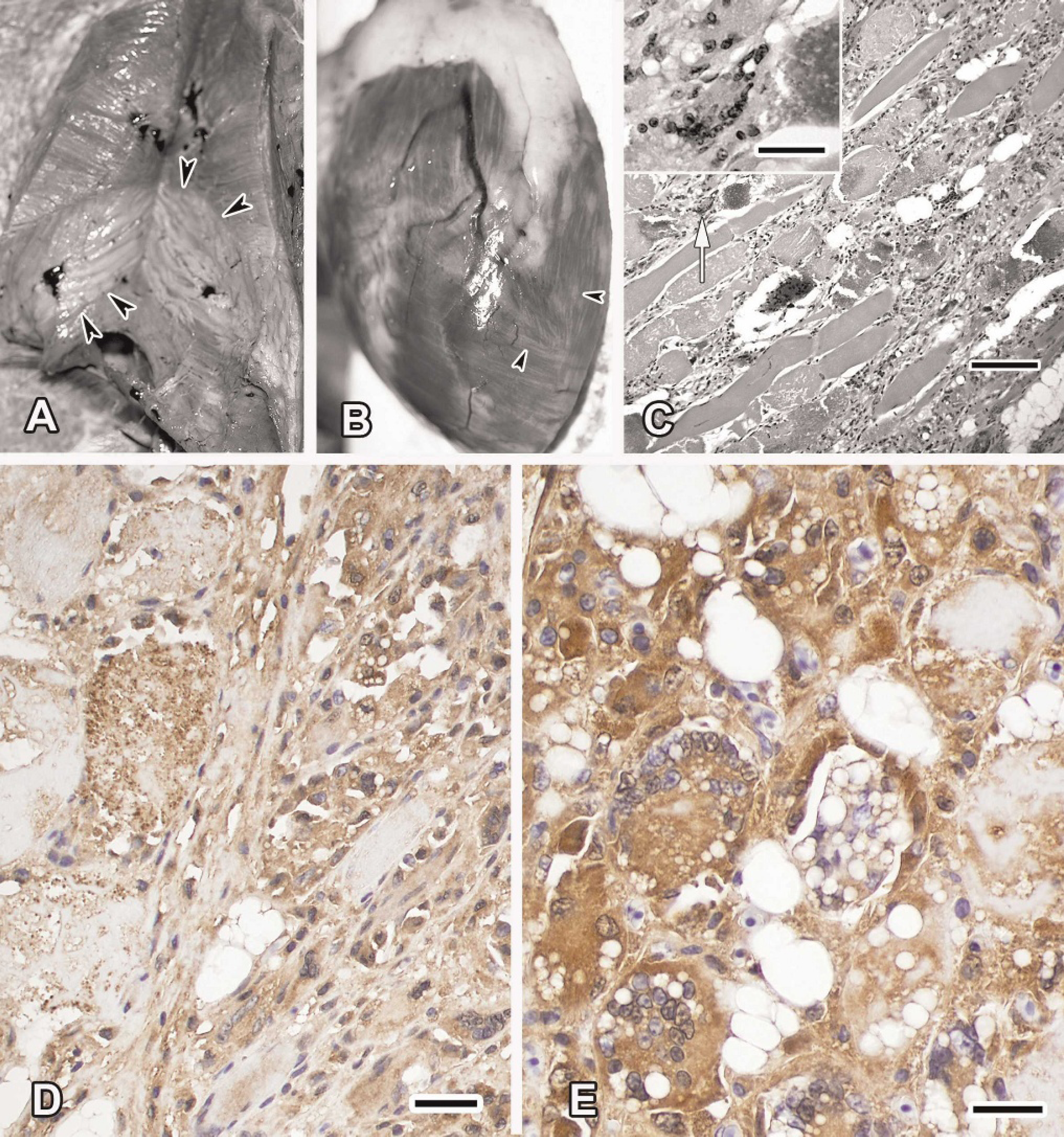
Muscle of a brown pelican (Pelecanus occidentalis) that died from nutritional myopathy.
Footnotes
a.
Anti-Superoxide Dismutase (Mn Enzyme), Human Liver (Sheep) and Anti-Superoxide Dismutase (Cu/Zn Enzyme), Human (Sheep), Calbiochem, San Diego, CA.
b.
DAKO, Carpinteria, CA.