
Abstract
Background
The interplay between emotional stimuli and cognitive control is crucial to understanding adaptive behaviour, particularly in young adolescents whose executive functioning and emotional regulation are still developing. While prior research has examined these influences, the underlying neurobehavioural correlates remain insufficiently understood.
Purpose
This study investigated the influence of attentional demands on emotional valence in young Indian adolescents through the analysis of behavioural responses and event-related potentials (ERPs) in an affect-primed flanker task.
Methods
Forty-four young adolescents (68.18% female, aged 13–14) participated in a flanker task with congruent and incongruent trials at two levels of difficulty, wherein each flanker trial was presented immediately after time-locked affective picture stimuli (positive/negative/neutral valence). Electroencephalography recordings were analysed to identify ERP components alongside an examination of the behavioural data (reaction times/accuracy). Robust statistical methods addressed gender-specific ERP and behavioural patterns.
Results
ERP-based mass-univariate analysis revealed significant differences between positive and negative stimuli at early (88–140 ms) and late (352–412 ms) time windows. Negative stimuli elicited stronger N1 amplitudes, while positive stimuli demonstrated enhanced P3b amplitudes. This differentiation aligns with dual-processing models of emotion and attention, whereby negative stimuli are processed rapidly at an early stage, while positive stimuli engage deeper cognitive processing at later stages. The absence of a typical late positive potential component highlighted the prioritisation of task demands over emotional processing, suggesting that limited attentional resources were allocated to sustained emotional evaluation. Gender differences were noted, with females demonstrating slower reaction times yet higher accuracy, as well as more positive ERP waves in fronto-temporal regions, regardless of valence.
Conclusion
The findings underscored the temporal dynamics of emotion–attention interactions during young adolescence, highlighting the salience-driven nature of early attentional processes and the role of emotional valence in cognitive engagement. Furthermore, gender differences suggested distinct strategies for emotion–cognition integration.
Introduction
Emotions shape how we perceive and respond to the world, influencing attention to either enhance or hinder cognitive performance. Whether a stimulus demands immediate focus or is ignored depends on its emotional significance, making attentional control a critical adaptive function. 1 This emotional significance—determined by valence and arousal—engages neural mechanisms that prioritise salient information, directing cognitive resources towards or away from a task. 2 This dynamic is particularly evident in conflict resolution tasks, where emotionally charged stimuli can either facilitate or disrupt attentional control.3, 4
Despite advances in understanding these processes in adults, they remain underexplored in adolescents. Adolescence is a critical period of emotional sensitivity and cognitive development. Therefore, this study aimed to fill this gap by understanding how emotion and cognition interact in this formative stage, providing insight into mechanisms governing emotional regulation and executive control.5–7
Emotional Stimuli and Cognitive Control
Emotional regulation and executive functioning intertwine, influencing processes such as attentional control and error monitoring.8, 9 Specifically, dual-process theories challenge the traditional separation between ‘hot’ emotional and ‘cold’ cognitive processing by suggesting shared, overlapping neural circuits.3, 4 The cognitive load theory posits that individuals have limited cognitive resources; thus, the extent to which emotional stimuli are affected by a task depends on the available attentional capacity. Within this, the broaden-and-build theory suggests positive emotions expand attentional scope and flexibility, 10 while negative emotions narrow them.11, 12 Conversely, the threat-capture hypothesis states that negative—especially threatening—stimuli automatically capture attention, disrupting or enhancing performance depending on task demands.13, 14 Other evidence suggests that emotional processing may vary based on whether attention is explicitly directed towards or away from emotional stimuli.15–17
Research and empirical findings are mixed. Some studies suggest negative emotional stimuli slow reaction times (RTs),18–20 while others indicate enhancement or no effect.21–23 These inconsistencies highlight the need for methodologies such as event-related potentials (ERPs) to capture behavioural and neural dynamics. Notably, early neural responses—particularly the N1 component—remain understudied in adolescents despite their relevance in attentional allocation during emotionally charged tasks. 24 Emotional interference slows responses due to preferential stimulus processing,25, 26 further shaped by conflict monitoring mechanisms.27, 28
Flanker Task for Measuring Cognitive Control
The flanker task is a well-established paradigm for assessing executive control, as it requires responses to a central target while ignoring surrounding distractors. 29 Specifically, congruent conditions facilitate faster and more accurate responses, whereas incongruent conditions result in slower responses and higher error rates.30, 31 When paired with emotional stimuli, the task allows investigation of emotion–cognition interactions. Research shows that emotional distractors can impair performance by diverting attentional resources.31–33 Conversely, some reports conclude that emotional targets, particularly those with threatening valence, may enhance performance under certain conditions.31, 34 Additionally, emotional stimuli can be presented before trials (challenging attentional reorienting)35, 36 or throughout trials (taxing attentional sustainment and contextual processing),37–39 providing valuable insights into how attention to emotional valence influences task performance.
ERPs and Emotional–Cognitive Processing
ERPs provide a powerful tool for examining the temporal dynamics of emotion–cognition interactions. The N100 (N1), peaking approximately 100 ms after stimulus in fronto-central regions, reflects early attention allocation and is modulated by emotional valence and arousal.40–42 Emotional stimuli can influence both the amplitude and latency of N1, thereby highlighting its role in regulating attention during early cognitive processing.43, 44 It is also sensitive to attentional focus, task demands and stimulus predictability.45, 46 Later components, such as the P3b (~300 ms), reflect attentional resource allocation and working memory updating, modulated by emotional content. 47 Emotional stimuli with a high valence or arousal elicit larger P3b amplitudes,48, 49 suggesting prioritised processing of emotionally salient information. 50 Its amplitude is also influenced by factors such as stimulus probability, task complexity and the allocation of attentional resources.47, 51–54 An even later component, the late positive potential (LPP, 400–800 ms), is modulated by both the emotional valence of stimuli and the cognitive demands of concurrent tasks. 55 Its amplitude is typically enhanced by high-arousal stimuli, 56 but lowered under cognitive load. 57
Gender Differences in Emotional–Cognitive Processing
Neuroimaging studies suggest robust sex differences in emotional processing, particularly in the amygdala and prefrontal cortex.58, 59 Women typically exhibit greater left amygdala activation in response to negative stimuli, indicating enhanced emotional regulation and memory encoding. 60 They also show greater left hippocampal activation regardless of emotional valence.61, 62 Conversely, men show greater activation in the right amygdala and inferior frontal gyrus for positive stimuli, suggesting a possible reward-processing bias.58, 60
Behaviourally, women often exhibit slower RTs but higher accuracy (ACC), suggesting a cognitive strategy that prioritises precision over speed,63, 64 especially under conditions of emotional interference.65, 66 In contrast, men generally demonstrate faster RTs at the expense of ACC, indicative of a more impulsive processing style.63, 67 These traits may contribute to sex differences in engagement with emotionally complex environments, particularly during adolescence when emotion regulation strategies are still maturing.
Present Study
Understanding the interaction between emotional stimuli and cognitive control is critical for uncovering the mechanisms underlying adaptive behaviour. Although significant progress has been made in adult populations, this research cannot be reliably extrapolated to adolescence as it is characterised by heightened emotional reactivity and rapidly maturing cognitive control. Previous research has typically either used an in-task- or a non-image-based affective stimulus interruption within the flanker framework. The present study employing neither, therefore, offers a novel paradigm in the young Indian adolescent context. Studying these dynamics with ERPs provides high temporal resolution. Early components such as the N1 reveal attentional allocation, while later components such as the P3b show cognitive processing of emotionally salient stimuli. Previous research has indicated that emotional stimuli modulate N1 and P3b amplitudes, but inconsistencies require further study, particularly employing paradigms that engage both emotional and cognitive processes. By integrating behavioural and ERP measures, this study seeks to address the following research question:
‘How does a cognitive interruption via a flanker task modulate neural and behavioural responses to emotional valence in young adolescents?’
This question aligns with dual-process models of emotion and cognition,4, 68 emphasising the role of attentional demands in modulating processing and response to emotional stimuli.
Methods
Participants
Forty-four young adolescents (68.18% female) aged 13–14 years (M = 13.64) participated. All were reported as healthy with normal or corrected-to-normal vision and no prior experience with any affective picture paradigm or the flanker task. Written informed consent was obtained from all participants and their parents/guardians prior to involvement.
Stimuli
Affective Imagery
Sixty images (20 per valence) were randomly selected from the International Affective Picture System (IAPS), 69 based on suitability for adolescents. IAPS images whose valence ratings ranged from 1.47 to 8.05 (M = 5.23) were categorised based on valence ratings: negative images (valence <5), neutral images (5–5.39) and positive images (>5.39).
Cognitive Interruption
An arrow flanker task immediately followed the presentation of the affective imagery. 29 Congruent and incongruent trials were randomly presented in a 7:5 ratio to amplify the flanker congruency effect. 70 Moreover, trials were equally divided by difficulty. In easy trials, a single row of arrows was presented; in difficult trials, five rows were shown, thereby creating a ‘2 × 2’ factorial structure of ‘Congruence × Difficulty’.
Experimental Paradigm and Procedure
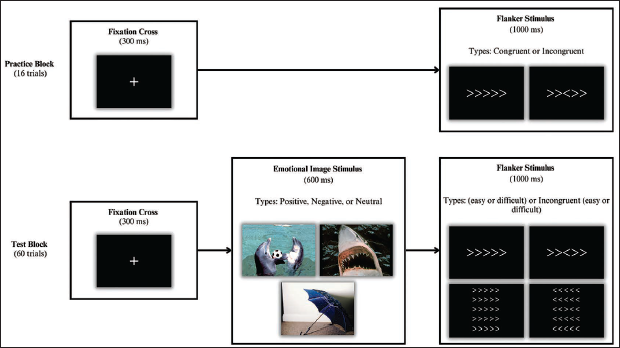
Designed on E-Prime 3 software, 18 our design interrupted affective processing with the flanker task rather than embedding an affective component within it. Participants were seated 70 cm away from a 17-inch computer (~8.9° × 13.2° visual angle span for both stimuli-flanker and affective picture). They were instructed to focus on a central fixation cross and to respond accurately to the flanker task by identifying the direction of the middle arrow. Responses were recorded using Psychology Software Tools Chronos buttons: the ‘1’ key for a left-pointing arrow and the ‘5’ key for a right-pointing arrow. Participants began with a 16-trial practice session without images, incorporating only easy flanker tasks. After receiving performance feedback, the main experiment commenced (60 trials), each beginning with the fixation cross (300 ms), followed by the presentation of a positive, negative or neutral image (600 ms). Subsequently, the flanker target appeared for 1000 ms or until a response was recorded (Figure 1).
Schematic Representation of the Experimental Paradigm. The Study Began With a Practice Block (Top) Where Each Trial Began With a Fixation Cross (300 ms), Followed by Either Simple Congruent or Incongruent Flanker Stimuli (1000 ms or Until Response). The Test Block (Bottom) Included One of Three Types of Affective Images (600 ms) After the Fixation Cross (300 ms), Followed by One of Four Possible Flanker Stimuli (1,000 ms or Until Response).
Electrophysiology
Electroencephalography (EEG) Recording
EEG data were acquired using a 64 Ag/AgCl electrode Easycap system (Brain Products GmbH), following the extended International 10–20 system. 71 Signals were amplified via LiveAmp 64 (Brain Products GmbH; 131-Hz third-order sinc filter), with FCz as the reference and AFz as the ground electrode, using Brain Products Recorder (v1.25.0001), at a sampling rate of 500 Hz.72–74 Besides, electrode impedances (5–15 kΩ) were verified for each session.72, 73
EEG Data Preprocessing
Preprocessing was conducted in MATLAB (vR2021a) using EEGLAB v2023.1 75 and ERPLAB v10.04. 76 First, event codes were adjusted by 28 ms to compensate for display lag, and inactive intervals (≥2,000 ms without an event) were excluded, with a 500 ms buffer before the first event and 1,500 ms after the last. Next, the preprocessing RELAX pipeline, 77 incorporating EEGLAB 75 and FieldTrip functions, 78 was employed. A fourth-order Butterworth bandpass filter (0.25–70 Hz) was applied, followed by power noise reduction using a zapline plugin. 79 Faulty electrodes were identified and removed via the PREP pipeline’s ‘findNoisyChannels’ function.79, 80 Criteria for removal included excessive muscle activity, 81 extreme kurtosis, voltage drift,81, 82 highly unlikely voltage distributions and extreme outlier amplitudes. 83 If more than 20% of electrodes were flagged for removal, only the poorest-performing 20% were excluded. Muscle activity, blinks and eye movement were addressed using a multichannel Wiener filter.83, 84
Thereafter, artefact removal was complemented by normalisation via independent component analysis (ICA) using fastICA. 85 Artifactual components were identified with ICLabel 86 and rejected via wavelet-enhanced ICA. 87 Continuous data were transformed to scalp space, enabling spherical interpolation to replace missing electrodes. Following the artefact-cleaning procedures, a high-pass filter (0.2 Hz) and low-pass filter (30 Hz) were applied to focus on the frequency range most relevant to ERP components. This step improved the data for ERP extraction and visualisation by removing leftover low-frequency drift and high-frequency noise. Lastly, the online reference FCz was recovered employing spherical interpolation and a mean-mastoid re-referencing was applied to all channels.72, 73 Thereupon, in ERPLAB, 76 cleaned data were segmented into 600 ms epochs, time-locked to image presentation, with a 200 ms prestimulus baseline. Each epoch underwent artefact rejection to eliminate large, commonly encountered artifactual potentials. 88 This process utilised simple voltage thresholds (SVTs) and moving window peak-to-peak amplitude thresholds (MWTs). SVTs identified epochs with extreme voltage values exceeding ±100 µV, while MWTs detected rapid voltage changes with a threshold of 100 µV within a 200-ms window, moving in 100-ms steps. The number of rejected epochs for each participant adhered to the recommended predetermined exclusion criteria.89, 90
ERP Data Analysis
ERPs for each subject were averaged in ERPLAB, 76 with an analytic standard measurement error computed for data quality for each of the subject-wise averaged sets.76, 91 These were then grand-averaged, and the subject-averaged ERP sets underwent mass-univariate toolbox (MUT) and factorial MUT (FMUT) analyses.92, 93 First, ERP differences among valence conditions were tested using a repeated-measures, two-tailed cluster-based permutation test (MUT and FMUT toolboxes, α = .05). 94 The analysis covered 4–596 ms across 63 electrodes (9,450 total comparisons). Repeated-measures t-tests were conducted on observed data and 10,000 random within-subject permutations. Significant t-scores (uncorrected p ≤ .05) formed clusters based on temporal and spatial proximity (~5 cm). Cluster mass (sum of t-scores) served as the test statistic, with the null distribution estimated from 10,001 permutations (original and shuffled datasets). Nonsignificant clusters (p ≥ .05) were assigned p = 1. This method enhanced sensitivity to distributed effects across time and scalp locations, outperforming traditional mean amplitude analysis of variances (ANOVAs). Scalp topographic plots were generated for the N1 and P3b time windows to visualise statistically significant channels. In the subsequent analysis, data were split between genders and similarly analysed for differences in ERP potentials between female and male participants.
Behavioural Data Analysis
Behavioural data recorded included RT, that is, the time between the presentation of the flanker task and response in milliseconds, as well as ACC, indicating whether the response was correct or not. Analysis was conducted via the Jamovi software (Version 2.4) paired with the Walrus robust statistical methods package. 95 Trials with no reaction (2.11% of total trials) were excluded as they represented missed responses. RT data were trimmed (0.2 proportion) to minimise outlier influence while preserving variance. The descriptive statistics suggested potential gender differences. Thereupon, a robust ANOVA was used to examine patterns in RTs, wherein the observed significant main or interaction effects were subjected to post hoc testing for specific differences between groups. Psi-hat, a robust effect size metric, was reported to quantify the differences between groups. A large effect size would be interpreted if the psi-hat value exceeded 50% of the range of the variable measured. Further, to explore specific comparisons, robust paired t-tests were employed with effect sizes (Cohen’s d) used to provide additional context for the observed differences. Interpretation of Cohen’s d is based on the benchmarks previously suggested. 96 Lastly, given the binary nature of ACC data (correct vs incorrect), a binomial logistic regression model was used to examine the influence of predictor variables (Gender, Valence, ‘Congruence × Difficulty’) on task ACC. Interaction terms were included to explore potential moderating effects between predictors.
Results
Identified ERP Components: N1 and P3b
A one-factor, three-level FMUT analysis found no significant ERP differences after correction, suggesting that valence did not elicit measurable neural responses. However, following the nonsignificant omnibus results, pairwise MUT comparisons were considered based on visual inspection of the grand-averaged waveforms (Figure 2).
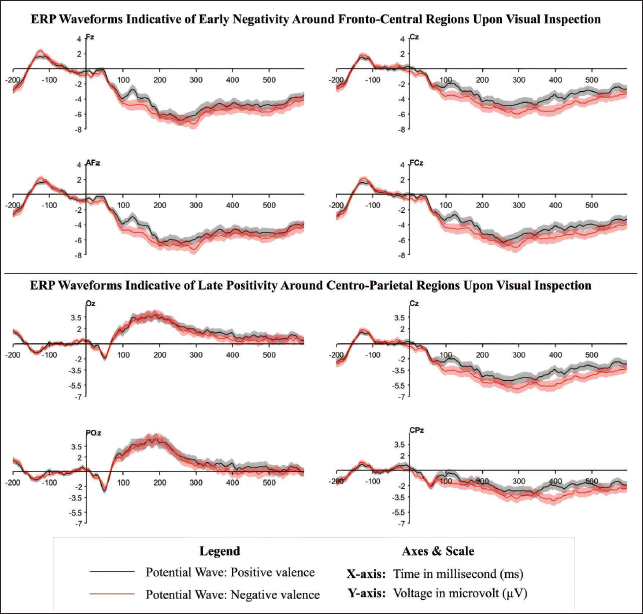
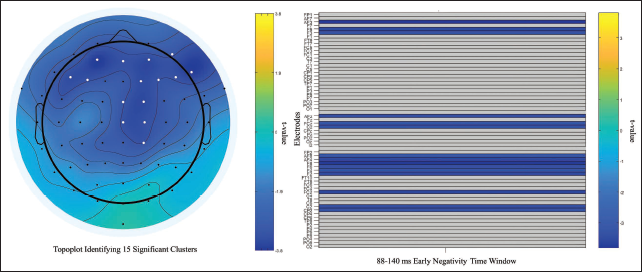
Timepoint-by-timepoint pairwise comparisons from 4 to 596 ms showed no significant ERP differences between positive and negative versus neutral conditions after correction via cluster-based permutation testing (all corrected p values ≥.2067 and ≥.2383, respectively). These findings indicate that neither negative nor positive affective imagery induces unique ERP responses relative to neutral imagery. However, significant ERP differences were observed between positive and negative conditions. Specifically, a cluster was identified at 396 ms after stimulus, localised over CP1 and Pz electrodes, with corrected p values ranging from .0461 to .0375. Consequent window-specific temporal analyses revealed that ERP activity diverged across two time windows. During the first identified time window (88–140 ms), significant negativity was observed at electrodes including F3, CP2, Cz and FC2 (15 significant clusters, corrected p = .0064–.0476, t ≥ −3.1736). The associated raster plot depicted consistent patterns of early ERP discrimination and heightened significant negativity to negative valence, while the topographic maps illustrated a predominantly fronto-central distribution of activity (Figure 3).
Scalp Topographic Plot of Identified Early Negativity Window (88–140 ms) Depicting 15 Significant Negative Clusters (White Dots) in Fronto-central Regions with Adjusted t Values and MUT Raster Plot for the Same Effect with Adjusted t Scores. Significant Clusters Are Identified When Testing the Null Hypothesis That Negative Minus Positive Valenced Potential Waves Have a Mean of 0 µV.
In the second window (352–412 ms), similar significant ERP differences were detected at electrodes such as C3, CP1, Pz and Cz (11 significant clusters, corrected p = .002–.0392, t ≥ −3.2309). The raster plot for this window illustrated a significantly enhanced positivity for the positive valence, and the topographic maps demonstrated a shift towards more posterior electrode sites, suggesting dissociation in the centro-parietal regions (Figure 4).
Scalp Topographic Plot of Identified Late Positivity Window (352–412 ms) Depicting 11 Significant Negative Clusters (White Dots) in Centro-parietal Regions with Adjusted t Values and MUT Raster Plot for the Same Effect with Adjusted t Scores. Significant Clusters Are Identified When Testing the Null Hypothesis That Negative Minus Positive Valenced Potential Waves Have a Mean of 0 µV.

Gender-specific ERP Modulations
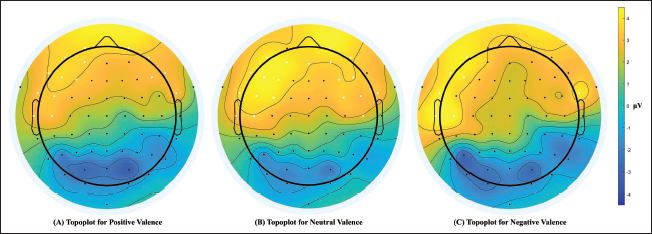
A cluster-based mass-univariate permutation test revealed significant gender-specific differences in ERP responses in all valence conditions. In the positive condition, significant and sustained gender differences emerged from 124 to 560 ms. Specifically, positive clusters were detected at electrodes FP1, F3, F7, FC5 and T7 (corrected p values: .0028–.0448). Females thus exhibited a more pronounced positive potential wave, particularly over higher left frontal and temporal areas (Figure 5A). For the neutral condition, uninterrupted gender differences appeared within the 192–560 ms window, with significant positive clusters at electrodes F3, F7, FC5 and T7 (corrected p values: .0046–.041). Females herein displayed an enhanced symmetrical fronto-temporal positivity pattern of potential waves, similar to the positive condition (Figure 5B). Lastly, in the negative condition, continual significant differences were observed between 192 and 560 ms, with positive clusters (corrected p values: < .05) located over electrodes such as FP2, FC6 and FT8. Topographic plots, therefore, demonstrated a more pronounced positive potential in females for negative stimuli, when compared to males, localised to left lateral frontal regions (Figure 5C).
Scalp Topographic Plots Depicting Gender Differences in ERP Amplitudes (in µV), with Significant Clusters (White Dots) Indicating Regions Where Females Showed Significantly Greater Positive Potentials Across the Three Valence Conditions. (A) Left Frontal and Temporal Activations in Positive Valenced Trials; (B) Symmetrical Fronto-Temporal Activations in Neutral Valenced Trials; (C) Left Lateral Frontal Activations in Negative Valenced Trials. Significant Clusters Are Identified When Testing the Null Hypothesis That the Female Group Minus the Male Group Potential Waves Have a Mean of 0 µV.
However, all pairwise comparisons between valence conditions were nonsignificant (all corrected p ≥ .341), suggesting that gender-specific neural processing differences may not be robust when contrasting between valence categories, as opposed to within individual valence conditions.
Behavioural Performance Outcomes
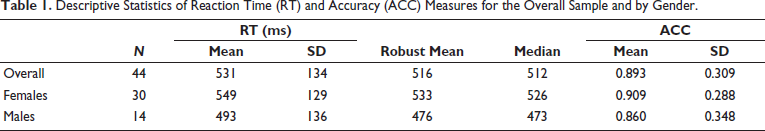
The mean RT for completed trials was 531 ms (±134 ms), with a median of 512 ms. ACC was high, with participants responding correctly in over 89% of trials. Descriptive statistics indicated gender differences in RT, with females having both a slower mean and median, but interestingly showing slightly higher ACC compared to males (Table 1).
Descriptive Statistics of Reaction Time (RT) and Accuracy (ACC) Measures for the Overall Sample and by Gender.
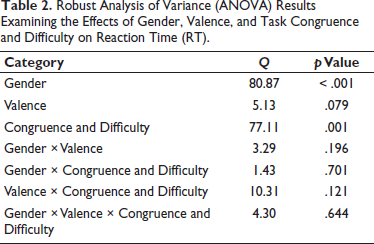
A robust ANOVA revealed significant main effects for Gender (Q = 80.87, p < .001) and ‘Congruence × Difficulty’ (Q = 77.11, p = .001), indicating that RTs varied between genders and across flanker task conditions. However, the main effect of valence was not significant, suggesting that emotional stimuli did not independently influence RTs. No significant interactions were observed between the factors, indicating that the effects of gender and flanker task difficulty on RTs are largely independent of valence and of one another (Table 2).
Robust Analysis of Variance (ANOVA) Results Examining the Effects of Gender, Valence, and Task Congruence and Difficulty on Reaction Time (RT).
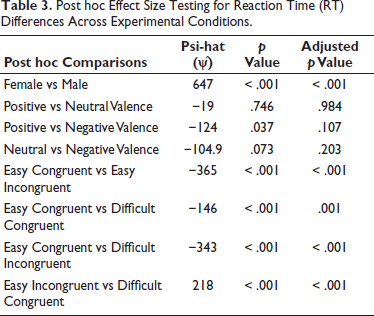
Post hoc tests revealed that females had significantly slower RTs than males, with a large effect size. Additionally, expected significant differences in RTs were observed between all pairs of ‘Congruence × Difficulty’ conditions, although consideration of flanker-based conditions is outside the scope of the current article. However, even in this inspection, RTs did not appear to be significantly affected by valence. A small difference was seen between negative and positive stimuli, though this result was not significant after adjustment (Table 3).
Post hoc Effect Size Testing for Reaction Time (RT) Differences Across Experimental Conditions.
Pairwise t-tests confirmed that females had significantly slower RTs than males. However, the small-to-medium effect sizes suggest that these differences, while statistically significant, were modest (Table 4).
Pairwise t-tests for the Reaction Time (RT) Differences Between Male and Female Adolescents.
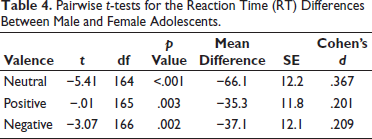
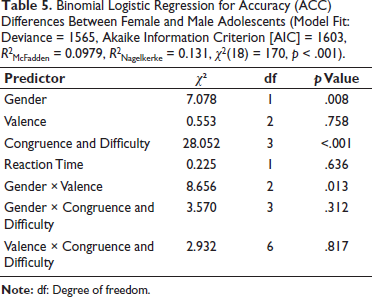
Lastly, a binomial logistic regression was used to analyse ACC data, posteriori satisfying necessary assumptions. Significant predictors included gender, wherein females showed higher odds of ACC, and ‘Congruence × Difficulty’, indicating again that the flanker-based conditions substantially affected response ACC (Table 5). Again, valence did not significantly predict ACC, suggesting that the emotional content of the stimuli had minimal impact on participants’ ability to respond correctly.
Binomial Logistic Regression for Accuracy (ACC) Differences Between Female and Male Adolescents (Model Fit: Deviance = 1565, Akaike Information Criterion [AIC] = 1603, R2McFadden = 0.0979, R2Nagelkerke = 0.131, χ2(18) = 170, p < .001).
Discussion
The present study investigated how a flanker-based cognitive interruption modulated neurobehavioural responses to emotional valence in young adolescents aged 13–14 years. This investigation involved both ERPs and behavioural measures—via an affect-primed flanker task—with stringent criteria for inclusion and data quality to support its validity. Pairwise comparisons between positive and negative valence trials, utilising cluster-based permutation testing, identified significant differences and two ERP components.
The first was consistent with the N1 component (88–140 ms). This component, widely regarded as an index of early attentional allocation, was most pronounced over fronto-central electrode sites. Notably, negative stimuli elicited stronger N1 amplitudes than positive stimuli. Such early neural response dissociation aligned with literature suggesting that emotional stimuli prioritise attentional resources during initial cognitive processing stages.40–42 Similar results have been echoed, demonstrating that emotionally taxing stimuli prioritise attentional resources during initial cognitive processing stages.31, 34 The N1’s modulation by emotional valence thus highlighted the salience-driven early allocation of attention. Negative emotional stimuli, often associated with threat-capture mechanisms,13, 14 appear to automatically recruit additional attentional resources compared to pleasant stimuli, possibly reflecting an evolutionary mechanism for prioritising threat processing.2, 17
The second stage of neural differences between the negatively and positively valenced trials demonstrated a significant late ERP dissociation (352–412 ms), localised over centro-parietal regions. This aligned with the P3b component, 97 widely associated with higher-order cognitive processes, including attentional allocation, working memory updating and stimulus evaluation.51, 98 The observed larger P3b amplitudes for positive compared to negative stimuli are consistent with theoretical models emphasising the role of emotional valence in attentional and cognitive processing. The location of significant clusters highlighted the involvement of centro-parietal regions in higher-order evaluative processing. 47 In particular, the broaden-and-build theory of positive emotions suggests that positive emotional states broaden the attentional focus and facilitate cognitive flexibility, indicating enhanced processing and integration of emotionally rewarding information. 10 The stronger engagement with positive stimuli also aligns with reward sensitivity, which suggests that the brain prioritises the processing of rewarding stimuli to guide behaviour and decision-making.99, 100 Collectively, these findings suggested that positive emotional stimuli may be acting as motivational cues, facilitating a greater P3b activation during this evaluative stage.
Notably, the absence of an expected affective-LPP component may reflect the limited capacity of the cognitive system to engage in both emotional processing and cognitive responding simultaneously.101, 102 The collapse of the LPP component implied that the cognitive demands of the flanker task overwhelmed the system’s ability to sustain attention to emotional stimuli, a phenomenon that highlighted the dynamic nature of emotion–cognition interactions. This suggested that in environments where emotional and cognitive demands are both high, emotional processing may be deprioritised. These results aligned with dual-task theories and the limited-capacity framework, which suggested that when cognitive demands are high, attentional resources are redirected from emotional processing to the cognitive task at hand.68, 103
Overall, the findings elucidated the distinct temporal dynamics of emotional processing in a cognitively demanding environment, with the N1 reflecting early automatic attentional capture and the P3b indexing later cognitive evaluation and resource allocation. Moreover, the temporal dissociations underscored the utility of ERPs in disentangling the multifaceted processes underlying emotion–cognition interactions. 24
Furthermore, our analyses prompted an exploration of gender-based differences in cognitive processing strategies. Females exhibited significantly enhanced positive ERP potentials compared to males across all valence conditions, particularly around fronto-temporal regions. For female adolescents, positive valence trials were associated with a more positive potential in left fronto-temporal areas, while the neutral valence trials elicited a higher, fronto-temporally symmetric positive potential. Negative valence trials were also associated with a greater positive potential for females; however, this was limited to the left lateral frontal regions. Notably, positive valence gender differences were elicited earlier than negative and neutral valences. These patterns suggest a plausible heightened sensitivity to emotional content in females, potentially reflecting greater recruitment of executive and emotional processing networks, consistent with theories proposing that females engage frontal regions more extensively in emotion-related tasks.58, 104, 105 The observed enhanced, sustained omni-valent positivity in female adolescents under attentional demands, despite no pairwise valence differences, might suggest a greater affective sensitivity in both attentional and evaluative information processing among females.106–108
Behaviourally, females demonstrated significantly slower RTs than males; however, the effect of emotional valence was not significant. This indicated that the observed gender differences in response speeds may be independent of emotional content and instead are indicative of a more latent perceptual strategy. Despite slower RTs, females achieved significantly higher ACC rates than males. This trade-off between speed and precision suggests a distinct cognitive strategy employed by females, potentially prioritising ACC over response.63, 64 Overall, the behavioural findings, coupled with the observed enhanced frontal positivity in females, might imply that females allocate greater cognitive and neural resources towards processing emotional stimuli. Consequently, they may adopt a more deliberate and resource-intensive approach compared to males, who might rely on faster, more automatic processing mechanisms.63, 64, 67
Conclusion
To conclude, the study demonstrated that affective processing in young adolescents is characterised by distinct modulations of ERP components under heightened attentional load. The early ERP component (N1) revealed that negative emotional stimuli rapidly capture attention, whereas the later component (P3b) suggests that positive stimuli facilitate enhanced cognitive evaluation. Furthermore, significant gender-specific differences were observed, indicating that attentional-evaluatory processes were differentially affected by biological sex, specifically concerning individual valences, while not impacting any between-valence differences.
Future studies should replicate these findings in larger, more diverse samples. Specifically, the current sample was skewed towards females, which may limit the generalisability of the observed gender differences; future research may employ purposive sampling to balance gender distribution. Moreover, this study focused exclusively on young adolescents (aged 13–14); however, owing to the rapid developmental changes during adolescence, caution must be exercised in generalising these findings. Extending the age range in future studies would allow for examination of developmental variability across early, middle and late adolescence. Further exploration of neurobehavioural responses to flanker task perturbations is needed, along with analyses such as time–frequency, synchrony and source estimation.
Footnotes
Acknowledgements
The authors thank the UX Lab of the Indian Institute of Technology Delhi as well as all participants and their guardians for volunteering their time for the experiment. In addition, the authors express gratitude to the research intern, Ms. Manvi Jain, for assistance with experimental design and data collection.
Authors’ Contribution
MB: Overall supervision of the study, interpretation of data, study conceptualisation, study conduction, analysis of data, drafting of manuscript and approval of final manuscript.
HJ: Interpretation of data and drafting of manuscript.
PS: Recruitment of participants and study conceptualisation.
GS: Drafting of manuscript and approval of final manuscript.
DS: Approval of final manuscript.
JK: Overall supervision of the study.
Data Availability Statement
The datasets produced or analysed in this study are available from the corresponding author (MB) upon reasonable request.
Statement of Ethics
The Institute Ethics Committee (IEC) of the Indian Institute of Technology Delhi approved the research (Proposal No. P021/P0101), which followed the Indian Council of Medical Research (ICMR) ethical guidelines.
Declaration of Conflicting Interests
The authors declared no potential conflicts of interest with respect to the research, authorship and/or publication of this article.
Funding
The authors received no financial support for the research, authorship and/or publication of this article.
ICMJE Statement
The present work adhered to the manuscript guidelines of the International Committee of Medical Journal Editors (ICMJE).
Informed Consent
Adolescents and their guardians gave informed consent before participating in the research, ensuring they understood its aims, methods, potential hazards and outcomes.