
Abstract
Protein arginine methyltransferase 5 is one of the type II protein arginine methyltransferase family members that can symmetrically dimethylate arginine residues on target proteins in both the cytoplasm and the nucleus. Protein arginine methyltransferase 5 was reported to be an oncoprotein that participates in tumor progression through both epigenetic silencing and organelle biogenesis. So far, it has been implicated in various cancers, but its expression pattern in breast cancer has not been elucidated thoroughly. We analyzed the protein arginine methyltransferase 5 expression patterns in several breast cancer samples and tissue arrays to better characterize its contribution to breast cancer. Primary breast tumors showed increased protein arginine methyltransferase 5 expression compared with adjacent normal tissues in both the fresh tissue samples and tissue arrays. Also, there was a tendency that metastatic lymph nodes demonstrated enhanced protein arginine methyltransferase 5 expression compared to primary sites. Moreover, we found a significant correlation between protein arginine methyltransferase 5 and Ki-67, with higher Ki-67 and protein arginine methyltransferase 5 expressions in primary breast tumors compared with normal breast tissues. Moreover, the Cancer Genome Atlas cohort analysis revealed that high protein arginine methyltransferase 5 messenger RNA expression was associated with an unfavorable prognosis in human epidermal growth factor receptor 2 (HER-2) positive and triple negative breast cancer patients. Finally, the roles and mechanisms of protein arginine methyltransferase 5 in the proliferation, cell cycle progression, and apoptosis of MDA-MB-231 cells were assessed using protein arginine methyltransferase 5 and shPRMT5 transfection. In conclusion, we proposed that protein arginine methyltransferase 5 is an independent prognostic biomarker for breast cancer, and targeting protein arginine methyltransferase 5 might be a promising strategy for breast cancer treatment.
Keywords
Introduction
Breast cancer, the most commonly diagnosed cancer in women, has been considered as the leading worldwide cause of fatality among female patients.1,2 In China, breast cancer has become one of the most common cancers, with more than 1.6 million people being diagnosed and 1.2 million people dying each year, accounting for 12.2% of all newly diagnosed breast cancers and 9.6% of all breast cancer–related deaths worldwide. 3 Prognostic factors are frequently used for making clinical decisions in breast cancer, such as the histological types of the tumor, hormone receptor status, and other prognostic biomarkers. 4 Recently, accumulating studies have identified the gene signatures of breast cancer, and explored the biomarkers of breast cancer prognosis, leading to more accurate clinical decisions for cancer patients. However, barriers in the early diagnosis and treatment of breast cancer still exist because of the differences in the genetic nature and response to therapy. 5 There is still an urgent need to explore novel targets to indicate the outcomes of breast cancer.
Protein arginine methyltransferase 5 (PRMT5) exists universally in the nucleus and cytoplasm and is one of the type II PRMT enzymes, which covalently modify histone and non-histone proteins and maintain numerous cellular regulatory networks.6,7 As reported, PRMT5 can symmetrically dimethylate arginine residues on target proteins and is involved in tumor progression via both epigenetic silencing and organelle biogenesis.8,9 The overexpression of PRMT5 has been detected in various types of carcinomas, including ovarian, 10 glioblastoma multiforme, 11 melanoma, 12 colon, 13 bladder cancer, 14 and germ cell tumors. 14 Increased proliferation, induced anchorage-independent colony growth, as well as tumorigenesis were found when PRMT5 is overexpressed; therefore, PRMT5 has been considered a promising target for cancer therapy.15–17 Recently, several studies indicated that PRMT5 plays important roles in breast tumorigenesis.18–21 It is reported that PRMT5 binds to the zinc finger structures in tumor necrosis factor receptor–associated factor 4 (TRAF4) and is regulated by this scaffold protein; TRAF4 participates in nuclear factor (NF)-κB signaling and is involved in the promotion of cell proliferation and survival in breast cancer. 18 Programmed cell death 4 (PDCD4) is a tumor suppressor, but overexpressed PRMT5 can negate PDCD4 tumor suppressor function via methylating PDCD4 at residue R110; then, they work synergistically to promote breast tumor growth in the xenograft model. Also, breast cancer patients who highly expressed both PDCD4 and PRMT5 had worse outcomes compared with those who had high PDCD4 and low PRMT5, who had better outcomes. 19 DAL-1/4.1B is a growth suppression protein which is involved in breast tumorigenesis. 22 It is found to regulate PRMT5 by inhibition (Sm proteins), as well as enhancing (myelin basic protein) protein methylation, which is biologically important in the control of breast pathogenesis. 20 PRMT5-methylosome protein 50 (MEP50) is essential for regulating the transcription, promoting the invasive abilities of breast carcinoma cells, and maintaining the expression of metastatic and epithelial-to-mesenchymal transition (EMT) markers. Transforming growth factor (TGF)-β can facilitate EMT via PRMT5-MEP50-catalyzed histone methylation of chromatin at key metastasis suppressor and EMT genes. 21 However, the biological role and clinical significance of PRMT5 in breast carcinoma have not been elucidated thoroughly and should be further investigated.
In this study, we explored the expression of PRMT5 in normal breast tissues, primary breast tumor tissues, and metastatic tissues by immunohistochemistry and western blot. Moreover, the relationship between PRMT5 expression and breast cancer patient prognosis was analyzed, and we found that the overall survival (OS) in patients with high PRMT5 expression was shorter than that of patients with low PRMT5 expression. The effect of PRMT5 on breast cancer cell proliferation, cell cycle progression, and apoptosis was assayed using the overexpression and knockdown of PRMT5 in MDA-MB-231 cells. Furthermore, we found that PRMT5 contributed to breast cancer cell proliferation partly via regulating extracellular signal–regulated protein kinase (ERK) phosphorylation, and PRMT5 may regulate MDA-MB-231 cell apoptosis through regulating the Bcl-2/caspase 3/poly (ADP-ribose) polymerase (PARP) pathway. Our findings indicate that the overexpression of PRMT5 could be identified as a biomarker of poor prognosis for breast cancer.
Materials and methods
Specimens and tissue arrays
All the samples of human breast cancer tissues were obtained from our department between 2015 and 2016. The tissues were snap frozen in liquid nitrogen and stored at −80°C until use. The use of human tissues was reviewed and approved by the Ethics Committee of Xijing Hospital. Prior to the study, all patients gave their written informed consent to allow us to use the leftover tissue samples for scientific research.
Two sets of breast tissue arrays (1604A.B and BC-001A.B) were gifted from the Department of Pathology, Xijing Hospital, The Fourth Military Medical University. One included normal breast tissues and primary breast tumors, and the other included primary and metastatic breast tumors. They were obtained from patients treated by breast tumor resection between 2015 and 2016 at the Xijing Hospital. The patients provided informed consent, and the study was approved by the Ethics Committee of Xijing Hospital. Areas that represented primary breast cancer, normal breast tissue, and metastatic lymph nodes were marked on hematoxylin/eosin-stained sections. In addition, 1.60 mm tissue cores were extracted and mounted; then, the cores were transferred to tissue microarrays and cut into 5 µm sections and placed in a water bath at 50°C. Finally, the slides were dried at room temperature and stored in a freezer at −80°C until use. To minimize the loss of antigenicity, the slides were used within 4 months.
Western blot analysis
Tissue specimens were homogenized and lysed in radioimmunoprecipitation assay (RIPA) lysis buffer (100 mM NaCl, 50 mM Tris-HCl pH 7.5, 1% Triton X-100, 1 mM ethylenediaminetetraacetic acid (EDTA), 10 mM β-glycerophosphate, 2 mM sodium vanadate, and 1 mM protease inhibitor). The protein concentration was assayed using the micro BCA protein assay kit (Thermo Fisher Scientific, Rockford, IL, USA). Equal amounts of protein extracts were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and transferred onto polyvinylidene fluoride (PVDF) membranes (Merck Millipore, Bedford, MA, USA), which was blocked with 5% non-fat milk in 0.1% Tween-20/Tris-buffered saline (TBS) and incubated with a PRMT5-specific primary antibody (1:1000; Abcam, Cambridge, UK) and α-tubulin (1:1000; Cell Signaling Technology, Danvers, MA, USA) overnight at 4°C. The membrane was then incubated with a secondary antibody conjugated to horseradish peroxidase (HRP; 1:2000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) and developed using a chemiluminescent substrate (PerkinElmer, Waltham, MA, USA) according to the manufacturer’s recommendations, and the resulting membranes were exposed on a Kodak X-Omat processor (Rochester, NY, USA).
Immunohistochemistry
The methods were carried out in accordance with the approved guidelines, and all experimental protocols were approved by a named institutional and/or licensing committee. After deparaffinization with xylene, the tissue sections were sequentially rehydrated through gradient ethanol immersion and then washed three times with phosphate-buffered saline (PBS). For antigen retrieval, tissue slices were immersed in boiled citrate–disodium hydrogen phosphate buffer (pH 6.0) with high pressure for 5 min. Then, a 3% hydrogen peroxide (H2O2) solution was added to the naturally cooled sections for 15 min to quench the endogenous peroxidase activity. The sections were then blocked with normal goat serum, followed by incubation with polyclonal rabbit anti-human PRMT5 antibody (1:100) and Ki-67 (1:50; Proteintech Group, Chicago, IL, USA) antibody overnight at 4°C.
The sections were incubated with biotinylated secondary antibody and streptavidin–peroxidase (Invitrogen, Grand Island, NY, USA). For color development, fresh 3,3′-diaminobenzidine (DAB) solution was added to the tissue slices, followed by counterstaining with hematoxylin. As negative controls, adjacent sections were processed as described above, except that they were incubated in blocking solution without the primary antibody overnight at 4°C. After washing with PBS, slides were incubated for 25 min at room temperature with biotinylated goat anti-rabbit immunoglobulin followed by incubation with peroxidase conjugated streptavidin for 20 min and 0.05% fresh DAB. Slides were counterstained with Mayer’s hematoxylin and mounted with crystal mount.
Evaluation and quantification of immunostaining
It has been reported that PRMT5 promoted tumor proliferation, 23 and we analyzed the expression of Ki-67 on the same set of tissue arrays, which was used as an indicator of tumor proliferative capacity. Two pathologists blinded to the clinical data evaluated the staining of each specimen. To avoid inter-observer variability, the mean value of the scores was adapted for further analysis. For PRMT5, the assessment of expression levels included the staining intensity and the percentage of stained cells. PRMT5 was analyzed for both nuclear and cytoplasmic staining. The staining intensity was scored as 0 = no staining, 1 = mild, 2 = moderate, and 3 = strong expression; the results were categorized according to the following distribution: 0 = <6%, 1 = 6%–20%, 2 = 21%–50%, and 3 = >50% staining. The PRMT5 expression score was determined as a combined score of staining intensity and distribution. Samples with a final immunoscore ≥4 were considered as having a high expression of PRMT5. 10
Ki-67 detection
As for measuring Ki-67 expression, nuclei from at least 1000 cells were counted from the fields, and the labeling index was calculated as the percentage of labeled nuclei of the total number of cells that were counted. The median was used to dichotomize into low- and high-expressing groups for Ki-67 for statistical analysis, 24 which was 20%. 23
Analysis of PRMT5 expression in the Cancer Genome Atlas samples correlation with outcomes
The expression level of PRMT5 was investigated in The Cancer Genome Atlas (TCGA) breast cancer samples (998 cancer samples and 108 normal samples) in starBase v2.0 (http://starbase.sysu.edu.cn/index.php). 25 Prognosis analysis was performed in 26 datasets with 3554 breast cancer patients in Gene Expression Omnibus (GEO) database (Supplementary data 1). The raw CEL files were downloaded from the GEO database (http://www.ncbi.nlm.nih.gov/geo/), and the background was adjusted according to the previously described method. 26 The expression of PRMT5 was dichotomized using study-specific median expression as the cutoff to define “high value” as above the median versus “low value” as below the median. The Kaplan-Meier method and log-rank test were performed to show survival differences according to PRMT5 expression. The time for OS or relapse free survival (RFS) was calculated as the time from surgery until the occurrence of death and relapse, respectively.
Cell transfection
The PRMT5, shPRMT5, and negative control (NC) lentiviral vectors were gifted by Jiaji Lin of Department of Neurology, Tangdu Hospital, and transfected with Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) into MDA-MB-231 cells according to the manufacturer’s instructions. MDA-MB-231 cells were grown on six-well plates to confluency, and at 48 h post-transfection, cells were harvested for western blot for the detection of transfection efficiency.
MTT assay
The in vitro cell survival ability of MDA-MB-231 cells post-transfection was measured using the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. After 24 h of transfection, MDA-MB-231 cells were seeded on 96-well plates at 8000 cells/well density and incubated at 37°C in a humidified atmosphere with 5% CO2. Spent cell culture medium was replaced with 0.2 mL of fresh medium containing 0.5 mg/mL MTT every 24 h. Cells were then incubated at 37°C for 4 h and resolved using 0.15 mL of dimethyl sulfoxide (DMSO). The absorbance for each well was measured at 490 nm (A490) using a spectrophotometer. Three independent experiments were performed in quintuplicate.
Flow cytometry for apoptosis and cell cycle analysis
Cells transfected with PRMT5, shPRMT5, or NC for 72 h were harvested. For the cell cycle analysis, cells were washed with ice-cold PBS twice and were fixed in ice-cold 70% ethanol overnight at −20°C. Fixed cells were resuspended in flow cytometry buffer and were subjected to propidium iodide (PI)/RNase staining. Flow cytometry was performed using a FACScan instrument (Becton Dickinson, Franklin Lakes, NJ, USA), and analysis was performed with CellQuest software (Becton Dickinson). For cell apoptosis analysis, the cells were washed with cold PBS; then, an Annexin V-PE Apoptosis Detection Kit (Oncogene Research Products, Boston, MA, USA) was used to measure apoptotic cells according to the manufacturer’s instructions, and the cells were analyzed using flow cytometry (Becton Dickinson).
Statistical analysis
Statistical analyses were performed with SPSS 19.0 software (SPSS, Chicago, IL, USA). Cumulative survival time was calculated by the Kaplan-Meier method and analyzed by the log-rank test. For the comparison of individual variables, t tests, χ2 tests, and one-way analysis of variance (ANOVA) were carried out as appropriate. Statistical significance was determined for two-tailed tests at p < 0.05.
Results
Upregulated expression of PRMT5 in breast cancer
Using western blots as described above, we examined the expression profiles of PRMT5 in breast cancer tissue samples and in the adjacent normal tissues resected from patients diagnosed with primary breast cancer, including one bilateral synchronous breast cancer. PRMT5 expression levels were found to be increased in 25 of 31 (80.6%) tumor lesions (T) compared with the matched normal tissue (N); in the other 6 pairs (19.4%), the levels of PRMT5 in normal tissues were higher than tumor tissues, and some of the results are shown in Figure 1.
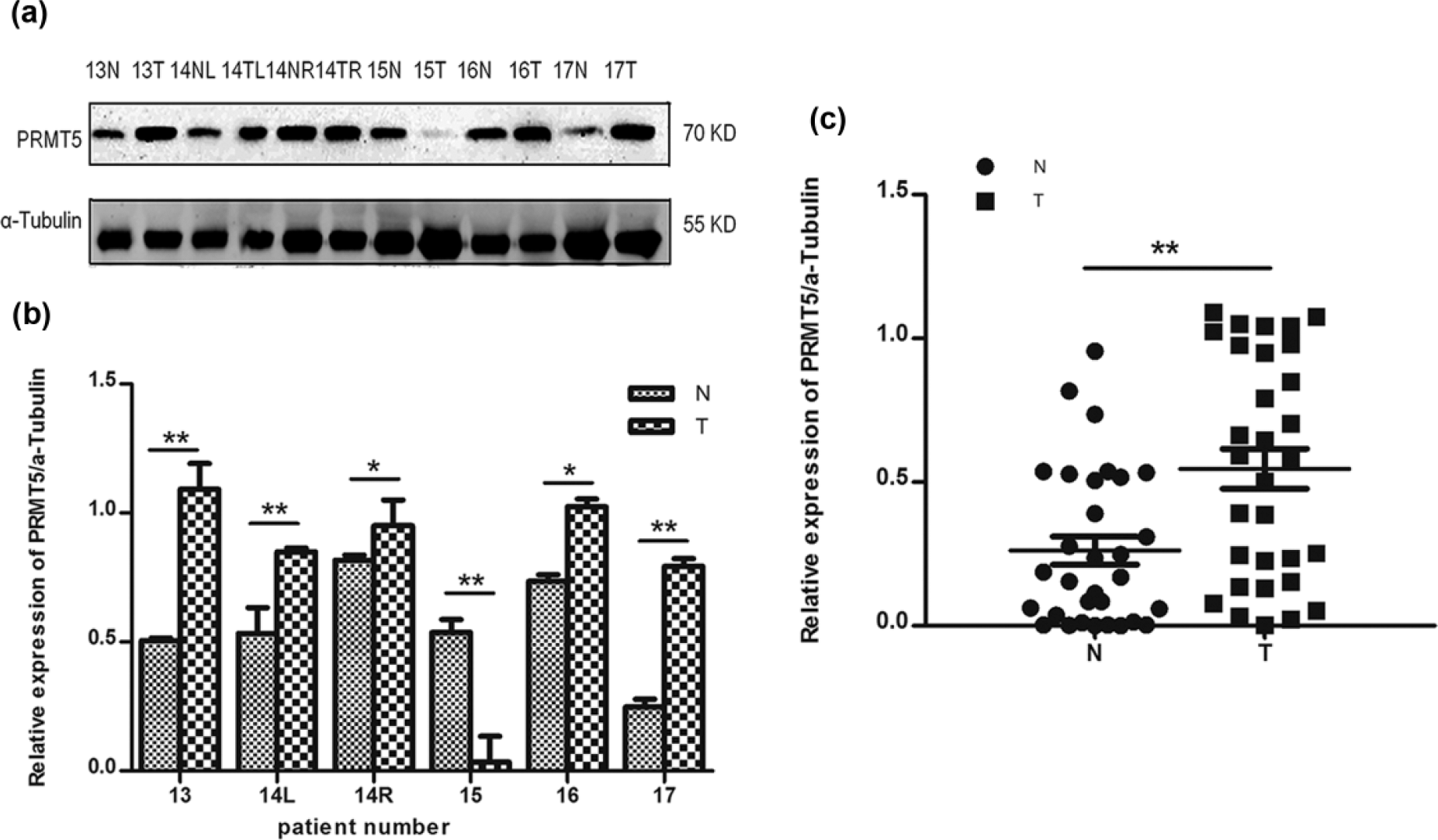
PRMT5 expression in breast tumors and adjacent normal breast tissues: (a and b) Western blot analyses showing that PRMT5 expression is higher in some breast tumor tissues than adjacent normal breast tissues. (c) PRMT5 protein expression levels are higher in breast cancer tissue (T) than in the paired normal tissue (N) (n = 31).
The expression of PRMT5 was also assessed using immunohistochemistry from two separate tissue microarrays. Normal breast tissues, primary breast tumors, and metastatic breast tumors were included in the two tissue arrays. Positive staining for PRMT5 was observed in both the nucleus and cytoplasm (Figure 2; Table 1). According to the criteria described previously, in the first tissue array, 1 of 26 (3.8%) normal breast tissues and 23 of 40 (57.5%) primary breast tumors showed high PRMT5 expression. The level of PRMT5 in primary breast tumors was statistically higher than that in normal breast tissues (p < 0.01). In another array, high PRMT5 expression in 19 of 35 (54.3%) primary breast tumor tissues and 25 of 34 (73.5%) metastatic breast tumors was detected (p > 0.05; Figure 2). The correlation analysis of PRMT5 expression with the clinical pathological features of primary and metastatic breast cancer patients is shown in Supplementary Table 2.

Immunohistochemical staining for PRMT5 and Ki-67: (a) PRMT5 staining on normal breast tissues. (b and c) Exemplary image of PRMT5 staining on primary and metastatic breast cancer tissues showing cytoplasm and nuclear staining pattern. (d) Ki-67 staining on normal breast tissues. (e and f) Exemplary image of Ki-67 staining on primary and metastatic breast cancer tissues with nuclear positive only.
Pattern of PRMT5 localization in normal breast tissues and cancer tissues.
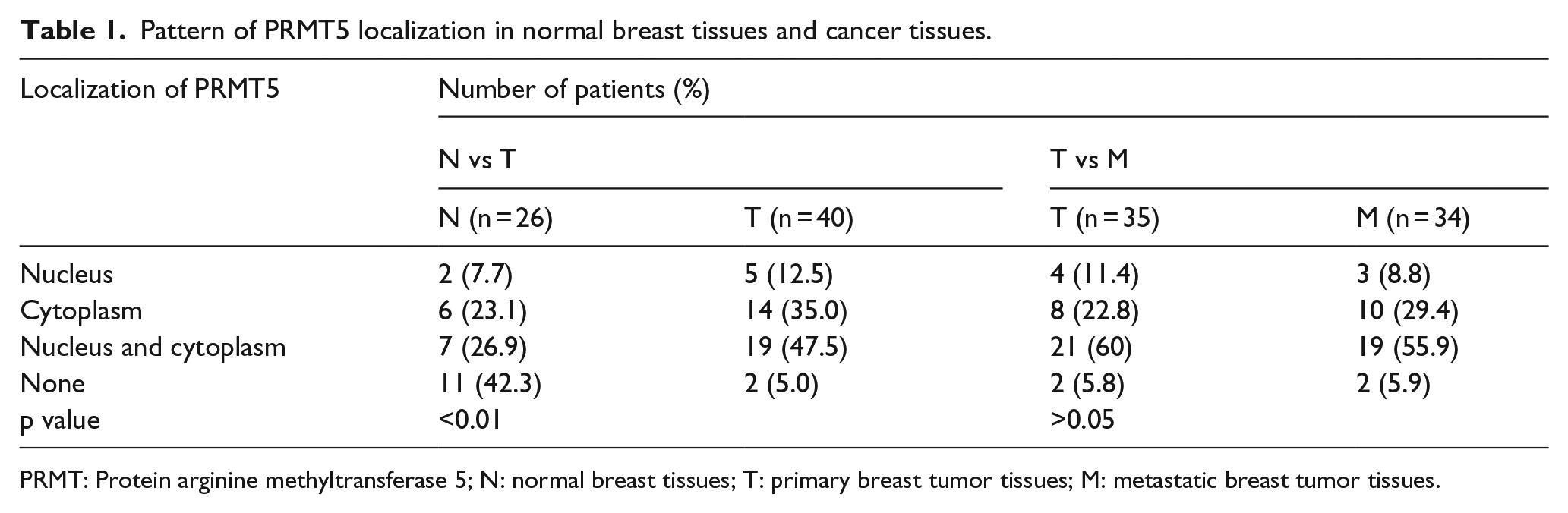
PRMT: Protein arginine methyltransferase 5; N: normal breast tissues; T: primary breast tumor tissues; M: metastatic breast tumor tissues.
Relationship of PRMT5 and Ki-67 in breast cancer tissue specimens
There was a significant positive relationship between Ki-67 and PRMT5 expression; samples with high Ki-67 had significantly higher PRMT5 expression in primary (70.6% vs 35.3%, p < 0.05) and metastatic breast tumors (94.4% vs 56.25%, p < 0.05; Figure 2) compared with the samples with low Ki-67 expression in primary and metastatic breast tumors.
Correlation of PRMT5 with breast cancer metastasis
We proved that PRMT5 levels were higher in primary breast tumor tissues than in the paired normal tissues; next, we wanted to investigate whether PRMT5 was correlated with breast cancer metastasis. To answer this question, seven primary breast tumor tissues, matched normal tissues, and the corresponding metastatic lymph nodes were used to analyze the expression of PRMT5 by western blot. The results showed that PRMT5 expression was higher in all seven pairs of tumor samples than the normal tissues. Also, as shown in Figure 3, four of seven (57.1%) cases expressed enhanced PRMT5 protein levels in metastatic lymph nodes compared with primary breast tumors. These data suggested that PRMT5 might participate in breast tumorigenesis and metastasis.

Western blot analysis of PRMT5 expression in breast tumors, adjacent normal breast tissues, and corresponding metastatic lymph nodes: (a and b) Western blot analyses showing that PRMT5 expression is higher in some breast tumor tissues than in adjacent normal breast tissues, and even higher in metastatic lymph nodes. (c) PRMT5 protein expression levels were higher in breast cancer tissues (T) and metastatic lymph nodes than in the paired normal tissues (N) (n = 7).
Correlation of PRMT5 with breast cancer survival
To better understand the clinical significance of PRMT5, a gene expression profile of PRMT5 was obtained from the TCGA database to analyze the survival outcomes. We analyzed the expression of PRMT5 in 998 primary tumors and 108 normal samples and found that the normal samples expressed less PRMT5 messenger RNA (mRNA; Student’s t test p = 0.000400302; Figure 4(a)). With the follow-up period ranging from 0 to 250 months, breast cancer patients with higher PRMT5 expression had significantly decreased RFS time (hazard ratio (HR) = 1.73 (1.47–2.05); log-rank p = 3.7e−11; Figure 4(b)). Figure 4(c) demonstrated that in human epidermal growth factor receptor 2 (HER-2)-positive breast cancer, PRMT5 expression was inversely correlated with patients’ OS time, and the difference between the survival curves of the two groups was statistically significant (HR = 0.39 (0.16–1); log-rank p = 0.043). In the triple negative breast cancer (TNBC) subgroups, PRMT5 expression was a significant predictor of OS time. TNBC patients with higher PRMT5 levels showed decreased OS time compared with those with lower PRMT5 levels. The numbers of TNBC patients at risk at the cumulative 0, 50, 100, and 150 months in the higher PRMT5 expression group were 63, 26, 6, and, 1. In the lower PRMT5 expression group, all of which were decreased compared to the numbers of those at risk of 69, 41, 13, and 1, respectively, in the lower PRMT5 expression group (p < 0.01; HR = 2.53 (1.15−5.56); log-rank p = 0.017; Figure 4(d)). The difference in PRMT5 expression in luminal-A and luminal-B tumors was not statistically correlated with patients’ OS time.
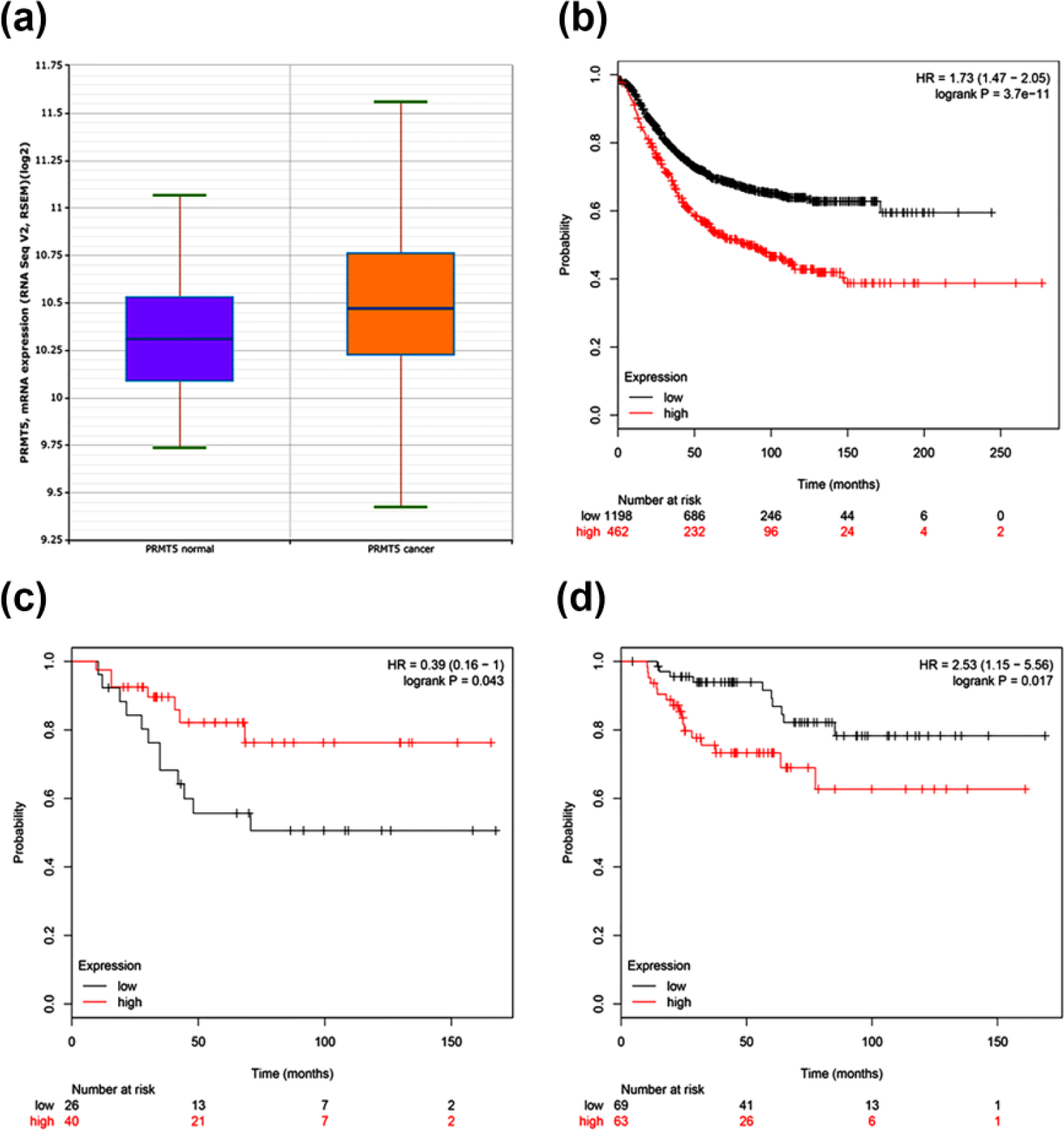
Overexpression of PRMT5 in breast cancer is negatively correlated with patient survival: (a) Analysis of PRMT5 expression status in breast cancer patients and normal people. (b) Correlation analysis of PRMT5 expression status in the breast cancer patients and RFS. (c) The surviving curves of HER-2 positive breast cancer patients with high and low PRMT5 expression. (d) Correlation analysis of PRMT5 expression status in the TNBC patients and patient survival.
PRMT5 expression affects MDA-MB-231 cell proliferation ability in vitro
To assess the role of PRMT5 in breast cancer cells, breast cancer MDA-MB-231 cells were transfected with PRMT5, shPRMT5, or NC, and western blot showed that PRMT5 expression was significantly upregulated in MDA-MB-231 after transfection with PRMT5 and downregulated after transfection with shPRMT5 (Figure 5(a), (b), (d), and (e)). We investigated the effect of the change of PRMT5 expression on cell proliferation. An MTT assay was performed, and it revealed that cell growth was promoted in MDA-MB-231-PRMT5 cells, whereas it was impaired in MDA-MB-231 cells transfected with shPRMT5, compared with control vector-transfected cells (Figure 5(c) and (f)).

(a, b, d, and e) Association of PRMT5 with the proliferative potential of breast cancer cells. (c and f) Detection of PRMT5 expression by western blot. MTT assay was performed to determine the proliferation of PRMT5-, shPRMT5-, or NC-transfected MDA-MB-231 cells.
PRMT5 expression affects MDA-MB-231 cell cycle and apoptosis in vitro
We further examined the role of PRMT5 in cell cycle progression and apoptosis in MDA-MB-231 cells following PRMT5, shPRMT5, or NC transfection. Fluorescence-activated cell sorting (FACS) analysis showed that PRMT5 knockdown suppressed the cell cycle progression by decreasing the numbers of cells in S phase and increasing the numbers of cells in G1 phases (Figure 6(a)). Cell cycle progression was promoted in MDA-MB-231-PRMT5 cells via increasing the number of cells in S phase and decreasing the numbers in G2/M phase (Figure 6(c)). Moreover, PRMT5 downregulation could increase the apoptosis rate of MDA-MB-231 without drug treatment (Figure 6(b)). Western blot analysis revealed that PRMT5 knockdown can decrease Bcl-2 expression, and promote caspase 3 and PARP expression in breast cancer MDA-MB-231 cells; moreover, p-ERK expression in shPRMT5-transfected cells was significantly inhibited compared with NC cells without changing the total ERK protein level (Figure 6(e) and (f)). In addition, the expression of p-ERK in MDA-MB-231-PRMT5 cells was higher than NC cells (Figure 5(a)).

PRMT5 expression can affect MDA-MB-231 cell cycle progression and promote cell apoptosis. MDA-MB-231 cells transfected with PRMT5, shPRMT5, or NC are harvested 72 h post-transfection. Flow cytometry analysis of (a and c) cell cycle and (b and d) apoptosis indicated that PRMT5 expression can affect cell cycle progression and PRMT5 knockdown can induce cell apoptosis. Western blot was performed to evaluate protein level of (e) Bcl-2/caspase 3/PARP and (f) p-ERK.
Discussion
Breast cancer is one of the malignant tumors with a high incidence among females all over the world. Although the etiology, pathogenesis, and epidemiology of breast cancer have been widely studied and significant improvements in surveillance and clinical treatment strategies have been made, barriers in the early diagnosis and treatment of breast cancer still exist. The development of breast cancer is connected with multiple molecular mechanisms. Various molecular mechanisms are involved in the development and progression of breast tumors. Recently, more evidence indicates the functional significance of the expression of PRMT5 in the hallmarks of cancers. However, its explicit role in breast cancer still remains unclear. In this study, we examined the PRMT5 expression profile in breast tumors and its correlations with prognosis.
Several studies have confirmed that PRMT5 was highly expressed in cell lines and tissues of breast cancer.18–21 Expectedly, we found PRMT5 expression was upregulated in 25 of 31 fresh primary breast tumor tissues compared with normal tissues. Immunohistochemistry analysis revealed that the PRMT5 expression level was significantly higher in primary breast cancer compared with normal breast tissues (p < 0.01). Importantly, although we did not demonstrate that PRMT5 was significantly higher in metastatic breast tumors compared with primary breast tumors (p > 0.05), there was a tendency that PRMT5 protein expression level was higher in metastatic breast cancer tissues; 25 of 34 (73.5%) metastatic breast tumors expressed high PRMT5 levels compared with that of 19 of 35 (54.3%) primary breast tumor tissues. In 2015, Yang et al. 18 reported that PRMT5 nuclear expression was significantly associated with HER-2 expression in breast cancer tissues (p < 0.05). 18 However, we did not find this phenomenon; our data showed that the relationship between high PRMT5 and estrogen receptor (ER), progesterone receptor (PR), or HER-2 status was not significant (Supplementary Table 1). This discrepancy may have been due to the lower number of breast tissue samples we used (31 vs 114) or the demographic differences within geographical and socioeconomic regions among the Chinese population. 3 Furthermore, Kaplan-Meier analysis revealed that the 462 patients with high PRMT5 levels had significantly shorter RFS than 1198 patients with low PRMT5 expression (HR = 1.73 (1.47–2.05); log-rank p = 3.7e−11). PRMT5 expression was inversely correlated with OS of HER-2 positive breast cancer patients (HR = 0.39 (0.16–1); log-rank p = 0.043), and TNBC patients with higher PRMT5 levels showed decreased OS time compared with those with lower PRMT5 levels (HR = 2.53 (1.15–5.56); log-rank p = 0.017). At present, we evaluated the relationship between PRMR5 and Ki-67 in breast cancer, indicating the function of PRMT5 in promoting cell proliferation. These observations support the hypothesis that PRMT5 may function as an oncogene in breast tumors and also suggest that PRMT5 may play an important role in the tumorigenesis of breast cancer.
By changing PRMT5 expression with PRMT5 transfection or shRNA interference, we revealed that PRMT5 expression could effectively influence the MDA-MB-231 cell growth rate, which means MDA-MB-231 cell proliferation was markedly promoted after PRMT5 transfection, whereas MDA-MB-231-shPRMT5 cell growth was inhibited. ERK is known to play a significant role in cell proliferation, and several growth factors and proto-oncogenes facilitate tumor growth by stimulating the ERK/mitogen-activated protein kinase (MAPK) cascade.27–30 Our data showed dramatic decreases in ERK phosphorylation in the PRMT5-knockdown MDA-MB-231 cells compared with the control cells, but there was no difference in total ERK protein expression. Thus, we proposed that PRMT5 exerted crucial roles not only in breast tumorigenesis but also in tumor progression, and the impaired MDA-MB-231 cell growth in vitro due to the knockdown of PRMT5 could be explained at least in part by the inhibition of ERK phosphorylation. However, the explicit relation between PRMT5 and ERK activity in breast cancer needs to be studied further.
Our research found that the downregulation of PRMT5 in MDA-MB-231 cells can decrease Bcl-2 expression and promote the expression of caspase 3 and PARP without any drug treatment. Bcl-2, caspase 3, and PARP are key proteins in the intrinsic mitochondrial apoptotic pathways; 31 this suggested that the inhibition of PRMT5 may strengthen apoptosis through regulating the apoptotic proteins related to the mitochondrial pathway. FACS analysis revealed that the knockdown of PRMT5 in MDA-MB-231 cells could enhance the apoptotic cell percentage, again suggesting that PRMT5 participates in apoptosis through the Bcl-2/caspase 3/PARP pathway. In addition, we found that PRMT5 overexpression and knockdown can affect cell cycle progression by enhancing and delaying S progression, respectively. To the best of our knowledge, our study is the first to show the association between PRMT5 and cell cycle progression and cell apoptosis in human breast cancer, but an in vivo study needs to be performed in the future to further study the oncogenic role of PRMT5 in breast cancer.
Taken together, we demonstrated the high expression of PRMT5 in primary and metastatic breast cancer lesions and indicated its function in cell proliferation, cell cycle progression, and apoptosis. Moreover, the association of PRMT5 expression with poor prognosis was verified in HER-2 positive breast cancer and TNBC patients, guaranteeing the promising application of PRMT5 in developing effective targeted biomarkers for breast cancer patients.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
This work was funded by research grants from the Social Development Research Project (No. 2014K11-01-02-01) in Shaanxi province.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.