
Abstract
Environmental extremes such as high temperature, cold, alkalinity, drought, and rocky substrate modulate plant growth and promulgate adaptive characteristics in plants. The same plants can be found inhabited in different life forms, namely epiphytic, lithophytic, terrestrial, halophytic, and psammophytic, based upon adaptive characteristics essential for their survival. Microorganisms residing on the surface or within rocks make a unique habitat referred to as “lithobiontic habitat,” which further facilitates weathering phenomena and accumulates water and nutrients from the ecosystem to form a lithobiontic ecological niche. Lithobionts are divided into two groups, namely epiliths and endoliths, based on their presence on rock surfaces. Lithophytes, the rock-inhabiting plants, thriving under extreme stress conditions adapted successfully and their extraordinarily challenging atmosphere encouraged them to make new forms of life. Many biotic and abiotic factors are responsible for dwelling in such environments. Lithophytes interact with their surroundings to obtain vital elements necessary for their metabolism and, because of natural selection, their morphology and physiology help them with better adaptation. Another factor that facilitates such adaptation in lithophytes is the association of a fungus with lithophytic roots to ensure nutrient supply for their survival and facilitate growth in extreme conditions. Arbuscular mycorrhizal (AM) root fungal associations commonly found among plants under stressed conditions help hosts adapt and promote plant life. In lithophytes, AM association aids in nutrient uptake from soil and facilitates plant survival under harsh conditions. The present review provides a comprehensive account of lithophytic life forms and covers different aspects of rock-dwelling plants concerning their historical background, classification, habitat, distribution in various families, root adaptive mechanisms, and phytopharmacological status. This review focused on plant species that can be considered among the most common representatives of lithophytes. This manuscript furnishes insights into different perspectives on the ecological and biological properties of the most common lithophytic species adapted to extreme rocky structures and explains their wide array of applications for pharmacological use.
Introduction
Plant Adaptations
All living organisms depend upon their surroundings and feed on them to grow and establish themselves. They live with the help of air, water, soil, and different climatic conditions. It provides all the desired components or nutrients that are required for their morphological and physiological establishment. Any feature of an organism that enables it to exist under the conditions of its habitat is known as adaptation. This special feature allows an organism to survive in adverse conditions. Some principles affect plant distribution that is managed by different climatic conditions, soil types, and biotic factors. In the case of plants, the whole community, or population, adapts accordingly, which adds survival value to their lives through a chain of adaptations. 1 Eugenius Warming (1895) explained these adaptations and was influenced by limiting factors for vegetation in ecology and classified plants into different groups based on their requirement for water and also based on the substratum on which they grow into different categories, namely oxylophytes (plants of acidic soil), psammophytes (plants inhabited on sand), halophytes (plants flourish on saline soil), lithophytes (plants growing on the surface of rocks), and chasmophytes (plants inhabited in crevices of rocks). 2 Natural selection leads to adaptations that help to emerge distinctive phenotypic traits among floral species, and they are identified by being variable and heritable. They fit according to Darwin’s “Survival of the Fittest” theory. During evolution, there have been various phenotypic changes across diverse environments and among taxa. These changes are often reflected in different adaptation series. 3 Many biotic and abiotic factors are responsible for adaptations in plants and other living organisms. There are many conditions prevailing, like water deficit, to cope with variations in the basic metabolic rate of higher plants. 4 There are two ways through which plants correspond to environmental stress, namely adaptation and acclimation. An “adaptation” is considered a heritable modification in structure or functions that mold an organism to survive or reproduce under stress conditions, whereas “acclimation” confers to a nonheritable modification in response to switching environments. These modifications are caused by the structural and physiological plasticity of the plants. 5 Also, when our ancestors shifted from wild plants to actively cultivated plants, they imposed intensive selective pressures on traits, resulting in phenotypic variations. These traits include not only conscious selection, like increased yield, but also unconscious selection, for example, reduced dormancy. 6 Evolutionary adaptations lead to advancements in genetics and become symbols of natural genetic variation, too. Field studies are required for better recognition of the concept of emerging adaption sequences. Plants furnish excellent opportunities to explore the basis of evolving sequences of different environmental conditions. 7 Stoichiometric flexibility is the inherent ability of organisms to adjust their requirement for essential elemental balance concerning environmental changes. 8 This flexibility plays a critical role in maintaining elemental balance in a particular plant species concerning changes in the environment. Changes in substrate availability directly influence nutrient assimilation in plants, thereby causing fluctuations in stoichiometry.9–11 Wu et al., 12 reported that because substrate shift, there exists a remarkable stoichiometric and isotopic flexibility between epiphytes and lithophytes. With changes in the substrate, the difference in foliar stoichiometry is encountered in two different ecotypes of facultative epiphytes. Lithophytic species were found to be enriched in δ15N and Ca and depleted in Fe, K, and Mn when compared with epiphytic ones. 12
Methodology
We conducted a literature review and data compilation of lithophytes and tried to inculcate all published literature available till date. We performed an exhaustive search from December 01, 2022 to December 10, 2023 in scientific databases, including Web of Science, Sci-Hub, Sci-Finder, Scopus, Sciencedirect PubMed, and Google Scholar with the following keywords and combinations, “lithophytes,” “lithobionts,” “chasmophytes,” “petrophytes,” “pharmacology of lithophytes,” “plant adaptations in lithophytes,” and “rock plants.” In this article, we have incorporated information from some rare books that date back to the seventeenth to twentieth centuries, for which Google books (
Lithophytes
Historical Perspectives
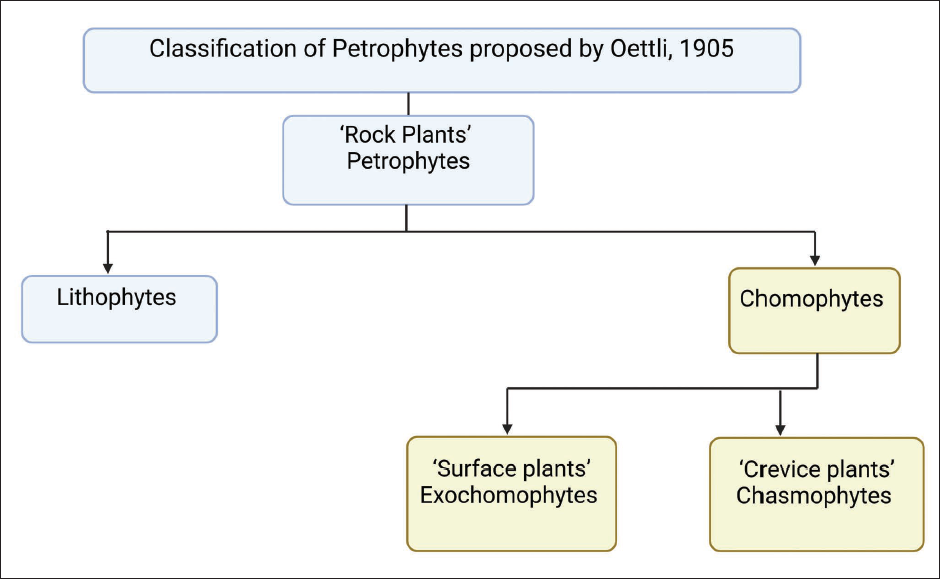
The word “lithophyte” finds its roots in the earlier dictionaries of the seventeenth and eighteenth centuries. 13 In The New and Complete Dictionary of the German and English Languages, composed chiefly after the German dictionaries of Mr. Adelung and Mr. Schwan, “lithophytes” are described as natural bodies that have nature-like stones or corals. 14 In some writings, lithophytes are described as stony plants 15 that nourish from rocks and stones, not from the Earth. 16 The term “lithophyte” was revived in the mid-nineteenth century in famous writings. A History of British Sponges and Lithophytes 17 was authored by George Johnston (1797–1855), a Scottish physician and naturalist who was a fellow of the Royal College of Surgeons, Edinburgh. He mentioned British sponges and lithophytes and arranged lithophytes into two families, namely Corallinade (calciferous algae) and Nulliporide (amorphous solid calcareous vegetation). 17 John Quekett (1815–1861), a renowned professor of histology, delivered “Lectures on Histology” at the Royal College of Surgeons of England, in the session 1851–1852, Vol –II. In Structure of the Skeleton of Plants and Invertebrate Animals, he mentioned the term lithophytes (stone plants) in context to Corallines (red algae associated with order Corallines) and Nullipore (lime-secreting coralline algae) coated with calcareous material occurred in abundance on coral reefs. In lecture 10, “Skeleton of Lithophytes,” he remarked that lithophytes were previously considered animals, but later investigations revealed them as plants coated with calcareous material. 18 From 1799 to 1804, Alexander von Humboldt (German naturalist and explorer), along with his fellow Aime Bonpland (French explorer and botanist) traveled to Latin America. They published their scientific results of the expedition in 1853 in a book entitled Personal Narrative of Travels to the Equinoctial Regions of America, During the Years 1799-1804, Volume 3, 19 in which they also incorporated the term “lithophyte polypi” about limestone corals while mentioning breccia (sedimentary rock) of Cumana (currently, a city in Venezuela). 19 John Edward Gray (1800–1875), a British zoologist, in Catalogue of Lithophytes or Stony Corals, published in 1870, which is in the collection of the British Museum, mentioned lithophytes as “stony corals” under the class Polypes or Coralliaria belonging to Zoophytaria order and Rupicole as suborder, 20 and it was continued to be used in the context of “corals.” 21 In 1898, the visualization of plants as lithophytes was observed by German Botanist, Andreas Franz Wilhelm Schimper (1856–1901) in his famous writings Pflanzen-Geographie auf Physiologischer Grundlage, that is, Plant-Geography upon a Physiological Basis in 1898 (translated by W.R. Fisher in 1903). 22 He encompasses both lithophytes and chasmophytes. Schimper used the term lithophytes for the vegetation found on the rock/stone surfaces, whereas copious vegetation accumulated on crevices of rocks, like chasmophytes. Lithophytes, as the name suggests, comprise two words: “litho” meaning rock and “phytes” meaning plants. 23 They are regarded as petrophytes or rock plants. Max Oettli (1879–1965), a Swiss Educator, studied in 1905 the ecology related to rock vegetation in Switzerland and defined pteridophytes/rock plants as plants that are populated or colonized permanently on the rocks or parted pieces of rocks as enduring species. He classified pteridophytes (rock plants) on the basis of growth on rocky substratum into lithophytes and chomophytes (Figure 1). Chomophytes are further subdivided into exochomophytes (surface plants) and chasmochomophytes/chasmophytes (crevice plants). 24 Danish Botanist Eugenius Warming (1841–1924) critically examined plant communities relationship with the environment and opened new doors for research in plant ecology. He gave insights into the adaptive behavior of lithophytes and chasmophytes. In 1909, Warming also incorporated the term lithophytes in his book Oecology of Plants: An Introduction to the Study of Plant Communities 25 with special reference to Schimper and Oettli. He also described different forms of lithophytes that are arranged uniquely and can colonize in steep, inclined, and bare rocks, and he mentioned chasmophytes as rooted plants in rock clefts filled with detritus. Lithophytes grow on the surface of rocks and usually get their nutrients from available water sources such as rain, melting snow, running water, and dew. Their ability to survive extreme climatic conditions may change their morphology or physiology for better adaptation. There is a special feature in lithophytes known as haptera, that is, an outgrowth of the stem that aids plants in fixing themselves on the base of rocks. They are essentially required by lithophytes to grow in such utmost circumstances. 25 Lithophytes growing solely on rocks are obligate ones, whereas both rocks and soil are facultative. 26 The terms “saxicolous” and “rupicolous” are also used concurrently in the context of plants growing in cracks predominantly spread in rocky habitats.27, 28 Some examples of saxicolous lichens include Parmelia saxatilis. Parmelia glabratula ssp. fuliginosa is abundantly found on slate rocks, 29 and Saxifrago syntaxa are the rupicolous plant communities inhabited on limestones. 30
Classification of Petrophytes.
Lithobiontic Ecological Niche
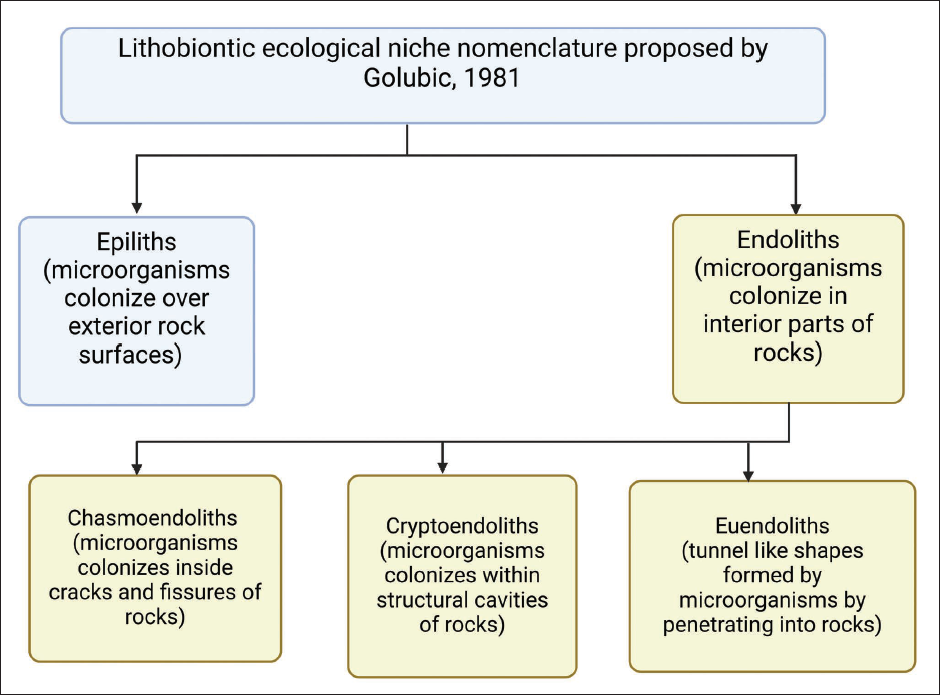
The concurrent existence of plants and microbes leads to the building of a symbiotic relationship between the two, which helps them survive and aids in their growth and development. They interact with each other via various classical and novel inter-kingdom signaling mechanisms. Plants can sense and respond to microorganisms inhabiting their environments. 31 Plant growth-promoting bacteria isolated from the rock-inhabiting the giant cactus species Pachycereus pringlei facilitate the growth of the plant when compared to the control. 32 Dark septate endophytes (DSEs) also interact with plants in stressful conditions.33, 34 Plants residing in harsh environments interact with microorganisms for their survival. The lithobiontic term was used to represent organisms that inhabit the surface and inside hard rock substrates. It is confirmed that microorganisms dwell on various rocks, whether it is marine, freshwater, or terrestrial, and terminology is proposed according to that, and it is relevant to all rock-inhabiting organisms irrespective of their environment. Lithobionts are divided into two categories: epiliths (living on the rock surface) and endoliths (living within the rock). Organisms that dwell inside the rocks have a special tendency to adapt and differentiate themselves into particular microbial communities, and the endolithic ecologic niche is subdivided into chasmoendoliths (dwell in fissures and cracks of rocks), cryptoendoliths (hidden endoliths, that colonize into the cavities of porous rocks), and euendoliths (true endoliths, penetrates actively into the interiors of rocks and forms tunnel shapes) (Figure 2). 35 Thin accumulations of microorganisms found either in epilithic or crypto endolithic situations are sometimes referred to as “biofilms.” These biofilms involved in the weathering of rocks are further classified by Viles depending upon their thickness into different categories, namely biofilms (accumulations of less than 1-mm thick microorganisms), biorinds (microbial association with rocks 1–5 mm), and biocrust (association with greater than 5-mm organisms). 36 Colonization of microbes, especially in association with rocks, makes an unusual and differentiating feature that refers to the “lithobiontic environment.” Biotic components of nature exclusively combine with microorganisms to produce a new type of community that is recognized for their exceptional distinctive prevalence on the rock’s surfaces, crevices, and underneath. Such a consortium of rocks and microbes is generally seen in extreme environmental conditions such as the desert, marine, and arctic regions where temperature, salinity, and humidity vary. These places, because of their extraordinarily challenging atmosphere, promote a new form of life that is resistant to extreme stress conditions.37–39 These ecological habitats resulting from the collaboration of microbes with the rock or rock boulders are known as the “lithobiontic ecological niche.” 37 Lithobionts play a significant role in an impoverished environment. They are known as primary producers, and they own genetic as well as functional nitrogen fixation, representing a direct contribution to growing plant communities requiring carbon and nitrogen. They follow primary metabolic pathways and have immense genetic plasticity; therefore, lithobionts are expository and involved in harsh ecosystem survival.40, 41 An association of microbes with rocks is accountable for the colonization of plants, which further promotes the formation of soil because of the weathering process and also accumulates water and nutrients from the ecosystem. 42 Jennie Fry 43 conducted experiments on lithophytic algae and successfully explained the mechanical action of these lichens on rocks which facilitates weathering phenomena in rocks. These lichens penetrate rhizoids into tiny fissures of rocks to establish themselves and are often termed rhizolithophytes, whereas exolithophytes are algae that only attach to the rock surface. 44 Weathering of rocks by microbes is crucial for the availability of nutrients required for the growth of plants on rocks. The microbes provide essential minerals, for example, P, K, Mn, Fe, Cu, and Zn, and make them tolerant in extreme conditions of temperature and drought-like situations. 32 Algae, considered both aquatic and nonaquatic organisms and survive in extremely arid and desert-like conditions, seem to improve and enhance their ecological constraints through the moisture, soil, and nutrients available on the rocks. They need mineral substrates to feed upon, but owing to the lack of soil, they accomplish their nitrogen needs from the atmosphere, and their capability of nitrogen-fixing in such an environment makes them an exceptional form of life. 45 Vascular plants have also been reported to exist in a close relationship with the microbial communities of rocks and their substrates. Some examples include Pachycereus pringlei, Stenocereus thurberi, Mammillaria fraileana, Opuntia cholla, Ficus palmeri,32, 42, 46 Pachycormus discolor, 47 Festuca rubra subsp. pruinosa, 48 and Bouteloua dactyloides. 49 Considering the plant’s unique ability to adapt to different life forms, Lopez and Bacilio, 50 recently proposed a conceptual model of rock weathering involving microbial association with succulent plants such as agave and cacti in arid landforms. This hypothetical model explains how organic acid exudations from succulents interact with lithobiontic microorganisms to enhance nutrient cycling and also aid in soil formation, 50 and such an adapted form of life could be for a new life outside our planet too. 51
Lithobiontic Ecological Niche Nomenclature Proposed by Stjepko Golubic, 1981.
Root Fungal Associations
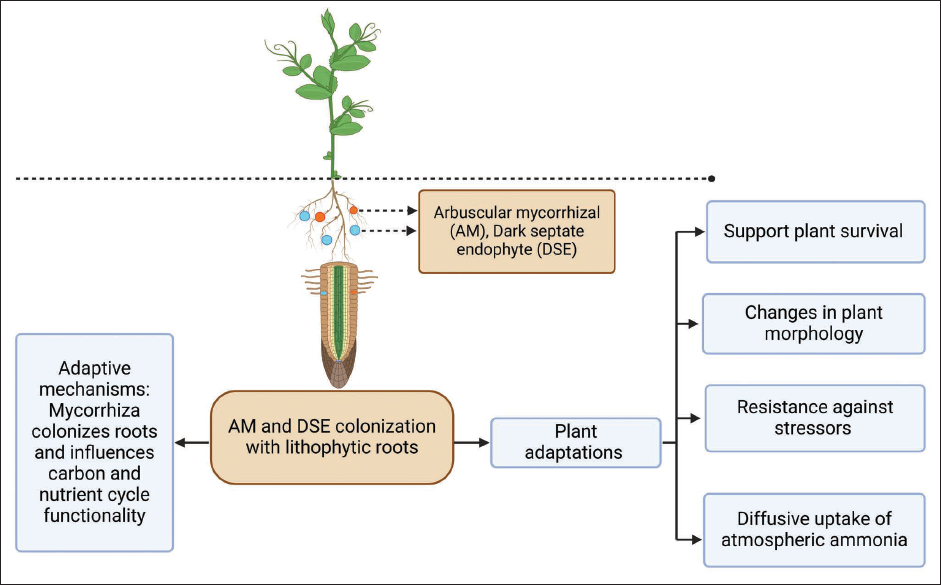
In 1885, an association between plant roots and mycorrhizal fungi was propounded by German Botanist, Albert Bernhard Frank. 52 Based on the structure and function of mycorrhiza, four distinct types of association were observed in plants, namely arbuscular mycorrhizal (AM), orchid mycorrhiza, ectomycorrhiza, and ericoid mycorrhiza. 53 AM colonization is frequently found in plants surviving in different habitats and exists in a symbiotic relationship with the host. Plants can control this fungal symbiont functioning via a feedback mechanism depending upon the requirements of nutrients needed by the plant. 54 Mycorrhiza colonizes roots and influences carbon and nutrient cycling functionality. 55 This relationship often helps the plant to adapt and survive in harsh conditions, namely high temperature, salinity, and drought. 56 In addition, more stressful conditions in plants may lead to diverse fungal associations, particularly in epiphytes and lithophytes. 57 Several examples include an increased volume of roots of Hordeum vulgare in colonization with Glomus intraradices when exposed to drought. 58 Improvement in root and shoot biomass along with the content of copper, phosphorous, and zinc was observed when Acacia nilotica in colonization with Glomus fasciculate was exposed to excessive salinity conditions. 59 Studies have also been conducted to understand mycorrhizal relationships between epiphytic and lithophytic life forms. 60 Yokoya et al. 61 hypothesized that mature lithophytes/epiphytes having well-developed root systems are less associated with fungus than with juvenile ones. 61 Coelogyne corymbosa, an endangered epiphyte/lithophyte, is associated with mycorrhizal fungal communities such as Cantharellales and Sebacinales. 62 The AM association is also reported in lithophytic mosses 63 and ferns.64, 65 Muthukumar et al. 66 observed this relationship in some nonorchidaceous lithophytes and found it widespread in association with AM and DSEs in saxicolous plants (lithophytes). Coelogyne viscose exists in both epiphytic and lithophytic life forms. Rhizoctonia fungal communities associated with this plant vary significantly. Lithophytic life forms are predominantly associated with the Tulasnellaceae in comparison to Sebacinales with epiphytic life forms. 67 Jahan 68 studied this association in Porpax reticulate, an epiphytic or lithophytic orchid in which mycorrhizal fungus resides inside plant roots in a symbiotic relationship with a host. Dearnaley et al. 69 mentioned different techniques to study mycorrhizal orchid relationships such as molecular barcoding, stable and radioactive isotopes, gene expression studies, and genome sequencing. Lithophyte root fungal associations illustrated in Figure 3 indicated several adaptive characteristics.
Lithophyte Root Fungal Associations and Adaptive Characteristics.
Distribution of Lithophytes in Different Plant Families
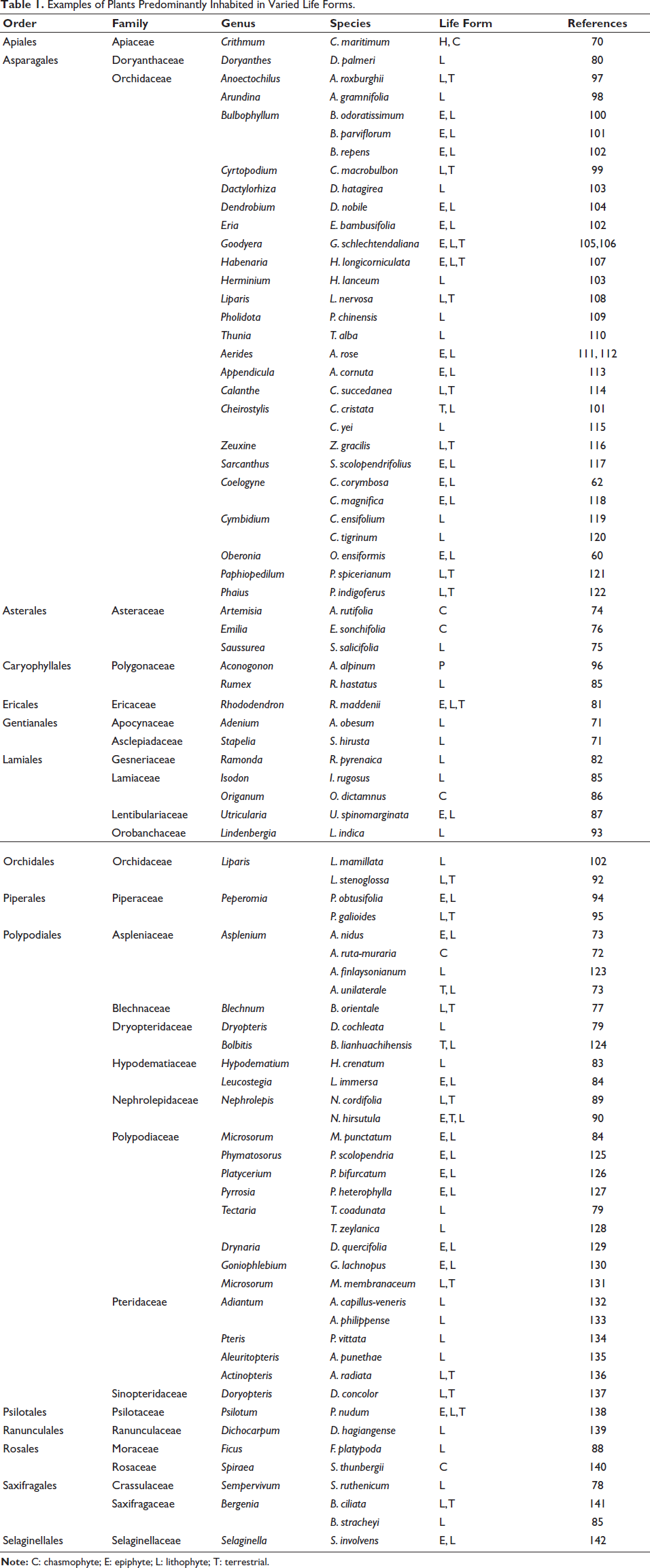
Plants presented in this article are inhabited in different life forms: lithophytic, chasmophytic, terrestrial, and epiphytic. They are predominantly distributed in various families, as represented in Table 1, namely Apiaceae, 70 Apocynaceae, Asclepiadaceae, 71 Aspleniaceae,72, 73 Asteraceae,74–76 Blechnaceae, 77 Crassulaceae, 78 Dryopteridaceae, 79 Doryanthaceae, 80 Ericaceae, 81 Gesneriaceae, 82 Hypodematiaceae,83, 84 Lamiaceae,85, 86 Lentibulariaceae, 87 Moraceae, 88 Nephrolepidaceae,89, 90 Orchidaceae,91, 92 Orobanchaceae, 93 Piperaceae,94, 95 Polygonaceae,85, 96 and Orchidaceae.97–99
Examples of Plants Predominantly Inhabited in Varied Life Forms.
Phytopharmacological Potential of Plants Inhabited in Multiple Life Forms
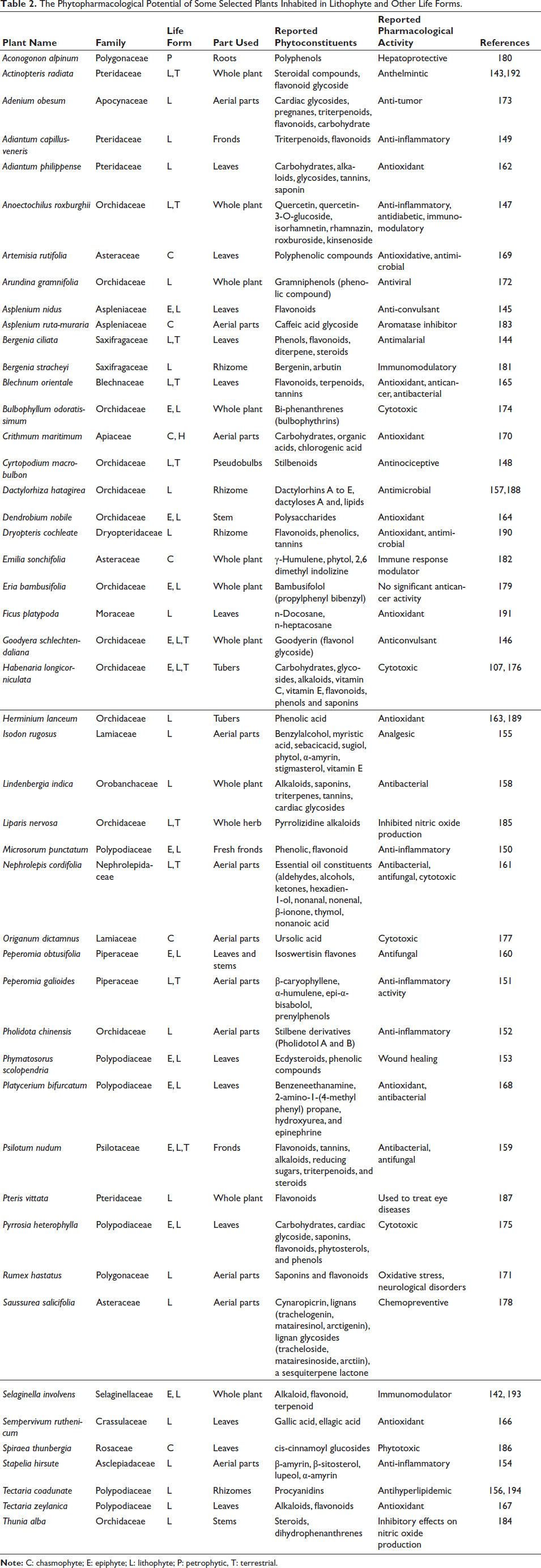
Here, we mention published pharmacological activities of plants that exist in different life forms, including lithophytic species, and we have tried to review every aspect, highlighting the therapeutic potential of a given species as represented in Table 2. It includes anthelmintic activity in Actinopteris radiata 143 ; antimalarial activity in Bergenia ciliata 144 ; anticonvulsant activity in Asplenium nidus 145 and Goodyera schlechtendaliana 146 ; anti-inflammatory activity in Anoectochilus roxburghii, 147 Cyrtopodium macrobulbon, 148 Adiantum capillus-veneris, 149 Microsorum punctatum, 150 Peperomia galioides, 151 Pholidota chinensis, 152 Phymatosorus scolopendria, 153 Stapelia hirsuta, 154 and Isodon rugosus; 155 antihyperlipidemic activity in Tectaria coadunata; 156 antimicrobial activity in Dactylorhiza hatagirea 157 and Lindenbergia indica; 158 antifungal activity in Psilotum nudum, 159 Peperomia obtusifolia, 160 and Nephrolepis cordifolia 161 ; antioxidant activity in Adiantum philippense, 162 Herminium lanceum, 163 Dendrobium nobile, 164 Blechnum orientale, 165 Dryopteris cochleata,Ficus platypoda,Sempervivum ruthenicum, 166 Tectaria zeylanica, 167 Platycerium bifurcatum, 168 Artemisia rutifolia, 169 Crithmum maritimum, 170 and Rumex hastatus; 171 antiviral activity in Arundina gramnifolia; 172 cytotoxic activity in Adenium obesum, 173 Bulbophyllum odoratissimum, 174 Pyrrosia heterophylla, 175 Habenaria longicorniculata, 176 Origanum dictamnus, 177 Saussurea salicifolia, 178 and Eria bambusifolia; 179 hepatoprotective activity in Aconogonon alpinum; 180 and immunomodulatory potential in Bergenia stracheyi, 181 Selaginella involvens, 142 and Emilia sonchifolia. 182 Other activities include aromatase inhibitory activity in Asplenium ruta-muraria; 183 inhibited nitric oxide (NO) production by Thunia alba (T. alba) 184 and Liparis nervosa; 185 and phytotoxic effects in Spiraea thunbergii 186 and Pteris vittata (P. vittata) in eye disorders. 187
The Phytopharmacological Potential of Some Selected Plants Inhabited in Lithophyte and Other Life Forms.
Anthelmintic
Aqueous and ethanolic extracts obtained from the whole plant of Actinopteris radiata were evaluated in vitro in Pheretima posthuma, an earthworm, for anthelmintic activity. The ethanolic extract showed efficacy at 200 mg/mL when compared with the standard drug Albendazole. 143 The plant is inhabited in Savannah as a lithophytic fern. 136
Antimalarial
Bergenia ciliata, commonly found in moist rocky crevices/slopes, showed promising antiplasmodial activity. 141 Ethanolic extracts of leaves showed activity against Plasmodium berghei in a dose-dependent manner with IC50 < 10 µg/mL and in vivo suppressive activity in Peter’s 4-day test. This study also supports the traditional claim that this plant is an antipyretic. 144
Anticonvulsant
The methanolic extract obtained from leaves of Asplenium nidus was evaluated for its anticonvulsant effect in pentylenetetrazole and isoniazid-induced convulsions in Swiss mice. The crude extract significantly delayed the duration and onset of convulsions. 145 The plant commonly exists as an epiphytic, lithophytic fern. 73 Goodyerin (25 and 50 mg/kg), a flavonoid glycoside isolated from Goodyera schlechtendaliana, significantly reduced locomotor activity and prolonged pentobarbital-induced sleeping time in mice. 146 Plants are populated in all habitats, such as terrestrial, lithophytic, or epiphytic.105, 106
Anti-inflammatory Activity
Epiphytic and lithophytic fern Adiantum capillus-veneris is found on humus-rich rocks in shady areas. 132 Alcoholic extract, hexane fraction, and triterpenes isolated from hexane fraction exhibited anti-inflammatory activity in formalin and croton oil-induced inflammation in mice. 149 Aqueous and organic extracts obtained from pseudobulbs of Cyrtopodium macrobulbon (C. macrobulbon) were evaluated for antinociceptive activity in acetic acid-induced writhing and hot plate assay. Aqueous extract showed 35% inhibitory activity at 100 and 316 mg/kg, respectively, and organic extract exhibited 30% inhibitory activity at 100 mg/kg. This study also supported the ethnopharmacological evidence of C. macrobulbon used in traditional Mexican medicine for urinary ailments. 148 This plant is generally inhabited in rocky areas as a terrestrial or lithophyte. 99
Silver nanoparticles synthesized from fronds of fern Microsorum punctatum showed in vitro anti-inflammatory potential in protein denaturation bioassay. These nanoparticles effectively suppressed thermally induced albumin denaturation. 150 The plants are usually found as low epiphytes or lithophytes. 84 Peperomia galioides (P. galioides), a lithophytic or terrestrial herb, 95 when evaluated against cotton pellet-induced granuloma, croton oil-induced ear edema, myeloperoxidase, and Escherichia coli (E. coli) lipopolysaccharide-induced inflammation showed anti-inflammatory potential. The data provided evidence that P. galioides has the potential to inhibit cellular inflammatory mediators such as NO and tumor necrosis factor. 151 Stilbene derivatives (Pholidotol A and B) isolated from ethyl acetate fraction obtained via partitioning ethanolic extract from aerial parts of Pholidota chinensis exhibited significant NO production in RAW 264.7 (macrophage cell line) with IC50 value at 24.3 and 17.1 mM. 152 This lithophytic orchid is frequently used by Indian tribes in scrofula, hemorrhage, dysentery, etc. 109 Stapelia hirsuta, a succulent lithophyte, 71 showed anti-inflammatory potential when evaluated in carrageenan-induced rat paw edema using Indomethacin as a standard drug. Several compounds have been isolated from the n-hexane fraction, namely lupeol, β-sitosterol, α-amyrin, and β-amyrin. 154 Isodon rugosus, a lithophyte predominantly populated in mountain gorges, 85 showed analgesic effects in different pharmacological models, namely acetic acid-induced writhing test (53% analgesia), formalin (61% analgesia) at a dose of 100 mg/kg, and hot plate assay (average reaction time of 7 min). GC-MS detected major eight compounds responsible for the analgesic action of plants, namely sebacic acid, myristic acid, phytol, tocopherol, sugiol, stigmasterol, benzyl alcohol, and α-amyrin. This study also justifies ethnomedicinal uses of this plant as a potential analgesic medicine. 155
Anoectochilus roxburghii, a terrestrial and lithophytic herb, 97 showed anti-inflammatory, antidiabetic, and immunomodulatory activities in different experimental models. 147
Antihyperlipidemic
Lithophytic fern, Tectaria coadunata, 79 showed antihyperlipidemic activity in fat diet-induced hyperlipidemia in rats. Powdered rhizome, 540 mg/kg significantly reduces triglycerides, very-low-density lipoprotein (VLDL), and low-density lipoprotein (LDL). The dose of rhizome powder was found to be safe even at 2000 mg/kg, as no toxic effects were observed at this dose. 156
Antimicrobial
Aerial parts of lithophyte, Dactylorhiza hatagirea, 103 showed resistance against E. coli with MIC > 125 mg/mL.157, 188
Lindenbergia indica showed antibacterial activity when evaluated against several bacterial strains with minimum inhibitory concentrations (MICs) ranging from 31.25 to 125 mg/mL. 158 This plant is commonly found in rock crevices. 93 Phymatosorus scolopendria, a scrambling, creeping epiphyte/lithophytes, 125 showed in vitro antibacterial activity against Staphylococcus aureus with MIC ≥ 4 µg/mL. 153 Psilotum nudum showed in vitro antibacterial and antifungal activities. Chloroform, ethanol, and water extracts of this plant showed significant antifungal activity against Microsporum gypseum,Trichophyton mentagrophytes, and Trichophyton rubrum. 159 P. nudum is a pteridophyte generally inhabited as an epiphyte, terrestrial, and lithophyte. 138 Isoswertisin flavones isolated from leaves and stem portions of Peperomia obtusifolia exhibited weak antifungal activity against Cladosporium cladosporioides and Cladosporium sphaerospermum when evaluated in the bioautographic assay. This plant is typically an epiphyte and lithophyte. 94 A terrestrial and lithophytic plant, Nephrolepis cordifolia 89 exhibited antibacterial and antifungal effects in vitro. Essential oils from aerial parts tested against Klebsiella pneumonia showed an MIC of 3.9 mg/L. When screened for antifungal activity, oil showed significant activity against Microsporum gypseum and Trichophyton rubrum with MICs of 0.98 and 3.9 mg/mL respectively. 161
Antioxidant Activity
Methanolic extract prepared from the leaves of Adiantum philippense showed in vitro antioxidant activity having IC50 of 140.00 ± 0.86 µg/mL. The extract also showed notable cytotoxic and thrombolytic activities. 162 This plant exists in a lithophytic life form inhabited near shaded walls in humid places. 133 Tubers of Herminium lanceum are sources of phenolic acid. 189 Ethanolic extract of tubers, when evaluated in the 2,2-diphenyl-1-picrylhydrazyl (DPPH) assay, showed significant antioxidant activity in a dose-dependent manner. 163 This plant is predominantly found as lithophytes in the Bhaba Valley of Himachal Pradesh, India. 103 A water-soluble heteropolysaccharide (DNP) isolated via column chromatography from stems of Dendrobium nobile exhibited antioxidant activity in a 2,2-azino-bis-3-ethylbenzothiazoline-6-sulphonic acid (ABTS) assay with 78% scavenging activity. 164 Dendrobium is an epiphytic and lithophytic orchid native to the Himalayas, China, and Indochina. 104
Polyphenols present in different fractions of leaves of Blechnum orientale exhibited strong antioxidant potential in the DPPH assay. 165 The plant is populated by exposed rocks. 77 Lithophytic fern, a Dryopteris cochleata 79 acetone extract, is reported to exhibit antioxidant activity in DPPH, ABTS, lipid peroxidation assays, etc. The results found that rhizomes exhibited free-radical scavenging capacity. 190 A lithophytic Ficus species Ficus platypoda found in an arid region reported 88 exhibiting antioxidant potential in the DPPH assay. The ethanolic extract prepared from leaves showed scavenging activity at different concentrations with an EC50 value of 232.6 µg/mL. 191 Sempervivum ruthenicum, a petrophytic plant that usually grows in stony places, 78 contains polyphenolic compounds (gallic acid and ellagic acid) present in the ethanolic extract and showed antioxidant activity in the DPPH assay with the EC value 4.6112 ± 0.08 mg/mL. 166 The endangered lithophytic fern Tectaria zeylanica 128 showed free-radical scavenging potential when its ethanolic extract obtained from leaves was evaluated in a DPPH assay with an IC50 value of 0.78 mg/mL. 167 Platycerium bifurcatum is found predominantly in different life forms, namely epiphytic and lithophytic. 126 The chloroform fraction obtained via leaves at 400 µg/mL concentration showed optimum antioxidant activity in the DPPH assay. 168 Artemisia rutifolia, a drought-adapted petrophytic plant found exclusively in chasmophytic vegetation, 74 when evaluated against DPPH, showed antioxidant activity with an IC50 of 39 µg/mL. The plant yielded several polyphenolic compounds, namely caffeic acid, chlorogenic acid, gallic acid, sinapic acid, syringic acid, m-coumaric acid, p-coumaric acid, vanillic acid, ferulic acid, quercetin, and myricetin. 169
Aerial parts of Crithmum maritimum showed in vitro antioxidant activity in the DPPH and ABTS assays. Meot-Duros and Magné 170 compared the free-radical scavenging activity of C. maritimum distributed in contrasting habitats (sandhills and cliffs). 170 The plant is commonly known as sea fennel and is inhabited as halophyte and chasmophyte. 70 Rumex hastatus, a lithophyte, 85 showed strong in vitro antioxidant activities. Crude saponins and flavonoids showed scavenging activity with IC50 312 and 104 µg/mL, respectively, in the DPPH assay in a dose-dependent manner. 171
Antiviral
Gramniphenols (phenolic compounds) isolated from the whole plant of Arundina gramnifolia showed anti-tobacco mosaic virus activity (U1 strain) along with anti-HIV activity in the cellosaurus cell line (C8166). 172 Arundina gramnifolia is a rare lithophyte found in the Meghalaya region of India. 98
Cytotoxic Activity
Adenium obesum, often known as a desert rose inhabited as a lithophyte, 71 showed cytotoxic activity when evaluated against human epidermoid nasopharynx carcinoma, and hepatocellular carcinoma, cervix cancer, breast cancer. 173 Bulbophythrins A and B (dimeric phenanthrenes) isolated from the whole plant of Bulbophyllum odoratissimum (B. odoratissimum) exhibited cytotoxicity against HL-60 (human leukemia cell line) and BEL-7402 (hepatoma cell line). 174 B. odoratissimum typically resides in epiphytic and lithophytic habitats. 100 The ethyl acetate extract from leaves of Pyrrosia heterophylla showed cytotoxicity in A375 cells with an IC50 of 250 µg/mL. 175 This plant exists as an epiphytic/lithophytic fern. 127 Extracts prepared from tubers of Habenaria longicorniculata exhibited cytotoxic activity when evaluated against MCF 7, HepG2, and HeLa cell lines at different concentrations, 1000 and 500 µg/mL, with CTC50 value of >1000 on all tested cell lines. 176 The plant inhabits all terrestrial, epiphytic, and lithophytic habitats. 107 Aerial parts of perennial chasmophyte, Origanum dictamnus, 86 when evaluated against murine leukemia (P388) and human bronchial epidermoid cancer cell lines (NSCLC-N6) showed cytotoxicity. The dichloromethane extract (ID50 of 8, 14 µg/mL) and ursolic acid (ID50 of 3.5, 9 µg/mL) isolated from it manifested cytotoxicity against cell lines. 177 Saussurea salicifolia, a lithophyte forb, 75 exhibited chemoprotective action against AGS cells and Hepa 1c1c7 cells. 178 Eria bambusifolia is an epiphytic/lithophytic orchid. 91 A new (propyl phenyl) bibenzyl isolated from this plant was tested against different cell lines, HL-60, SMMC-7721, A-549, MCF-7, and SW-480, but no significant activity was observed. 179
Hepatoprotective Activity
Polar fractions obtained from dried roots of Aconogonon alpinum showed a reduction in elevated levels of biochemical parameters (aspartate aminotransferase, alanine aminotransferase, alkaline phosphatize, total bilirubin) attributed to the liver in a dose-dependent manner against carbon tetrachloride-induced liver injury in mice because of the presence of polyphenolic compounds. 180 This petrophytic plant is frequently found in coniferous forests of the Ural Mountains. 96
Immunomodulatory
Bergenin and norbergenin isolated from Bergenia stracheyi showed anti-arthritic potential via modulating Th1/Th2 cytokine balance in mice. 181 The plants are commonly inhabited in rocky vegetation. 85 The water extract (100 mg/kg) of Selaginella involvens stimulated thymus growth in mice in a dose-dependent manner. The plant is usually inhabited in the Western Ghats of Kerala as an epiphyte and lithophyte. 14 Emilia sonchifolia, a chasmophyte, 76 stimulates an immune response in experiment models. 182
Other Biological Activities
Asplenium ruta-muraria, a chasmophyte, 72 showed aromatase inhibitory activity in vitro. 183 Extracts obtained from a lithophytic fern, P. vittata, 134 showed the potential to treat eye disorders. Wahid et al. 187 studied the biological effects of extracts of P. vittata on visual processes involving retina neuronal cells in experimental models. 187 The extract showed a 96% safety profile even at a low 500 µg/mL concentration. Cis-cinnamoyl glucosides obtained from Spiraea thunbergii exhibited phytotoxic effects. 186 The plant is commonly found as a chasmophyte. 140 T. alba, a lithophytic orchid, 110 exhibited inhibitory effects on NO production. A series of stibenoids isolated from stem portions of T. alba via column chromatography was evaluated for NO inhibitory activity, which showed promising results with IC50 values ranging between 3.20 and 20.50 µM. 184 Liparis nervosa, a terrestrial or lithophytic orchid, 108 inhibited NO production in vitro. Pyrrolizidine alkaloids isolated from plants inhibited LPS-induced NO production in RAW264.7 macrophages with IC50 values ranging between 2.16 and 38.25 µM. 185
Summary and Future Research
Plants can successfully adapt to contrasting environmental conditions for their survival. Some plants are found to flourish in distinct life forms such as terrestrial, epiphytic, and lithophytic. Each life form has its unique adaptive characteristics, which may differ in morphology and physiology and may also undergo stoichiometric elemental changes. Vegetation on rocks has attracted the interest of scholars/researchers in the past decades, and they favorably classify and establish rock plants (petrophytes) as lithophytes and chomophytes. Microorganisms residing inside and on rock surfaces create an exceptional environment referred to as the “lithobiontic environment” and are also reported to interact with algae, mosses, and other vascular plants, facilitating them to thrive under harsh conditions. Lithophytes represent a multifaceted group of plants that can be grown in stressful conditions with minimal nutrient requirements. They are adaptable in such a way that they combat different environmental stressors by interacting with their surroundings. Lithophytic life forms also establish symbiotic relationships with fungal communities, as AM resides inside roots. This relationship further enhances plant adaptability by diffusive uptake of atmospheric ammonia, changing plant morphology, protection against stress, etc. There has been no comprehensive review published on lithophytes till date. This review article aims to rejuvenate the concept of lithophytes and focus on its origin, ecological niche, root fungal associations, and therapeutic potential. Lithophytes are widely distributed among different orders and more likely into Asparagales and Polypodiales. The phytopharmacological profile of plants was selected by their existence in different life forms, inculcating lithophytes exhibiting diverse biological activities. Based upon phytopharmacological data, it may be concluded that the same plant exists in different life forms, namely terrestrial, epiphytic, and lithophytic, exhibiting a wide array of pharmacological activities containing complex phytoconstituents such as tannins, flavonoids, glycosides, carbohydrates, and essential oils. We felt that there is a need to exploit lithophytic life forms with a different view, considering their ability to tolerate abiotic stress by interacting with their surroundings. Furthermore, lithophytes also serve as precious reservoirs to monitor and furnish novel stress responsiveness, and the data incorporated in this article will enlighten our perception of lithophytes.
Footnotes
Abbreviations
Acknowledgments
The authors express their heartfelt gratitude to CT University, Ludhiana, Punjab; Dr. Vinay Thakur, Principal, Govt. Pharmacy College Rakkar H.P.; and Dr. Vivek Sharma, Professor, Govt. College of Pharmacy Rohru, for providing the support necessary to complete this work.
Declaration of Conflicting Interests
The authors declare no potential conflicts of interest concerning this article’s research, authorship, or publication.
Ethical Approval
Not applicable to this study.
Funding
The authors received no financial support for the research, authorship, and/or publication of this article.
Informed Consent
Not applicable.