
Abstract
Background
The endogenous opioid system regulates diverse functions including pain, physiological processes and motivation. An earlier study had demonstrated increased cell proliferation in the neurogenic niche, that is, the ventricular-subventricular zone (V-SVZ) of adult male zebra finches (Taenopygia guttata) following administration of naloxone, a general opioid antagonist.
Purpose
To explore gene expression changes underlying the increase in cell proliferation in the V-SVZ of these birds after blocking opioid receptors (ORs) with systemic injections of naloxone. To assess whether opioid antagonism influences neuronal differentiation in the V-SVZ adjacent to the song control nuclei HVC and Area X, both critical for song learning and production.
Methods
Whole transcriptome microarray analysis of the V-SVZ was performed following naloxone or vehicle administration. The analysis identified 26 differentially expressed transcripts (fold change [FC] ≥ 1.5), including upregulated genes associated with neuroblast migration, neuronal differentiation, synaptic plasticity, and chromatin remodelling. Quantification of bromodeoxyuridine (BrdU) and doublecortin (DCX) positive cells was performed to determine proliferation and neuronal differentiation in the V-SVZ adjacent to HVC and Area X.
Results
Among differentially expressed transcripts, tgu-miR-124-201 (the precursor of miR-124-3p, a key regulator of neuronal differentiation), showed significant upregulation. Selected transcripts were validated by qRT-PCR, confirming their altered expression following naloxone treatment, compared to controls. Furthermore, we observed higher neuroblast density in the ventral V-SVZ adjacent to striatal song nucleus Area X, but not in the V-SVZ above the pallial song nucleus HVC, suggesting region-specific opioid modulation of neurogenesis.
Conclusion
Together, these results demonstrate that OR blockade may promote neurogenesis in adult songbirds and is accompanied by the upregulation of key transcripts, including miR-124-3p. Given the established role of miR-124 in promoting neuronal differentiation, its upregulation suggests that OR blockade may enhance neuronal differentiation via microRNA (miRNA)-mediated pathways. This finding provides the first evidence linking opioid modulation to miRNA-driven neurogenic processes in an adult avian brain.
Introduction
The two main niches that serve as the epicentres for adult neurogenesis in the vertebrate brain are the ventricular-subventricular zone (V-SVZ), lining the lateral ventricles in the telencephalon and the subgranular zone (SGZ) within the dentate gyrus of the hippocampus.1, 2 Whereas the rate of generation of new neurons in the V-SVZ varies among vertebrate species from fishes to primates, the magnitude of neurogenesis in the V-SVZ of adult songbirds such as zebra finches is significantly higher than that of other species.3, 4 In these birds, nascent neurons migrate and incorporate into various parts of the telencephalon, including some of the song control nuclei.5–7 Earlier studies have demonstrated that opioid receptors (ORs) are expressed by the V-SVZ of adult zebra finches, and blocking these receptors with systemic injections of the general opioid antagonist naloxone leads to an increase in cell proliferation in vivo.8–10 Similar results were observed when primary cultures of the adult zebra finch V-SVZ were treated with naloxone. 10 The rationale for this study was to move beyond earlier observations of naloxone-induced proliferation in the zebra finch V-SVZ and identify the underlying patterns of differential gene expression, while directly testing whether these changes also translate into measurable differences in neuronal differentiation. To this end, whole transcriptome microarray analysis, along with qRT-PCR validation, and quantification of new neuroblasts in the V-SVZ adjacent to HVC and Area X following OR blockade by naloxone, were performed. Besides an increase in transcripts encoding genes important for neuronal function and cell proliferation, a significant upregulation of a transcript for the precursor miRNA tgu-mir-124-201 was observed. These findings are interesting, since miR-124 inhibits cell proliferation and promotes differentiation into a neuronal fate via the SOX9 transcription factor in the subventricular zone.11, 12 Furthermore, miR-124 represses extracellular matrix proteins laminin subunit gamma 1 (LAMC1) and integrin beta-1 (ITGB1), leading to the differentiation of neuronal precursors into neurons. 13 Based on prior evidence, we hypothesised that naloxone-mediated OR antagonism would lead to an increase in the expression of the pro-differentiating microRNA, miR-124, in the neurogenic niche in adult zebra finches. This molecular alteration induced by the administration of an opioid antagonist may lead to the differentiation of newly proliferated cells into neuronal phenotypes. Thus, our study combined an exploratory whole transcriptome profiling with histology to evaluate whether naloxone treatment altered adult neurogenesis, with BrdU+/DCX+ cell density in the V-SVZ as the primary outcome.
Methods
Animals and Injections
All experimental procedures were performed in agreement with the recommendations of the Institutional Animal Ethics Committee of our institute, and in accordance with the guidelines of the Committee for Control and Supervision of Experiments on Animals (CCSEA), India (Protocol number Institute/IAEC/2018/143), which comply with international guidelines for animal welfare, and were reported in line with ARRIVE guidelines. 14 A total of 44 adult male zebra finches (>120 days post-hatch) were chosen from aviaries maintained at 12L:12D cycles of light and dark, with ad libitum access to feed (millets, eggs, greens) and water. Birds were weighed and then allowed to acclimatise for 24–48 hours in their cages before the injection regimen was started. No genetic modification, genotyping, surgical intervention, or invasive manipulation was performed during any step in the experimental procedures. Birds were handled gently to reduce stress during injections, and injections (naloxone, saline and BrdU) were administered at doses known not to cause adverse effects. 10 Group allocation and all subsequent procedures were conducted by one investigator. Blinding was not performed at any stage, as the procedures involved systemic injections and standardised downstream analyses. Twenty-two birds received intramuscular injections of 2.5 mg/kg body weight of the opioid antagonist naloxone (Naloxone hydrochloride dihydrate, Sigma, Cat. no. N7758) for four consecutive days, whereas 22 birds that acted as controls were injected with 0.9% saline (vehicle) during the same period. Injections were spaced 24 hours apart. The 2.5 mg/kg body weight dose of naloxone and the four-day treatment regimen were chosen based on a dose-response study, 10 which had demonstrated that this dose was effective in increasing cell proliferation in the V-SVZ. Whereas 18 birds (9 control and 9 treated) were used for microarray experiments, another set of 18 birds (9 control and 9 treated) was used for qRT-PCR experiments. For histology and immunolabelling experiments, another set of 8 birds was injected with naloxone (n = 4) or saline (n = 4) as described above, followed by an injection of 100 mg/kg body weight of BrdU (Bromodeoxyuridine, B9258, Sigma). Birds were observed daily for general health, activity, feeding behaviour, and plumage condition throughout the experimental period. All birds were sacrificed humanely for tissue collection, using an overdose of ketamine hydrochloride on the fourth day, two hours after the last injection.
Sample Preparation, Pre-processing and Whole Transcriptome Microarray Assay
For microarray and qRT-PCR experiments, sample preparation and pre-processing steps were performed in-house, under sterile, RNAse-free conditions. Birds were removed and cut into 1 mm thick coronal slices. Using a dissection microscope (Carl Zeiss™ Stemi 2000-C), a region approximately 250 µm lateral to the lateral ventricles was dissected from these slices, containing both the ventricular and subventricular zones (cf. 10 ). For both treated and control groups, V-SVZ from three birds were pooled and considered n = 1. The decision to pool samples for microarray and qRT-PCR analysis was based on protocols established in previous studies.8, 10 Total RNA was isolated by the guanidinium thiocyanate-phenol-chloroform or TRIzol method. Extracted RNA (n = 3 samples per group: control and treated) was treated with TURBO™ DNase (Invitrogen) to remove any traces of genomic DNA, quantified and checked for RNA integrity (RIN ≥ 7). The microarray study was conducted collaboratively: laboratory procedures and partial analyses were performed at Innovative Life Discoveries Private Limited, Manesar, India, whereas data interpretation and additional analyses were completed at the National Brain Research Centre, Manesar, India. For whole transcriptome profiling, 500 ng of total RNA from each sample was processed using the Affymetrix WT PLUS Reagent Kit. First-strand cDNA synthesis was carried out using random primers incorporating a T7 promoter sequence (2 hours), followed by second-strand cDNA synthesis to generate double-stranded cDNA (1 hour 10 minutes). This was transcribed in vitro using T7 RNA polymerase, producing amplified complementary RNA (cRNA), which was subsequently purified using magnetic purification beads. A 15 µg aliquot of purified cRNA was then reverse-transcribed into a sense-strand cDNA, using second-cycle primers and dNTP mix containing dUTP. The cRNA template strand was hydrolysed with RNase to obtain a single-stranded second-cycle cDNA. After purification and quantification, single-stranded cDNA (5.5 µg) was subjected to enzymatic fragmentation for an hour using a mixture of uracil-DNA glycosylase (UDG) and apurinic/apyrimidinic endonuclease-1 (APE1) enzymes, which cleaved the cDNA at dUTP residues. The fragmented cDNA was end-labelled with a biotinylated dideoxynucleotide in a reaction catalysed by terminal deoxynucleotidyl transferase (TdT). The biotinylated cDNA was mixed into a hybridisation cocktail (200 µL) containing hybridisation buffer, control oligo B2, and hybridisation controls. This mixture was loaded onto GeneChip Zebra Finch 1.0 ST Arrays, incubated at 45°C for 16 hours with rotation at 60 rpm. Post-hybridisation, arrays were washed on the Affymetrix Fluidics Station 450, stained with stain cocktails (available in the kit), and scanned using the GeneChip Scanner 3000 7G to capture fluorescence intensity values. Raw data files were processed for a background adjustment, quantile normalisation and summarisation in the Affymetrix Expression Console. The filtered probes were annotated using the Ensembl-based taeGut3.2.4 assembly.
Identification of Differentially Expressed Genes and Hierarchical Cluster Analysis
A moderated t-test was applied to the signal intensities of the filtered probes (Agilent GeneSpring software) for both control and naloxone-treated groups (n = 3; three sets of repeats; all samples were included in the analysis). To consider a gene as differentially expressed and statistically significant, the p < .05 and log 2 (fold change [FC]) ≥ 0.59216535 were selected as the cut-off points and represented by volcano plots. The normalised intensities of the differentially expressed and statistically significant genes with FC ≥ 1.5 were input into the Genesis© tool (
Quantitative Real-time Polymerase Chain Reaction (qRT-PCR) Analysis
For qRT-PCR validation, differentially expressed transcripts from the microarray dataset were selected using published literature and database annotations, focusing on genes encoding identifiable proteins or miRNA, particularly those participating in pathways related to cell division and differentiation. Specific Taqman™ microRNA assays (Assay ID: 000239, cat. no. 4427975) and miR-124-5p (Assay ID: 241919_mat, cat. no. 4440886) and specific primer sequences were used to detect the genes of interest by qRT-PCR. The detection of genes was performed on RNA extracted from tissue from another set of 18 birds (9 controls and 9 treated; n = 3, 3 sets of repeats; all samples were included in the analysis), adhering to manufacturer’s instructions from Taqman™ (Invitrogen) with the U6 snRNA 3p (Assay ID: 001973, cat. no. 4427975) serving as the reference standard gene. For other genes of interest, the glyceraldehyde 3-phosphate dehydrogenase (GAPDH) gene was used as an endogenous control. The relative amount of each gene of interest was normalised against U6 snRNA or GAPDH, and the FC for each gene in naloxone-treated samples compared with vehicle controls was calculated using the 2−∇∇Ct method.
Histology, Immunostaining, Imaging and Colocalisation Analysis
Following sacrifice and perfusion of adult male zebra finches (n = 8; 4 control and 4 treated birds), brains were removed, cryopreserved in 30% sucrose and sectioned coronally into five sets of 30 µm-thick serial sections using a cryostat (CM 3050 S, Leica). To identify song control nuclei and other anatomical markers, one series of the sections was stained with Thionin (Nissl). The density of total proliferating cells was also assessed as a secondary outcome, providing a broader measure of proliferation. To evaluate neuronal differentiation, immunolabelling with antibodies against BrdU and DCX was performed. The replication level (n = 3-4 birds per group) was guided by suitability for meaningful statistical comparisons. Three sections were selected from each bird, spanning the coronal planes containing the V-SVZ adjacent (a) Area X (n = 4; all samples included) and (b) HVC (n = 3; out of four birds administered with naloxone/saline, one sample per group was excluded due to tissue quality issues). 15 The sections were rinsed in 0.1 M phosphate-buffered saline (PBS), subjected to antigen retrieval (Vector Laboratories, H-3300, pH 6) and treated with 2N HCl to denature DNA. Sections were then rinsed in 0.1M borate buffer, followed by quenching with 1% hydrogen peroxide and rinsing in 0.1% Triton-X-PBS. Next, sections were blocked in 10% normal goat serum (NGS, Vector Laboratories, S-1000), 2% bovine serum albumin (BSA), and 0.1% triton-X-PBS for 2 hours. Blocking was followed by incubation in a solution containing the monoclonal anti-BrdU antibody (1:250, Sigma, Cat.no. B8434) and rabbit polyclonal anti-DCX antibody (1:1000, Abcam, ab18273 16 ) diluted in 5% NGS, 2% BSA, and 0.1% triton-X-PBS at 4°C for 36–48 hours. Excess antibody was rinsed off with PBS, and the sections were incubated with a cocktail of secondary antibodies [goat anti-mouse Alexa-488 (1:500, Vector Laboratories, A11001) and goat anti-rabbit Alexa-594 (1:1500, Vector Laboratories, A11012)] for two hours at room temperature. After a final rinse with PBS, sections were mounted on gelatin-coated slides and cover-slipped with Vectashield® antifade mounting medium with DAPI (Vector Laboratories, H-2000). For each immunolabelled section, images of BrdU-positive nuclei and DCX-positive neuroblasts within the region extending up to approximately 185 µm lateral to the V-SVZ [cf.10, 15] were acquired as a series of z-stacks at 60X, with a step size of 0.5 µm and a resolution of 1,024 × 1,024 pixels using a Nikon confocal microscope (A1 HD25) to capture three-dimensional distribution.
Newly proliferated groups of BrdU-positive cells were quantified automatically in maximum projection images generated from the z-stacks using a custom-designed cell counting macro in the ImageJ software. To minimise background noise and enhance signal clarity, we applied pseudo flat-field correction (blurring factor: 1,500), followed by 3D median filtering (2 × 2 × 2 kernel). This was followed by applying watershed segmentation and auto-thresholding (Yen method) to delineate individual cells more accurately, whereas particle analysis (size range: 5–∞, circularity: 0.05–1.00) helped exclude artefacts. A semi-automated object-based colocalisation analysis (OBCA) method 17 was used to count BrdU-positive cells co-labelled with DCX (BrdU+/DCX+ cells). First, thresholding was applied to include only brightly labelled cells near the V-SVZ region. Then, Plugin 1 transformed the z-stacked multichannel confocal image into an optimised binary and grayscale output image, in which 3D spatial information was projected onto a 2D plane for easier visualisation. This processed image displayed the overlapping BrdU and DCX signals using outlines to represent the colocalised regions, enabling the identification of colocalised cells. Colocalisation was assigned based on the presence of DCX signal with BrdU+ cells and detected automatically with Plugin 2 (Colocalisation Object Counter). Manual verification was also performed to check for accuracy; wherever cells were closely clustered, colocalised signals were manually verified across the Z-stack and also using the 3-D visualisation mode available in the Nikon confocal NIS-Elements software. For each bird, the density of newly proliferated cells and neuroblasts was averaged from three sections at the level of Area X and HVC and expressed as the density of BrdU+ cells and BrdU+/DCX+ cells per mm 2 of the V-SVZ region analysed.
Statistics
For qRT-PCR experiments, Student’s t-test (unpaired, two-tailed) was performed on the relative FCs in GraphPad Prism (version 10.0.0, GraphPad Software, Boston, Massachusetts, USA) to analyse differences in the normalised gene expression. The statistical comparisons between the means of the counts from the two groups were made using student’s t-test (unpaired, two-tailed) in GraphPad Prism. The data were represented as mean ± standard error (SEM), and the significance level was set at p < .05.
Results
Differentially Expressed Transcripts
Microarray analysis of total RNA from the V-SVZ of naloxone-treated birds and control birds revealed a total of 427 significantly differentially expressed transcripts (Figure 1a; Supplementary Table 1). A moderated t-test was applied to this data set, and a p value cut-off (p < .05) was used to identify significantly differentially expressed transcripts. Furthermore, a cut-off of log 2 FC ≥ 0.59216535, which corresponds to an FC ≥ 1.5, was used to identify biologically meaningful changes.18, 19 Following this analysis, the expression of 16 transcripts was found to be significantly upregulated (Supplementary Table 2), whereas that of 10 transcripts was significantly downregulated in V-SVZ (Supplementary Table 3), post-treatment with naloxone compared to vehicle (control) (Figure 1b and 1c). The Ensembl IDs of the transcripts and the genes encoding them, along with their expression FCs, are summarised in Supplementary Tables 2 and 3.
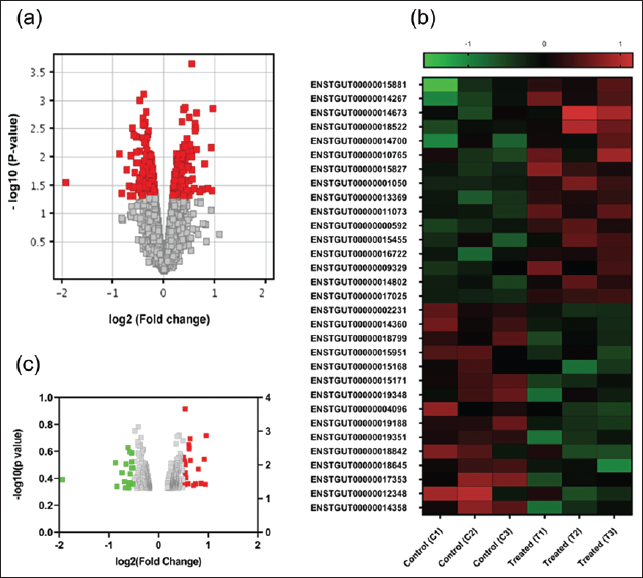
Blocking ORs Leads to an Upregulation of Genes that Influence Neuronal Function in the V-SVZ
The significantly upregulated transcripts in the V-SVZ of the birds treated with naloxone are known to be encoded by genes involved in biological functions, including cell-cell signalling, mRNA processing, chromatin remodelling, cell division, cell growth and survival, regulation of apoptosis, protein transport, and exocytosis. For qRT-PCR validation, we focused on genes encoding identifiable proteins or miRNAs, particularly those linked to cell division and differentiation pathways. Selected upregulated transcripts and their FCs (in the microarray dataset) following naloxone administration included tumor necrosis factor alpha induced protein 2 (TNFAIP2; FC = 1.53), Adaptor Related Protein Complex 3 Subunit beta 2 (AP3B2; FC = 1.8), pre-miRNA-124 (mir-124-201; FC = 1.7), TELO2 Interacting Protein 1 (TTI1; FC = 1.9) and Chromatin Accessibility Complex Subunit 1 (CHRAC1; FC = 1.9), whereas the induced myeloid leukaemia cell differentiation protein (MCL-1; FC = 1.8) was selected among the significantly downregulated transcripts). A qRT-PCR analysis of the selected upregulated genes revealed that their FCs were similar to those obtained in the microarray analysis: tgu-miR-124-3, FC = 1.6 ± 0.0577, AP3B2, FC = 1.8 ± 0.173; TTI1, FC = 1.7 ± 0.208; CHRAC1, FC = 3.27 ± 0.769 (Figure 2a–2g). Furthermore, we observed a higher level of TNFAIP2 in the V-SVZ of naloxone-treated birds compared to controls (FC = 2.63 ± 0.857), although this difference did not reach statistical significance. Despite the microarray analysis indicating a significant downregulation of the MCL-1 transcript (FC = 1.8), the qRT-PCR assay demonstrated no significant change in the gene expression levels in the V-SVZ of naloxone-treated birds compared to controls (FC = 1.03 ± 0.376).
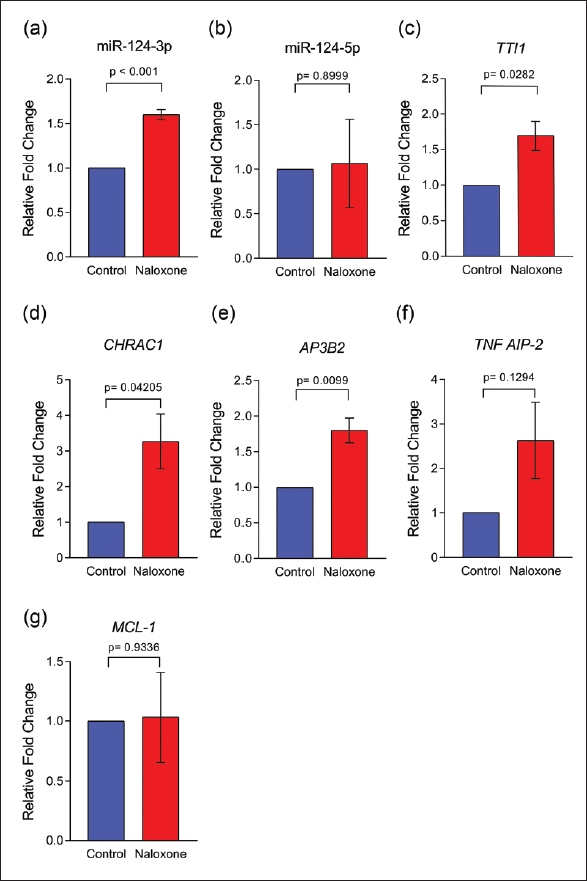
MicroRNA-124 may be Involved in the Opioid-Modulated Neurogenesis in the Zebra Finch V-SVZ
We found a precursor miRNA transcript, that is, tgu-mir-124-201 (miRNA Entry for MI0013699;
Effects of Naloxone on Neuronal Differentiation in the Adult Zebra Finch V-SVZ
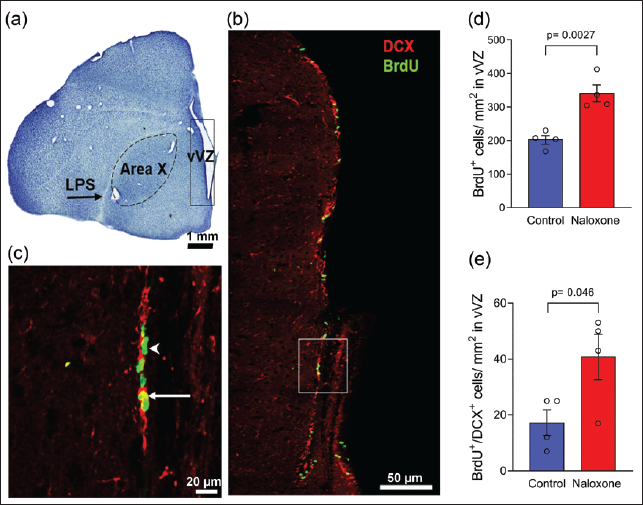
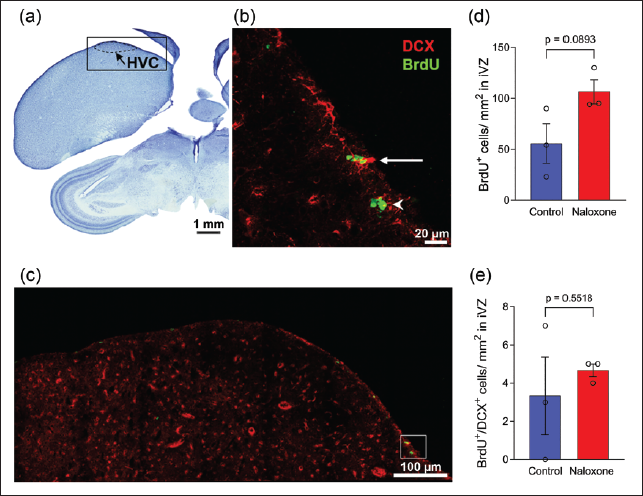
Previous studies have reported that zones of maximum cell proliferation called ‘hot spots’ are located in the V-SVZ adjoining the song control nuclei Area X and HVC and the anterior commissure in male zebra finches.15, 23 Here, we demonstrated that ventral V-SVZ (VVZ) adjacent to Area X and the intermediate V-SVZ (IVZ) dorsal to HVC contained BrdU-labelled cells (green, arrowheads) and neuronal precursors co-labelled for BrdU and DCX (yellow, arrows, Figures 3 and 4, respectively). The density of BrdU+ cells was significantly higher in VVZ adjacent to Area X of the naloxone-treated group compared to controls (341.0 ± 25.0 cells/mm 2 vs. 202.5 ± 12 cells/mm 2 , n = 4, p = .0026). Significant differences were also found in the density of BrdU+/DCX+ cells (40.8 ± 8.2 cells/mm² vs. 17.3 ± 4.6 cells/mm², n = 4, p = .0463) at this level. Despite an increase in cell proliferation in the IVZ adjacent to HVC, we found no significant differences in the density of either BrdU+ cells (55.7 ± 19.3 cells/mm² vs. 106.3 ± 11.8 cells/mm², n = 3, p = .0893) or BrdU+/DCX+ cells (4.7 ± 0.3 cells/mm² vs. 3.3 ± 2 cells/mm², n = 3, p = .5518) between naloxone-treated birds and controls.
Discussion and Conclusions
The Endogenous Opioid System Modulates Gene Expression in the V-SVZ of Adult Male Zebra Finches
A whole transcriptome microarray on the adult male zebra finch V-SVZ revealed that systemic administration of the general opioid antagonist naloxone significantly upregulated the expression of 16 transcripts and downregulated the expression of 10 transcripts. Since the V-SVZ houses neural precursors, ependymal cells, neuroblasts, microglia, and astrocytes, 24 the differentially expressed transcripts observed in this study likely represent the altered gene profiles of these cell populations in response to naloxone treatment. Furthermore, these results were validated by performing qRT-PCR analysis on selected transcripts, suggesting that their upregulation is indeed a consequence of OR blockade. Overall, these findings suggest that the endogenous opioid system modulates gene expression in the V-SVZ of adult male zebra finches.
Altered Expression of Specific Genes in the V-SVZ in Response to Naloxone Treatment
We found that the expression of the pre-miRNA tgu-mir-124-201 transcript was significantly elevated in the V-SVZ of adult male zebra finches following naloxone treatment. The processing of precursor miRNAs into mature miRNAs involves the generation of duplexes containing two RNA strands, namely miR-124-3p and miR-124-5p.20, 21 Subsequent qRT-PCR analysis of mature miR-124 in the neurogenic region after naloxone treatment confirmed the successful processing of tgu-mir-124-201 into mature miR-124-3p and miR-124-5p. The observed upregulation of miR-124-3p (FC = 1.6 ± 0.0577) but not of miR-124-5p (FC = 1.08 ± 0.498) in the naloxone-treated V-SVZ suggests that miR-124-3p is more stabilised and functionally relevant in this context.
MicroRNA-124 is the most abundantly expressed miRNA in the brain and favours the differentiation of neural precursor cells into neurons.25, 26 A constant interplay between miRNAs and the endogenous opioid system must be in effect in order to maintain stable levels of neurogenesis in the brain.27, 28 Our findings suggest that the levels of miR-124-3p, transiently upregulated during exposure to naloxone, favoured a shift towards the neuroblast stage from the neuronal precursor cells in the V-SVZ. Furthermore, a human postmortem study demonstrated that opioid use disrupts miRNA–mRNA networks, affecting pathways related to tissue development, morphogenesis, MAPK signalling, and cell-type-specific functions in astrocytes, neurons, and endothelial cells. 29 In all, miR-124 can be considered a conserved mediator linking OR activity to transcriptomic regulation, potentially influencing neurogenesis, vascular function, and inflammatory pathways across species.
We also found that the transcript encoding CHRAC1 exhibited a 1.9-fold upregulation in the V-SVZ of naloxone-treated birds versus controls. CHRAC1 belongs to a group of DNA-binding histone-fold proteins and plays a pivotal role in regulating the accessibility of DNA to the transcriptional machinery. 30 Therefore, the higher expression of this gene suggests increased chromatin remodelling and transcriptional activation in the neurogenic zone of zebra finches after naloxone treatment.
Furthermore, there was a FC of 1.7 in the AP3B2 transcript in naloxone-treated V-SVZ samples compared to controls. Mutations of AP3B2 are known to be linked to microcephaly, developmental impediments, seizures, and intellectual disability, 31 suggesting a potential role of AP3B2 in neuroblast migration and differentiation. Our results, therefore, suggest that blocking ORs may affect AP3B2 expression in the V-SVZ, thereby modulating neurogenesis in adult male zebra finches.
Besides miR-124, we found that a transcript encoding TTI-1 was also upregulated by a FC of 1.9 in the V-SVZ of naloxone-treated birds versus controls. TTI-1 interacts with mTOR (mechanistic target of rapamycin), a serine/threonine protein kinase of the PIKK protein family and an essential regulator of translation, cell proliferation, and survival, as discussed in a review. 32 An interplay between the µ-OR system and mTOR signalling pathway has been implicated in neuronal development, neuronal survival, and synaptic plasticity. 33 Furthermore, the mTOR signalling cascade is activated in the auditory forebrain regions [caudomedial nidopallium (NCM) and caudomedial mesopallium (CMM) and the primary auditory forebrain (Field L)] of adult male zebra finches involved in song recognition, 34 which are known to incorporate new neurons. 35 As a caveat, earlier studies demonstrated that general opioid antagonists led to the inactivation of the PI3K/AKT/mTOR pathway, thereby reducing cell proliferation in cancer cells, 36 whereas opioid agonists like morphine promoted cell proliferation and migration in squamous cell carcinoma via activating the PI3K/AKT/mTOR signalling pathway. 37 However, in our experiment, four days of opioid antagonist treatment led to increased expression of the TTI1 transcript, indicating the involvement of the activated mTOR signalling cascade in the V-SVZ cells versus that in controls, which may reflect species differences.
Changes in Adult Neurogenesis as a Result of Altering Opioid Modulation in the Zebra Finch Brain
Our results demonstrated that naloxone administration induced an increase in cell proliferation (cf. 10 ) as well as an increase in the density of new neuroblasts in the V-SVZ lining the lateral ventricles adjacent to the song control nuclei Area X and HVC, versus that in controls. 23 However, our results indicated that the increase in density of newly proliferated cells and neuroblasts was significant only at the level of Area X, suggesting that opioid modulation not only affects cell proliferation but also promotes the generation of neuronal precursors, in a region-specific manner, in the adult zebra finch V-SVZ. As a caveat, a study demonstrated that primary neural stem cells in mouse embryos expressed low levels of ORs during the first 53 days of differentiation. This study also reported that naloxone was effective in promoting neurogenesis only during the initial stages (Days 0–5) and inhibited neurogenesis during later stages (Days 11–15) of differentiation, and that naloxone promotes neurogenesis via pathways independent of ORs. 38 Although it has been shown that ORs are robustly expressed in the V-SVZ of adult male zebra finches, 10 we cannot rule out the possibility of naloxone promoting neurogenesis by means other than by directly binding to ORs. Additionally, there may be differences in the effects of naloxone on neurogenesis depending on the developmental age of neural stem cells from the V-SVZ, as well as species differences.
This study has a few limitations. First, modest sample sizes (n = 3–4 per group) may be the reason why variability was observed in the histological measures across biological replicates. Furthermore, although zebra finches are a powerful model for studying adult neurogenesis due to their robust V-SVZ activity and well-defined song nuclei, their translational relevance to human neurogenesis is indirect. The extent of neurogenesis in the adult human brain is considerably lower, and the regulation of opioid signalling, as well as the expression and regulatory networks of miRNAs such as miR-124, may be organised differently. Finally, naloxone was administered systemically, which can prevent the precise dissection of local versus global effects of OR blockade on neurogenic niches.
In all, findings from our study suggest that opioid modulation in the V-SVZ of zebra finches leads to upregulation of miR-124 along with other important genes involved in neuronal proliferation and differentiation. The modulation of neurogenesis by opioid antagonism appears to be robust in the ventral V-SVZ that supplies new neurons to song control region Area X, compared to the V-SVZ region that supplies new neurons to HVC. Future studies will focus on the mechanistic details of how these genes may contribute to song learning and vocalisation in adult male songbirds.
Footnotes
Acknowledgements
We would like to acknowledge Dr Sarbani Samaddar for providing insightful comments on this manuscript.
Authors’ Contribution
UD carried out the experiments, analysed the data and wrote the initial drafts of the manuscripts. SB and SI designed the experiments, reviewed and edited the manuscript.
Patient Consent
Not applicable.
Statement of Ethics
All experimental procedures were performed in agreement with the recommendations of the Institutional Animal Ethics Committee of the National Brain Research Centre, Manesar (NBRC), in accordance with the guidelines of the Committee for Control and Supervision of Experiments on Animals (CCSEA), India, which comply with international guidelines for animal welfare.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors disclosed receipt of the following financial support for the research, authorship and/or publication of this article: The work was supported by core/intramural funds from the National Brain Research Centre (NBRC), Manesar, provided to the corresponding author (SI).
Data Availability Statement
All data associated with this research are provided in the manuscript, the supplementary information and tables.
Supplemental Material
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.