
Abstract
Background
Cadmium, a toxic heavy metal, poses a significant risk to human health. Its potential to contribute to brain tumorigenesis is a growing concern. Brain tumours are notoriously difficult to treat due to their aggressive nature, poor prognosis and resistance to conventional therapies.
Purpose
To investigate the impact of cadmium exposure on brain tumour development, focusing on its influence on the Sonic Hedgehog (SHH)–glioma-associated oncogene 1 (GLI1) signalling pathway and the stem cell marker B cell-specific Moloney murine leukaemia virus integration site 1 (BMI1).
Methods
A total of 25 Wistar rats of 200–250 g body weight range were classified into five groups: Test groups one and two were orally administered with cadmium chloride in drinking water (30 mg/L, 60 mg/L). Positive controls one and two were administered orally with zinc in drinking water (30 mg/L and 60 mg/L) and the negative control group rats received double-distilled water continuously for 28 days. We calculated the rat’s water intake each week. From the third to the fourth week, we observed behavioural changes using an open field test (OFT). The animals were sacrificed on day 29. Thereafter, the brain and hindbrain were separated for further assessment, including real-time reverse transcription polymerase chain reaction (RT-PCR) to measure gene expression levels of key genes in the SHH–GLI1 signalling pathway and stem cell markers.
Results
Water intake decreased in both the test and the positive control groups. The OFT analysis revealed significant behavioural changes in all parameters of both the test and positive control groups compared to the negative control group, including movement patterns, speed, travel distance, activity levels and freezing behaviour. Gene expression indicates increased SHH, GLI1, BMI1 and tumour protein 53 (TP53) expression in both the test and positive control groups.
Conclusion
Cadmium exposure may initiate and promote brain tumour development by regulating SHH, GLI1, BMI1 and TP53. This may disrupt the normal regulatory mechanisms of the SHH–GLI1 pathway.
Keywords
Introduction
Cadmium is a heavy metal commonly found in the environment due to industrial processes, mining and the use of cadmium-containing products, such as batteries and pigments. It is a potent toxin that poses significant risks to human health, particularly when exposure occurs over prolonged periods. 1 In addition to its well-documented adverse effects on renal, pulmonary and cardiovascular systems, cadmium is recognised as a carcinogen by the International Agency for Research on Cancer. Among its various cancer-inducing mechanisms, cadmium’s potential to promote brain tumorigenesis has sparked growing concern. 2
Brain tumours remain among the most challenging forms of cancer due to their aggressive nature, poor prognosis and resistance to current therapies. Gliomas, including glioblastoma multiforme, are the most common type of malignant brain tumour and are notoriously difficult to treat.3–7 Understanding the molecular pathways underlying brain tumour development is crucial for identifying new therapeutic targets. Recent research suggests that cadmium may influence the Sonic Hedgehog (SHH)–glioma-associated oncogene 1 (GLI1) cell signalling pathway, a key pathway involved in neural development and tumour growth and the stem cell marker B cell-specific Moloney murine leukaemia virus integration site 1 (BMI1), 8 which regulates the self-renewal and proliferation of cancer stem cells. This study aims to explore the role of cadmium in brain tumour development by investigating its effects on these molecular targets.9–11
Cadmium exposure occurs through inhalation, ingestion or dermal contact, primarily from industrial emissions, contaminated food and tobacco smoke. Once absorbed into the body, cadmium is stored in various tissues, including the liver and kidneys, where it can remain for decades due to its long biological half-life. Cadmium’s toxicity is largely attributed to its ability to generate reactive oxygen species, leading to oxidative stress, deoxyribonucleic acid (DNA) damage and inflammation. 12 These processes contribute to the initiation and progression of cancer by inducing mutations, altering gene expression and disrupting cellular signalling.
Brain tumours can arise from various cell types within the brain. These tumours develop when genetic mutations disrupt normal cell growth and communication. One crucial pathway involved in this process is the SHH–GLI1 cell signalling pathway. Normally, this pathway helps to regulate cell development and organisation but when it malfunctions, it can contribute to the growth of brain tumours. However, aberrant activation of the SHH–GLI1 pathway has been associated with the development of gliomas and other brain tumours. Similarly, BMI1, a protein that regulates stem cell function, has been implicated in maintaining cancer stem cell populations within tumours, contributing to their growth and resistance to therapy. 13
The SHH–GLI1 cell signalling pathway plays a crucial role in embryonic development, particularly in the central nervous system, including the brain. SHH, a secreted protein, binds to its receptor, Patched1, thereby releasing the inhibition of the Smoothened (SMO) receptor. This activation of SMO ultimately leads to the upregulation of GLI transcription factors, which control the expression of genes essential for cell growth, differentiation and survival.14–17 In the context of cancer, dysregulation of this pathway can result in uncontrolled cell growth and tumour formation. Gliomas, one of the most aggressive forms of brain cancer, often exhibit aberrant SHH signalling. There is growing evidence that cadmium may interfere with this pathway, potentially promoting tumour development and progression.4–7
BMI1, a member of the Polycomb group proteins, is crucial for the self-renewal of stem cells, including those in the neural stem cell lineage. It regulates chromatin structure and gene expression. In cancer, BMI1 overexpression in various tumour types, such as brain tumours, fuels the survival and proliferation of cancer stem cells. These cells, known for their resistance to conventional treatments, are believed to drive tumour initiation, progression and recurrence. The upregulation of BMI1 in brain tumours is associated with poor prognosis and increased tumour aggressiveness. Studies suggest that cadmium may alter BMI1 expression, thereby contributing to the maintenance of cancer stem cell populations in brain tumours. 18
Several studies have indicated a potential link between cadmium exposure and increased cancer risk, particularly in the context of lung, prostate and breast cancers. However, its role in brain tumours is less well understood. Research has shown that cadmium can activate oncogenic signalling pathways, including the SHH–GLI1 pathway, by promoting the expression of GLI transcription factors. This, in turn, may lead to the proliferation of tumour cells and the formation of brain tumours. Additionally, cadmium has been shown to increase BMI1 expression in certain cancer cell lines, suggesting that it may contribute to the maintenance of cancer stem cells in brain tumours.19–21
The physiological metal-ion zinc is necessary for zinc-finger transcription factors such as GLI1 to function and maintain their structural integrity.22, 23 This control enables us to determine whether observed alterations are specific to cadmium-induced toxicity and zinc-site disruption, rather than a general response to divalent cation exposure, by comparing the effects of the toxic cadmium ion (Cd²⁺) with those of its essential analogue (Zn²⁺). 24
Zinc-finger proteins (ZFPs) are a class of proteins that contain zinc-finger domains, structural motifs in which zinc ions (Zn²⁺) are coordinated by cysteine and histidine residues. These domains are crucial for functions such as DNA/RNA binding, protein-protein interactions and gene regulation in transcription factors, DNA repair proteins and signalling molecules. Zinc-finger domains in DNA repair proteins, which rely on zinc ions for structure, can be damaged by carcinogenic metals through metal substitution or oxidation. This disruption compromises DNA repair and represents a key molecular mechanism in metal-induced carcinogenesis. 25
A plausible mechanism involves the replacement of zinc (Zn²⁺) by cadmium (Cd²⁺) in the zinc-finger regions of transcriptional regulators, driven by their chemical similarities and preferences for soft ligands. ZFPs have cysteine and histidine residues that bind Zn²⁺ for stability and function. Toxic heavy metals often target these proteins. Cadmium, which is similar to zinc and has a strong affinity for thiolate groups, can easily swap with zinc in these regions. For instance, studies have shown that Cd²⁺ can replace Zn²⁺ in the CCCH ZFP tristetraprolin.24, 26 This swap results in a protein with a modified RNA binding ability. This example supports our observation that cadmium exposure disrupts the zinc-finger transcription factor GLI1. Future research using techniques such as cellular thermal shift assays with GLI1 or mutating its zinc-binding residues, will be crucial to confirm this direct interaction in our model (Figure 8).
Another study demonstrates that oral cadmium exposure induces significant neurotoxicity in the rat cerebellum by disrupting oxidative balance, increasing free radicals and causing indirect DNA damage, leading to structural neurodegeneration. Furthermore, cadmium dysregulates the SHH signalling pathway, triggering its noncanonical upregulation and the increased expression of downstream targets Pax6 and Nkx2.2 as part of a compensatory repair response, while simultaneously impairing cellular proliferation. 27
Experimental studies of animal models have demonstrated that cadmium exposure can enhance the proliferation and survival of brain tumour cells by modulating key signalling pathways. 28 For instance, cadmium has been shown to increase SHH and GLI1 expression in glioblastoma cells, leading to enhanced tumour growth and treatment resistance. Furthermore, cadmium-induced oxidative stress may activate the SHH–GLI1 pathway and upregulate BMI1, thereby promoting tumorigenesis. These findings underscore the potential of cadmium to contribute to brain tumorigenesis and point towards targeting the SHH–GLI1 pathway and BMI1 as promising avenues for therapeutic intervention.29, 30 Tumour protein 53 (TP53) has been the most often documented malfunctioning molecule in tumours since its discovery as an oncogene in 1979. 31 Among all medulloblastoma (MB) brain tumour patients, SHH MBs with TP53 mutations are one of the two most vulnerable subgroups. In contrast, TP53 pathway activation is observed in SHH MB cells.32–37
Methods
Animals
Adult male Wistar rats (N = 25), weighing 200–220 g, were obtained from the Central Animal House Facility, JN Medical College (JNMC), Faculty of Medicine, Aligarh Muslim University (AMU), Aligarh, Uttar Pradesh, India. Rats were housed in five groups under controlled conditions, including a 12-hour light/dark cycle, a temperature of 22 ± 2 °C and access to an ample supply of food and water. After a four-week adjustment period, behavioural assessments were conducted daily between 10:00 am and 2:00 pm. All experiments were conducted in accordance with ethical guidelines and were approved by the Institutional Animal Ethics Committee (IAEC) at JNMC, AMU, Aligarh.
Twenty-five male Wistar rats were randomly divided into five groups of five animals each, based on prior literature on neurotoxicity models, to detect significant differences in pathway markers, such as GLI1 expression (Figure 1).
Negative control group: Received drinking water without the addition of Zn and CdCl2 and the drinking water was free of contaminants, only double-distilled water. Positive control group 1: Received 30 mg/L zinc in their drinking water. Positive control group 2: Received 60 mg/L zinc in their drinking water. Test group 1: Received 30 mg/L CdCl2/L in their drinking water. Test group 2: Received 60 mg/L CdCl2/L in their drinking water.
Methodology of the Study.Source: Adopted from Tayyab et al. 48
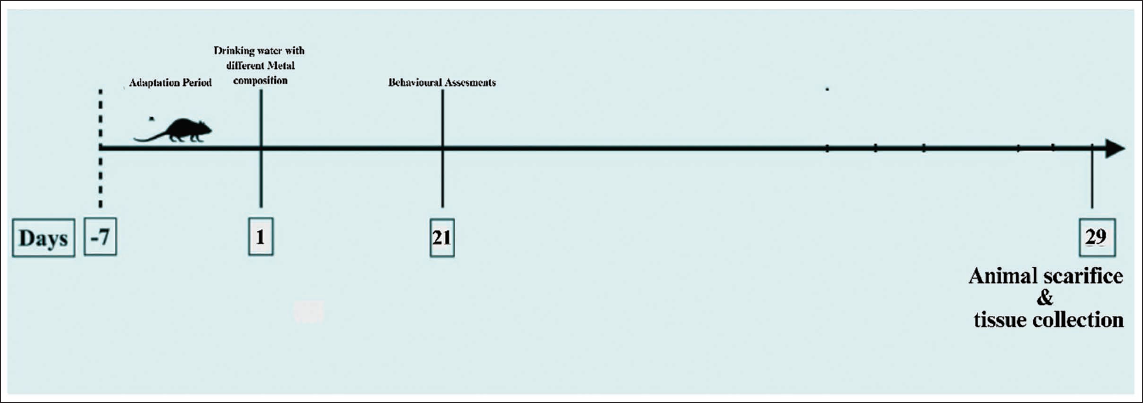
All the rats were observed for their behavioural changes on days 21 and 28. On day 29, all the rats were killed and their brain samples were isolated and preserved for further assessments.
Water Intake and Metal Uptake Assessment
In a 28-day study, rats were monitored daily at 9:00 am to assess their consumption of 500 mL of metal-mixed distilled water provided in sipper bottles. Each day, the sipper bottles were filled with freshly prepared distilled water mixed with metal and the remaining water was measured the following morning to determine the volume consumed by the rats. This daily monitoring enabled accurate tracking of water intake for each rat group.
The concentration of ingested metal in the rats was assessed by measuring metal levels in the water they consumed. Initial metal-mixed distilled water samples were collected to establish baseline metal concentrations. The difference between the initial and remaining water volumes, along with the metal concentrations, was used to calculate the amount of metal uptake for each group. This methodology ensured the collection of precise and consistent data on both water consumption and metal intake throughout the study period.
Open Field Test
The behavioural study was conducted on days 21 and 28 of the experiment using the open field test (OFT). The open field apparatus was a black wooden box measuring 60 cm by 60 cm, with walls that were 30 cm high. The floor was divided into 16 equal squares (15 cm by 15 cm). The centre consisted of four squares. An overhead camera connected to an ANY-maze tracking system (Stoelting, IL, USA) recorded exploratory behaviour. Between experiments, the arena was cleaned with cotton soaked in 70% alcohol. All videos were analysed by the experimenter. For each rat, the following behaviours were recorded: (a) Locomotion: The number of lines crossed by the rat over five minutes intervals, (b) distance, (c) mean speed, (d) time active, (e) time inactive, (f) active episodes, (g) inactive episodes, (h) maximum active episode, (i) line crossings, (j) maximum speed 10, (k) time freezing, (l) freezing episodes, (m) mean freezing score and (n) rotations.
RNA Isolation
RNA was extracted from the hindbrain of Wistar rats using a QuickPrep RNeasy Mini Kit (Qiagen, UK, Cat# 74104). Approximately 10 mg of stored RNAlater tissue samples were homogenised after centrifuging at 5000 rpm at 4 °C. The supernatant was removed and 500 µL PBS was added. For five minutes, centrifuge at 5000 rpm at 4 °C. Mix the cell lysate with 350 µL of RLT buffer and 350 µL of 70% ethanol before passing it to an RNase Mini spin column in a 2 mL collecting tube. The flow-through should be discarded after centrifugation at 8,000 g for 15 seconds at 4 °C. Add 700 µL of RW1 and spin at 8,000 g for 15 seconds at 4 °C, then discard the flow-through. Subsequently, 500 µL of RPE should be added to the RNase microspin column. The flow-through should be discarded after centrifugation at 8,000 g for 15 seconds at 4 °C. To the RNase micro spin column, add 500 µL of RPE. Centrifuge at 8,000 g for two minutes at 4 °C, then discard the flow-through. To dry the membrane, place the RNeasy column in a fresh tube and centrifuge at high speed for one minute. Place the RNeasy column in a 1.5 mL tube and fill it with 30 µL of RNase-free water. For 15 seconds, centrifuge at 8,000 g at 4 °C. Store the RNA at −80 °C for future complementary DNA (cDNA) synthesis.
cDNA Synthesis
A total of 2 µg of RNA was used to synthesise cDNA. The RNA (2 µg) was combined with 1 µL of the random hexamer and the final concentration was adjusted to 12 µL using nuclease-free water. After thoroughly mixing the tubes, they were incubated for five minutes at 85 °C. On the ice, the tubes were combined with 2 µL of first-strand synthesis buffer, 4 µL of 2.5 mM dNTP mix and 1 µL of Mu-MLV reverse transcriptase. The tubes were then incubated for 60 minutes at 42 °C, 10 minutes at 92 °C and five minutes at 4 °C. The cDNA was created and kept at −20 °C.
Real-time RT-PCR
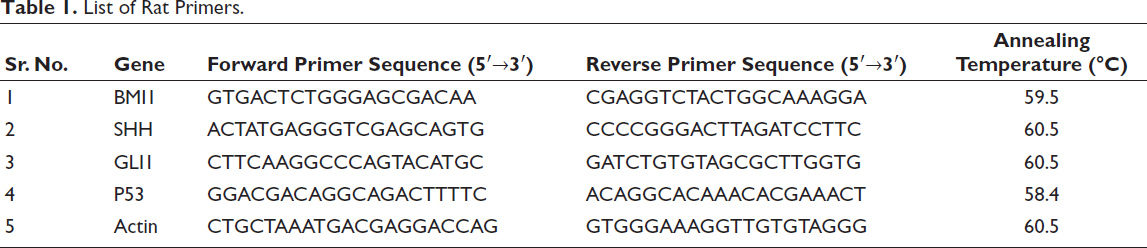
A total of 2 µL of 50 ng cDNA, 10 µL of Light Cycler 480 SYBR Green I Master (Roche Diagnostic, Germany), 1 µL each of forward and reverse primers (Table 1) and 6 µL of water were added to a well of a polymerase chain reaction (PCR) plate (Roche Diagnostics, Germany) and amplified on a Real-time PCR Machine, LightCycler 96 (Roche Diagnostic, Germany). Light Cycler@96 software was used to analyse data based on ∇∇Ct.
List of Rat Primers.
Statistical Analysis
We repeated each experiment three times and used the average results and standard deviation to analyse our data. We also performed real-time PCR three times and analysed the results using LightCycler® 480 Software. Statistical significance was set at p < .05. All analyses were performed using the software Prism 7.
Results and Discussion
Decreased Water Intake in Test and Positive Control Groups as a Marker of Cadmium-induced Physiological and Behavioural Changes
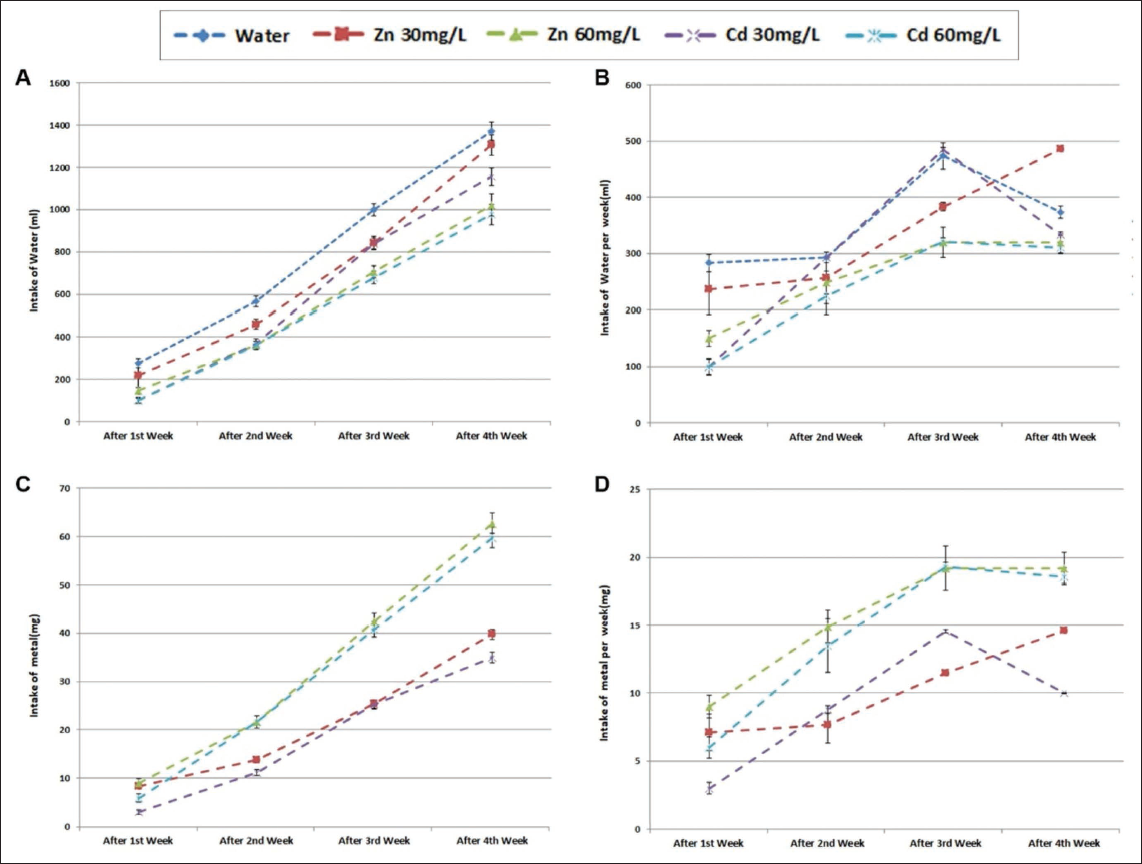
In both the test and positive control groups, a noticeable decrease in water intake was observed, suggesting that cadmium exposure may have induced physiological or behavioural changes that affected normal drinking patterns (Figure 2). This reduction in water intake could be an indicator of underlying stress or discomfort due to cadmium’s toxic effects, as changes in hydration behaviours are often associated with disruptions in metabolic processes, appetite and overall health. Furthermore, such reductions in water consumption may have broader implications for the body’s ability to manage toxins, as proper hydration is essential for maintaining cellular functions and eliminating harmful substances.
(A) Total Intakes of Different Metals in Mixed Drinking Water. (B) Total Intakes of Different Metals in Mixed Drinking Water Per Week. Total Intakes of Metal D. Total Weekly Intake of Zinc (30 or 60 mg/L) or Cadmium Chloride (30 or 60 mg/L) During the Four-week Administration Period (p < .05).
Behavioural Alterations in Cadmium-exposed Subjects Observed Through Open Field Test Analysis
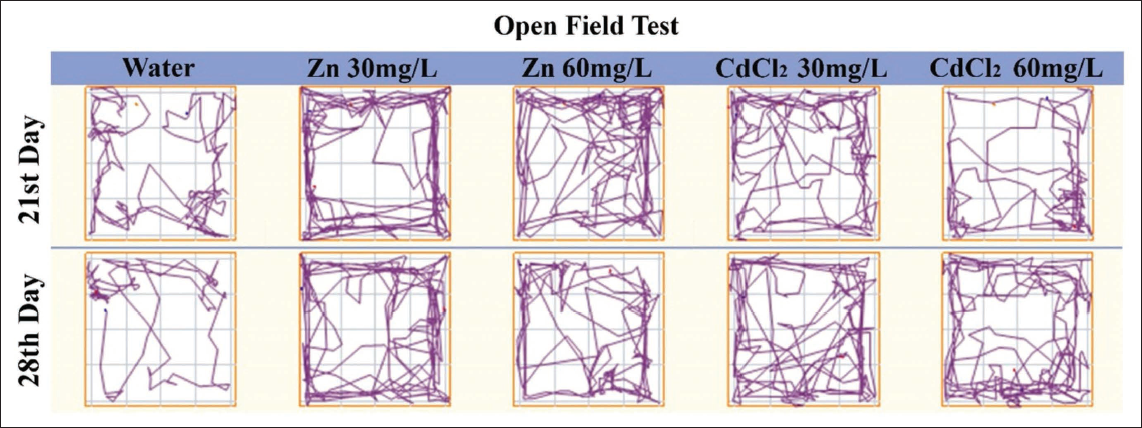
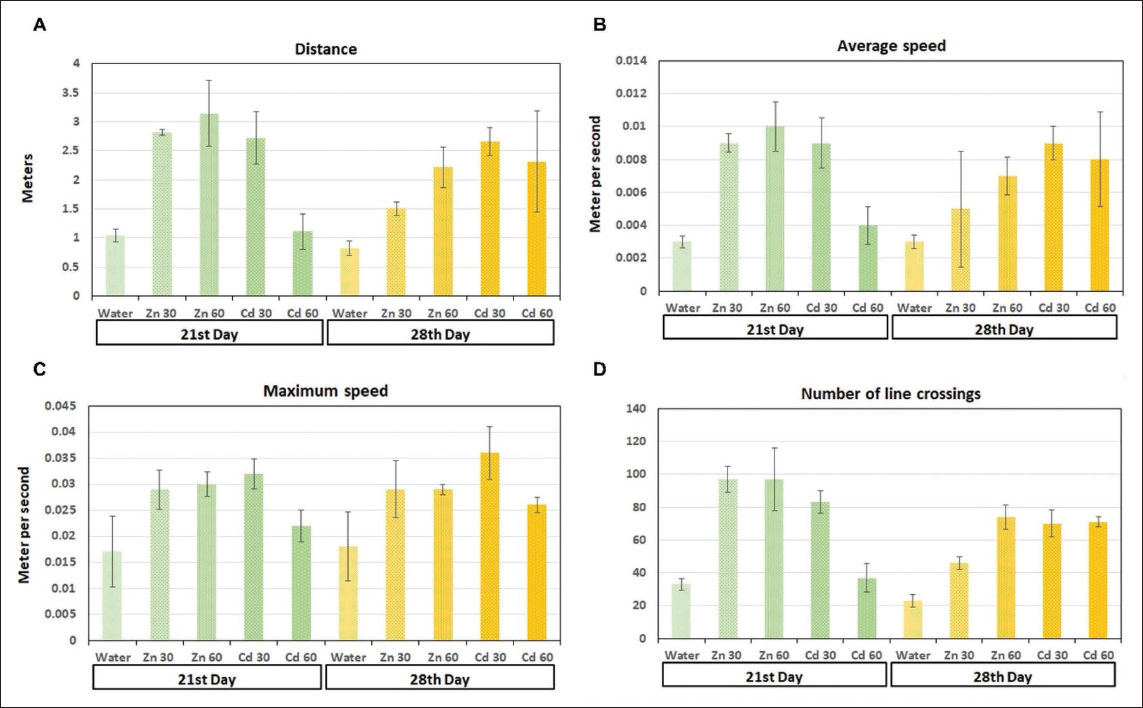
The OFT analysis provided additional insights into the behavioural impacts of cadmium exposure. In both the test and positive control groups, significant alterations in various behavioural parameters were observed compared with the negative control group (Figure 3). These changes included alterations in movement patterns, where cadmium-exposed subjects displayed irregular or reduced movement, potentially reflecting impaired motor function or heightened anxiety. Speed and travel distance were also significantly affected, with both groups exhibiting reduced overall locomotor activity, which could be interpreted as a result of neurotoxic effects on motor coordination and muscle control (Figures 4–6).38, 39 Additionally, activity levels in cadmium-exposed subjects were markedly lower, indicating a potential suppression of normal exploratory behaviour and motivation. The increase in freezing behaviour further suggests heightened anxiety or fear responses, which are common outcomes in animals experiencing neurotoxic stress.40–42
ANY-maze Images of the Open Field Test. After Days 21 and 28, the Administration of Zinc (Zn) at Concentrations of 30 or 60 Milligrams Per Litre (mg/L) and Cadmium Chloride (CdCl₂) at Concentrations of 30 or 60 mg/L was Compared to Distilled Water.
(A) Total Distance Travelled: The Total Distance the Animal Covered During the Test. (B) Average Speed: The Average Speed of the Animal During the Test, Calculated by Dividing the Total Distance Travelled by the Test Duration. (C) Maximum Speed: The Highest Speed Reached by the Animal. The Speed Between Positions is Calculated and the Maximum Value is Identified. (D) Area Changes: The Number of Times the Animal’s Centre Moved from One Area of the Apparatus to Another. The Apparatus is Divided into Different Zones. For Each Recorded Animal Position, the Zone Containing the Animal’s Centre is Determined. Every Time the Zone Changes, the Measure’s Value Increases by One (p < .05).
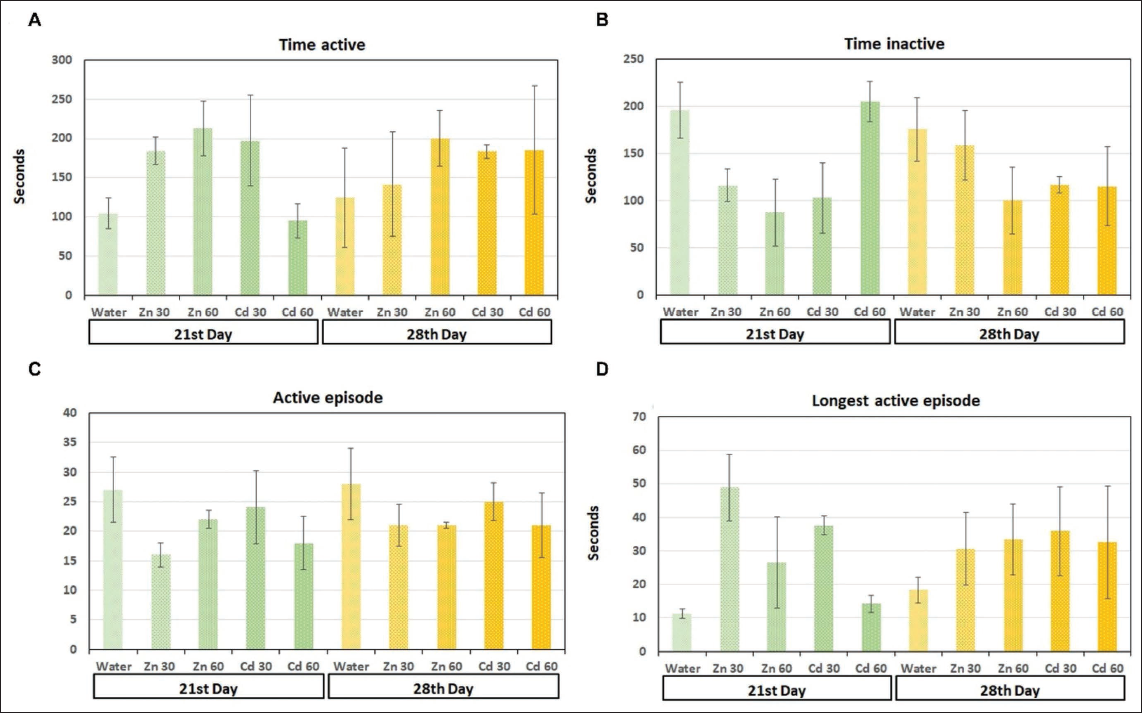
(A) Total Active Time: The Time the Animal was Active, Calculated by Subtracting Inactive Time from the Test Duration. (B) Total Inactive Time: The Combined Duration of all Inactive Periods. (C) Number of Active Episodes: The Count of Times the Animal Transitioned from Inactive to Active. (D) Longest Active Episode: The Duration of the Longest Continuous Period of Activity (p < .05).
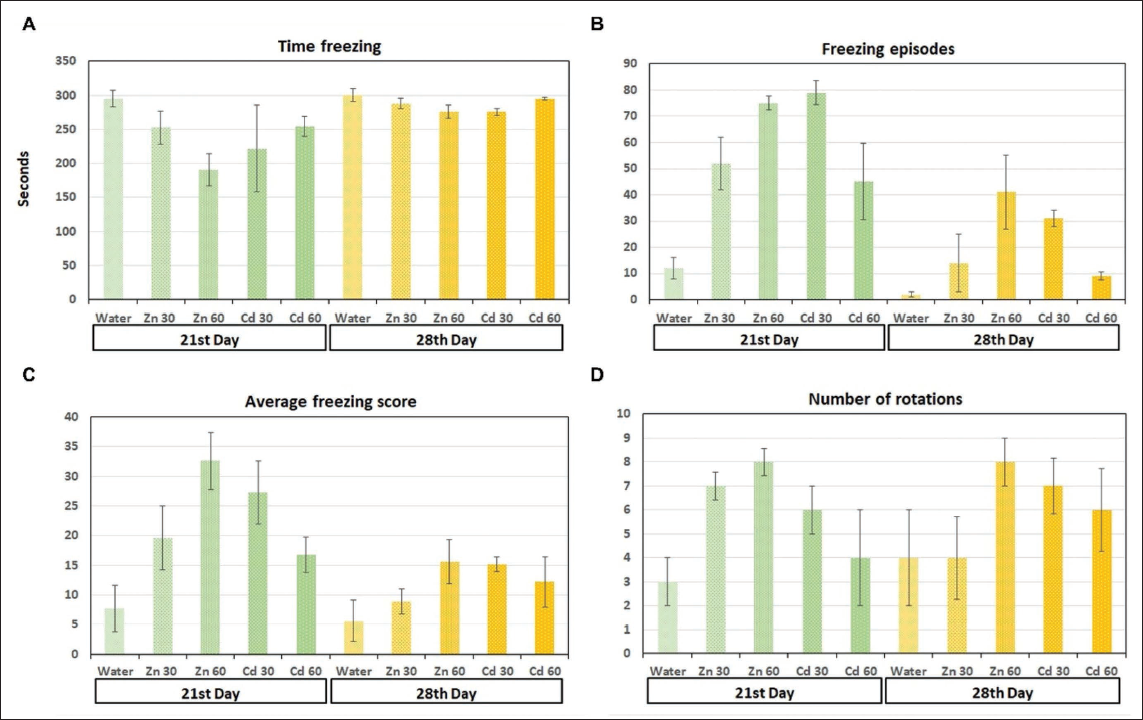
(A) Total Freezing Time: The Total Duration of all Freezing Episodes is Calculated. (B) Freezing Frequency: The Number of Times the Animal Froze During the Test is Counted. (C) Average Freezing Score: The Average Freezing Score is Calculated by Dividing the Total Freezing Score by the Number of Freezing Episodes. (D) Number of Rotations: The Number of Complete Rotations of the Encoder is Counted. A Rotation is Counted When a Continuous Sequence of Pulses in the Same Direction is Received (p < .05).
Molecular Changes in Gene Expression Induced by Cadmium Exposure
On the molecular level, gene expression analysis revealed critical changes in the test and positive control groups (Figure 7). Specifically, there was a notable upregulation of genes involved in key signalling pathways related to tumorigenesis and cell regulation. The expression of SHH and GLI1, both components of the SHH–GLI1 pathway, was significantly elevated, indicating that potential dysregulation of this pathway is essential for cell growth and development. Abnormal activation of this pathway is often associated with various cancers, including brain tumours. The increased expression of BMI1, a stem cell marker involved in maintaining the self-renewal and proliferative capacity of cancer stem cells, suggests that cadmium may promote the growth of tumour-initiating cells, contributing to tumour progression and resistance to therapy.9–11, 43
Relative mRNA Expression of Target Genes of Rats After 28 Days of AD Libitum Access to Drinking Water Containing Cadmium Compared to Control Animals Receiving Distilled Water. (A) SHH, (B) GLI1, (C) BMI1, (D) TP53 (p < .0001).
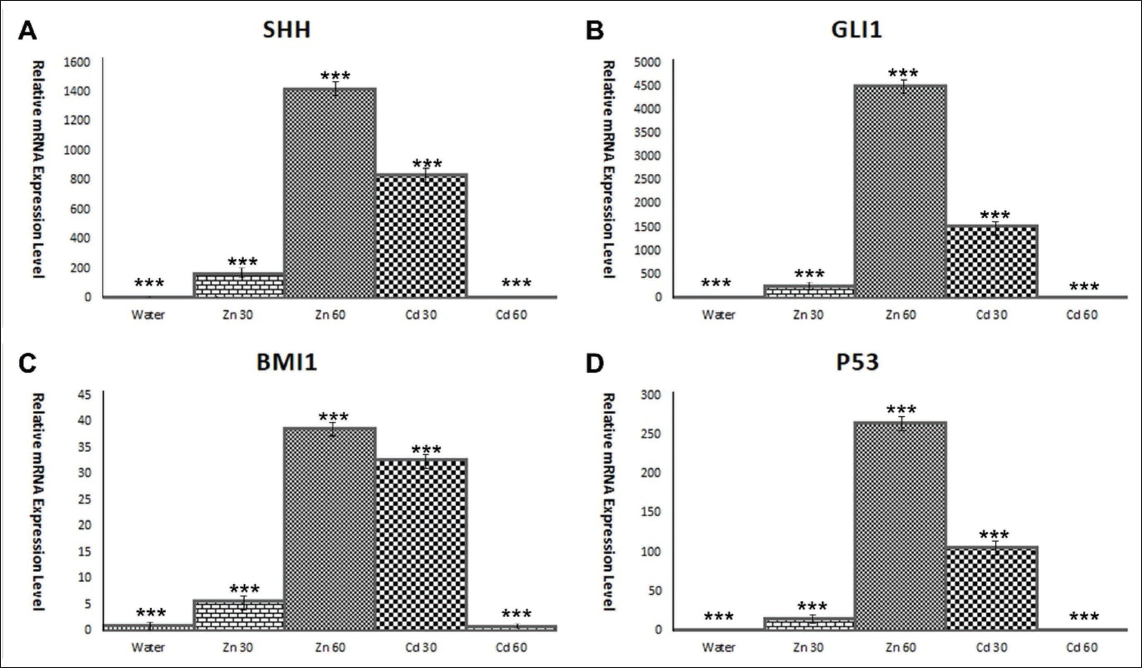
Moreover, the upregulation of TP53, a malfunctioning molecule in tumours since its discovery as an oncogene in 1979, 31 in both the test and positive control groups, could reflect the cellular response to DNA damage and stress induced by cadmium exposure. The TP53 protein is crucial for regulating the cell cycle and preventing the proliferation of damaged cells; its increased expression may indicate that cadmium has triggered cellular mechanisms to mitigate the harmful effects of oxidative stress and genomic instability. However, in the context of chronic cadmium exposure, the protective role of TP53 may become compromised, potentially leading to uncontrolled cell growth and the initiation of tumorigenesis.32–37 Overall, these findings highlight the profound impact of cadmium on both behavioural and molecular levels, with significant implications for its role in promoting cancer-related pathways.
While the proposed mechanism provides a molecular hypothesis, establishing a direct causal link between environmental cadmium and brain tumours in humans requires consideration of broader epidemiological evidence. Establishing a direct link between an environmental toxicant, such as cadmium and a specific cancer, such as glioma, requires strong longitudinal epidemiological data. This is difficult to obtain due to long latency periods and other influencing factors. Our laboratory findings do not substitute such evidence but address the important idea of biological plausibility within accepted causal frameworks. Cadmium is a known neurotoxicant and carcinogen in humans, mostly linked to lung cancer, but also possibly to other conditions. 44 Importantly, research indicates that glioma patients exhibit higher levels of heavy metals, including cadmium, which may promote tumour growth through oxidative stress and alterations in gene expression.9, 11 For instance, studies have reported elevated cadmium levels in serum, cerebrospinal fluid and tumour tissues of patients with malignant brain tumours, including gliomas, compared to controls. 45 By identifying a clear cancer-causing pathway (SHH–GLI1-BMI1) activated by cadmium in brain cells, our work offers a testable mechanism explaining how cadmium exposure could lead to changes in glial cells. This finding provides a mechanistic foundation that supports and explains observed toxicological associations, making a broader causal hypothesis more credible and testable.
Our findings, within the wider context, have real public health implications. Most people are exposed to cadmium mainly through cigarette smoke, contaminated food such as leafy vegetables and grains and, to a lesser degree, air and water near industrial areas. 46 Notably, national data reveal that smokers have much higher blood cadmium levels than non-smokers. The strongest links between cadmium and cancer are with the lungs, pancreas and bladder. 47 The risk of this toxic substance affecting other organs, including the brain, is concerning. Our demonstration that cadmium can activate key cancer-related factors in brain cells highlights the need for ongoing public health efforts to lower cadmium exposure. This includes enforcing environmental rules, promoting smoking cessation and monitoring cadmium levels in food. Moreover, our study suggests that molecular components of the SHH–GLI1-BMI1 axis could be considered as potential biomarkers of effect in populations with known high cadmium exposure, linking environmental exposure assessment to initial biological responses.
Conclusion
In conclusion, cadmium exposure may pose a significant risk of initiating and promoting brain tumours by disrupting critical regulatory pathways, particularly the SHH–GLI1 signalling cascade, which plays a vital role in maintaining normal brain cell function. By aberrantly activating the SHH pathway and upregulating GLI1, cadmium promotes uncontrolled cellular proliferation and tumorigenesis. Additionally, the overexpression of BMI1 under cadmium influence enhances the properties of cancer stem cells, contributing to tumour resistance and aggressiveness. Moreover, TP53 impairs the cell’s natural ability to repair DNA and undergo apoptosis, leading to the survival of genetically damaged cells that can further fuel tumour development (Figure 8).
(A) Cadmium (Cd²⁺) Competes with and Displaces Zinc (Zn²⁺) From Its Binding Site within the Zinc-finger Domain of GLI1. This Metal-ion Exchange is a Proposed Initial Molecular Event by Which Cadmium Exposure Could Directly Interfere with the Structure and Function of the GLI1 Transcription Factor. (B) Cadmium Exposure May Initiate and Promote Brain Tumour Development by Regulating SHH, GLI1, BMI1 and TP53.
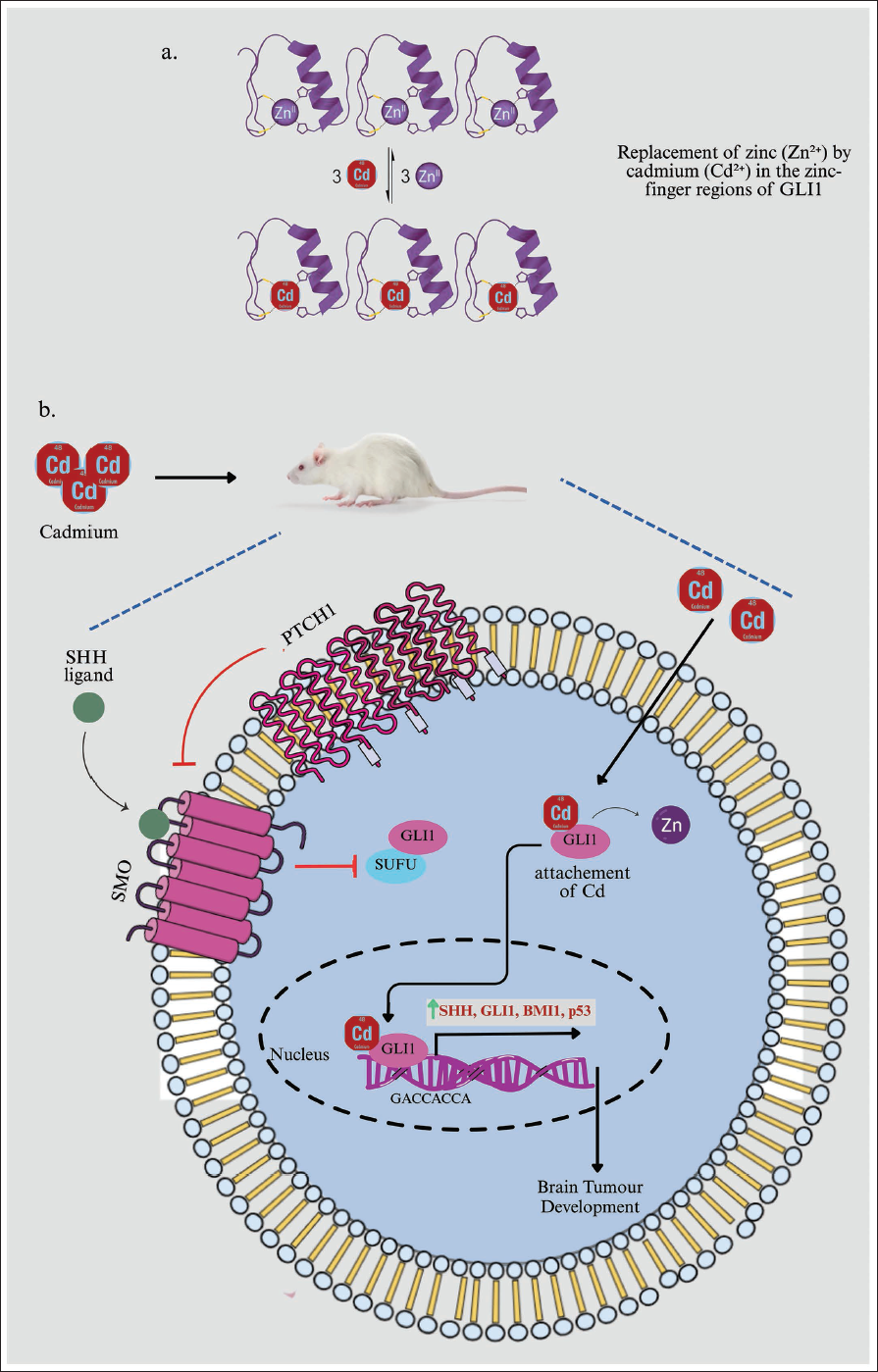
These combined effects underscore the importance of understanding environmental carcinogens such as cadmium in brain tumour research and public health. The disruption of these key molecular pathways by cadmium underscores the need for targeted therapeutic strategies to correct the deregulated signalling mechanisms. Furthermore, public health initiatives to reduce cadmium exposure could play a crucial role in preventing brain tumour development, especially in regions with high environmental cadmium contamination. Continued research into the molecular mechanisms underlying cadmium-induced tumorigenesis may offer new insights into preventing and treating brain cancers more effectively.
Authors’ Contributions
Swalih P Ahmed contributed to manuscript writing, materials preparation, methodology development, treatment of real-time RT-PCR and analysis of the results. Intezar Ali analysed the literature, wrote the manuscript and prepared Figures 1 and 8. Mehdi H Shahi provided the concept for this study and contributed to the design of the experiments, data analysis, writing the manuscript, securing research funding and overall supervision of the study.
Footnotes
Statement of Ethics
This study protocol had been approved by the IAEC and the Institutional Ethics Committee (IEC) of the Faculty of Medicine, Aligarh Muslim University, Aligarh 202002, India (Reference No. IECJNMC/1245).
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: This work was supported by a grant from the Indian Council of Medical Research, from the Government of India SRF (No. 3/2/2/69/2022/NCD-III).
Abbreviations
MB: Medulloblastoma, SHH: Sonic Hedgehog, GLI1: Glioma-associated oncogene 1, HH: Hedgehog, PTCH1: Patched 1, SMO: Smoothened, SUFU: Suppressor of fused, cDNA: Complementary DNA, DNA: Deoxyribonucleic acid RT-qPCR: Reverse transcription quantitative real-time PCR, PCR: Polymerase chain reaction, mRNA: Messenger RNA, TP53: Tumour protein 53, BMI11: B cell-specific Moloney murine leukaemia virus integration site 1.
Availability of Data and Materials
The authors confirm that the data supporting the findings of this study are available within the manuscript.