
Abstract
The adaptive features of cognitive mechanisms, the features that make them fit for purpose, have traditionally been explained by nature and nurture. In the last decade, evidence has emerged that distinctively human cognitive mechanisms are also, and predominantly, shaped by culture. Like physical technology, human cognitive mechanisms are inherited via social interaction and made fit for purpose by culture evolution. This article surveys evidence from developmental psychology, comparative psychology, and cognitive neuroscience indicating that imitation, mentalizing, and language are “cognitive gadgets” shaped predominantly by cultural evolution. This evidence does not imply that the minds of newborn babies are blank slates. Rather, it implies that genetic evolution has made subtle changes to the human mind, allowing us to construct cognitive gadgets in the course of childhood through cultural learning.
Keywords
What makes psychological mechanisms well suited for their designated roles (i.e., fit for purpose)? Why is it that visual systems can see, learning mechanisms can learn, and reasoning processes can reason? They are all complex products of nature and nurture, but who or what designs the features that enable cognitive processes to do their jobs?
In many cases, genetic evolution (nature) leads the design team. The neonatal face bias—the tendency of newborn babies to orient to faces (Johnson, 2005)—has been honed by natural selection over biological generations. Variant systems were genetically inherited, and through differential reproduction, those that were better at promoting early attention to faces proliferated, whereas the others died out. Learning (nurture) plays a relatively minor role in the development of the neonatal face bias, but in other cases, it is the dominant force. For example, the development of configural face processing—the capacity to recognize faces as wholes rather than using distinctive features—is relatively unconstrained by genetically inherited information and instead depends on experience of a large number of faces (Le Grand, Mondloch, Maurer, & Brent, 2004).
For some distinctively human cognitive mechanisms—absent in other animals or present there only in nascent form—cultural evolution (culture) is a third member of the design team, alongside nature and nurture. Recent evidence, reviewed in this article, suggests that many distinctively human cognitive mechanisms, including imitation, mentalizing (or “theory of mind”), and language, have been shaped by a cultural selection process analogous to genetic evolution. In this cultural evolutionary process, variants arise in individual development, rather than by genetic mutation, and are culturally learned; they are inherited via social interaction rather than DNA. “Good” variants are culturally learned by more agents, but this need not be because the teachers or the learners understand what makes them “good” (Heyes, 2018).
Cultural Evolution of Cognitive Mechanisms
It has long been recognized that behavior and technology, rather than cognitive mechanisms, can be made fit for purpose by cultural evolution (Boyd & Richerson, 1985). Cultural selection has produced the multistage behavioral process—involving scraping, grating, washing, and boiling—used by people in West Africa to remove cyanide from cassava, making it safe and nutritious to eat (Henrich, 2015). Similarly, the design of canoes has been shaped by cultural selection. When people wanted to make a new canoe, they copied the design of an old one. The old canoes available for copying were the ones that had not sunk or fallen apart—the canoes that were relatively good at doing their jobs. Therefore, without needing to understand why some canoes are better than others, people copied good canoes more often than less-good canoes, and the design of these simple boats improved incrementally as a result (Rogers & Ehrlich, 2008).
Now it is becoming clear that not just tangible technology, such as canoes, but also mental technology, cognitive mechanisms, can be shaped by cultural selection (see Fig. 1). Some distinctively human psychological mechanisms are not “cognitive instincts” (Pinker, 1994), shaped primarily by genetic evolution, but “cognitive gadgets,” made fit for purpose primarily by cultural evolution (Heyes, 2018).
Cognitive gadgets. Like simple pieces of tangible technology, such as canoes, distinctively human cognitive mechanisms can be made fit for purpose by cultural evolution. These cognitive gadgets are constructed in the course of childhood development through social interaction.
As we will see in the case studies that follow, evidence that a cognitive mechanism is a gadget rather than an instinct typically comes from research on its development—from studies showing that a way of processing information is acquired from other people through social interaction. The development of cognitive instincts is guided by genetically inherited, domain-specific information (e.g., innate grammar) and noncultural forms of learning (known as “individual learning” and “social learning”). In contrast, the development of cognitive gadgets depends on genetically inherited, domain-general information (e.g., enabling sensory, motor, and all-purpose learning capacities) and on a specific kind of social learning: cultural learning. Children are not merely influenced by their social environments; they learn from other people. In cultural learning, what the learner learns depends on what the model or teacher knows. When cognitive gadgets are culturally learned, input from the sender enables the receiver’s cognitive system to reconstruct the mental software that generated the sender’s modeling or teaching behavior. Thus, the information that builds a cognitive gadget from old parts (i.e., psychological processes shared with other animals) comes from other people who have already acquired the mechanism. It is inherited like money rather than like eye color, through cultural learning rather than DNA (Heyes, 2019).
Imitation
We are “Homo imitans” (Meltzoff, 1988), better able than any other species to copy the topography of body movements—the way in which parts of the body move relative to one another. We use our prodigious capacity for imitation to acquire the facial expressions, bodily gestures, and ritualistic movements (e.g., dance and sports) that promote cooperation with members of our own social group and act as shibboleths, setting us apart from other groups.
Since the 1970s, it has been widely believed that imitation is a cognitive instinct made possible by a powerful, genetically inherited mechanism that relates the “felt but unseen movements of the self with the seen but unfelt movements of the other” (Meltzoff & Moore, 1997, p. 179). In the last 20 years, this consensus has broken down for two reasons. First, the cognitive-instinct theory of imitation was based on experiments suggesting that human newborns can imitate a range of facial expressions, but subsequent studies have failed to replicate these experiments. For example, Oostenbroek et al. (2016) conducted a study of more than 100 neonates using the cross-target procedure introduced by Meltzoff and Moore (1977), which measures the frequency with which infants produce a gesture after observing the same gesture and alternative gestures. This study failed to find evidence of imitation of any of the nine gestures tested. Second, evidence is mounting that the capacity to imitate depends on a vocabulary of learned sensorimotor associations (Catmur & Heyes, 2019; Catmur, Walsh, & Heyes, 2009). Each of these associations links a visual image of an action with a representation of how it feels to perform the action. They are forged by self-observation—for example, when babies watch their own hands in motion—and, crucially, through social interactions in which children engage in synchronous activities (e.g., playing pat-a-cake) or are imitated by adults. Evidence supporting this cognitive gadget, or “associative-sequence-learning” model of imitation, comes from behavioral and neurophysiological experiments showing that, even in adults, the propensity to imitate is highly plastic. A brief period of incongruent sensorimotor experience, in which participants observe one action while performing another, is sufficient to block or reverse an automatic tendency to imitate the trained movements (Catmur et al., 2009). Convergent evidence comes from naturalistic studies of mother–infant interaction. In a longitudinal study in which spontaneous interactions within mother–infant dyads were observed monthly at home from 1 to 10 months, Markodimitraki and Kalpidou (2019) found that the frequency with which infants initiated imitation of sounds and body movements was correlated with that of their mothers. Mothers who initiate more imitative interactions have infants who do the same, and the effect is action specific. Maternal imitation of facial movements, recorded during spontaneous interaction, is positively correlated with infant imitation of face but not hand movements, suggesting that mother–infant interaction supports imitation by enabling infants to learn action-specific sensorimotor associations (Fig. 2; de Klerk, Lamy-Yang, & Southgate, 2019).
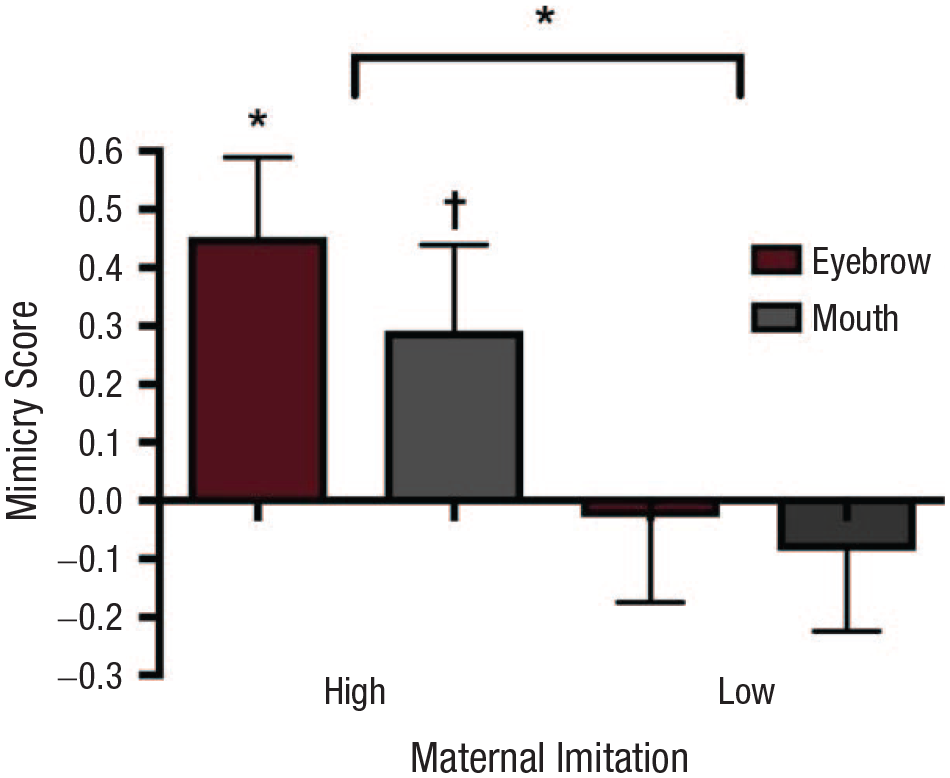
Maternal imitation promotes the development of infant imitation. The graph shows 4-month-olds’ mean mimicry score (activation over the corresponding muscle region minus activation over the noncorresponding muscle region) based on electromyographic recordings of their mouth and eyebrow muscles, separately for infants who are imitated frequently by their mothers (high group) and infants who are imitated less often (low group). The asterisk above the bracket indicates a significant difference between groups, and the symbols above the data bars indicate marginally significant and significant differences from zero (†p < .10, *p < .05). Error bars indicate +1 SEM. Figure adapted from de Klerk, Lamy-Yang, and Southgate (2019).
Mentalizing
Mentalizing (also known as theory of mind and mind reading; Wellman, 2018) occurs when we ascribe mental states, such as beliefs and desires, to ourselves and to others. In cognitive science, behavior is typically explained with reference to neural or computational mechanisms, but in everyday life, people in many contemporary cultures explain, predict, and justify behavior by appealing to what agents think and want. “She shoved the old man because she thought he was going to be hit by a car.” “She raised her hand because she wanted to bid for the painting.” As these examples illustrate, mentalizing underwrites our moral and legal obligations, and it is associated with prosocial behavior in childhood (Imuta, Henry, Slaughter, Selcuk, & Ruffman, 2016).
Increasingly, the development of mentalizing looks like the development of print reading or literacy, a skill that we know is not “in our genes” because scripts have been around for only 5,000 to 6,000 years—not enough time for us to have evolved a reading instinct (Heyes & Frith, 2014). For example, parents and other adults scaffold the development of print reading by exposing children to easy to read words, such as “cat,” before hard to read words, such as “yacht.” Similarly, when talking to their infants, mothers mention easy to read mental states, such as desires and emotions, before hard to read mental states, such as beliefs and knowledge, and the frequency of developmentally appropriate references to mental states predicts the children’s later development of mentalizing skill (Taumoepeau & Ruffman, 2008). Instruction is also important in the development of both print reading and mentalizing. Just as children are instructed in rules of pronunciation (e.g., told what “-tion” sounds like when read aloud), in conversation with their children, mothers make “causal-explanatory” statements that specify relationships among situations, behavior, and mental states (e.g., “He is smiling because he is happy. He is happy because he is playing with the puppy”), and the frequency of these causal-explanatory statements predicts individual differences and cultural variation in the development of mentalizing (Slaughter & Peterson, 2012).
These findings, and many others (e.g., Pyers & Senghas, 2009), suggest that people learn to mentalize through social interaction with members of their social group who are already skilled mentalizers. In principle, this cultural learning could be guided by a genetically inherited cognitive instinct for mentalizing. However, parallels with print reading cast doubt on what used to be regarded as evidence for the innateness of mentalizing. For example, print reading also shows neural specialization—it depends heavily on certain parts of the brain—indicating that cognitive gadgets are just as likely as cognitive instincts to be specialized at the neural level. Similarly, print reading, like mentalizing, is subject to developmental disorders. People with dyslexia have difficulty with print reading, and people with autism spectrum disorder have difficulty with mentalizing (Heyes & Frith, 2014).
Experiments using looking-time measures suggest that young infants are capable of mentalizing when they have had relatively little opportunity for cultural learning (Onishi & Baillargeon, 2005). However, the results of these studies are proving difficult to replicate (Baillargeon, Buttelmann, & Southgate, 2018; Dörrenberg, Rakoczy, & Liszkowski, 2018; Poulin-Dubois et al., 2018), and even if the looking-time effects are reliable, they can be explained by domain-general mechanisms, such as attention and working memory, that is, without assuming that human infants are born with a propensity to mentalize. Domain-general mechanisms cobble together cognitive gadgets using input from cultural learning. However, many human capacities, including behavior prediction in infancy, depend on “neat” domain-general mechanisms, not on the domain-specific, distinctively human cognitive gadgets built by those mechanisms (Heyes, 2014).
Language
Even language, the foundational cognitive instinct (Pinker, 1994), is beginning to look like a cognitive gadget, shaped predominantly by cultural rather than genetic evolution (Christiansen, Chater, & Culicover, 2016). Starting in the 1990s, evolutionary psychologists cited research on developmental disorders, neural specialization, and the genetic bases of language in support of Chomsky’s proposal that the development of language is grounded in innate knowledge of grammatical rules. However, much of this evidence has now been overturned. For example, specific-language impairment, a developmental disorder once thought to affect language acquisition alone, turns out not to be specific to language. Children diagnosed with the condition struggle to learn sequences of lights and objects, not just the order of words (Hsu & Bishop, 2014). Similarly, there does not appear to be a language center in the brain. Broca’s area in the left hemisphere has long been regarded as the seat of language, but recent research suggests that language is scattered throughout the cortex. Meta-analysis of brain-imaging studies shows that Broca’s area is in fact more likely to be active when people are performing tasks that do not involve language than when they are reading, listening to, and producing words (Poldrack, 2006). Furthermore, the forkhead box protein P2 (FOXP2) gene, once regarded as a language gene, has been implicated in sequence learning more generally. Transgenic mice implanted with the human version of FOXP2 are better than their siblings at finding their way around a maze (Schreiweis et al., 2014).
Alongside these developments, there is positive evidence that language can be acquired, by computers and by children, via domain-general sequence-learning processes without built-in language-specific constraints. For example, computers can learn grammatical rules without any preprogrammed grammatical knowledge (Christiansen & MacDonald, 2009). Experiments examining individual differences in typically developing adults and children indicate that they use the same sequence-learning processes to learn artificial and “real” linguistic grammars (Kidd & Arciuli, 2016). Research with nonhuman animals has confirmed that domain-general sequence-learning capacity has increased in the hominin lineage and provided a plausible model of how this change has been implemented in the primate brain (Bornkessel-Schlesewsky, Schlesewsky, Small, & Rauschecker, 2015).
Most interesting from an historical perspective, research now indicates that infants and children are frequently corrected by adults when they make grammatical errors and that these social signals are used in language learning (Taumoepeau, 2016). This evidence challenges Chomsky’s “poverty-of-the-stimulus” argument (Chomsky, 1975) for the innateness of language and, like research on sequence learning, confirms novel predictions of the hypothesis that language has been made fit for purpose primarily by cultural evolution.
Implications
The evidence that imitation, mentalizing, and language are cognitive gadgets does not imply that the mind of a newborn human baby is a blank slate. In common with other animals, humans genetically inherit not only perceptual and motivational processes but also a wide range of domain-general cognitive mechanisms. They are domain general in that they process information of different kinds—for example, about other agents and about the inanimate world—using the same set of computations. They mediate selective attention, memory, learning, and executive functions, such as response inhibition and task switching. Most of these domain-general processes are widely distributed across the animal kingdom and are found in birds and small mammals as well as primates. However, there is evidence that, in the human lineage, they have been tweaked by genetic evolution in ways that allow us to absorb information from other agents quickly and easily.
For example, humans genetically inherit a face bias, a tendency to look longer at faces than at other stimuli, which locks our attention on other agents from birth (Johnson, 2005). Similarly, adult humans are friendlier—more tolerant of social contact and more strongly motivated by social rewards—than adults of many other primate species (Cieri, Churchill, Franciscus, Tan, & Hare, 2014), which gives children the opportunity to learn from a host of other agents, not only their mothers. Furthermore, compared with other animals, including nonhuman primates, humans have expanded capacities for learning and memory, allowing us to absorb the information supplied by other agents in large volumes and at unprecedented speed (Heyes, 2018). Humans genetically inherit psychological features that are only subtly different from those of other animals, but they allow us to upload from our cultural environment not only a wealth of knowledge and technical skills but also new pieces of mental technology—cognitive gadgets.
I have focused on imitation, mentalizing, and language because, to date, these processes have been examined most closely from a cultural evolutionary perspective. There is also evidence that social-learning strategies (Heyes, 2018), moral reasoning (Heyes, 2019), and metacognition are culturally learned (Heyes, Bang, Shea, Frith, & Fleming, 2020). But more research is needed—in developmental, comparative, and cross-cultural psychology and in cognitive neuroscience—to find out exactly what cultural learning contributes to the development of these and other distinctively human cognitive mechanisms. We need to identify the contributions of nature, nurture, and culture. The evidence surveyed in this article suggests that we can no longer assume that adaptive features of distinctively human cognitive processes are due to genetic evolution (nature), to learning by direct interaction with the environment during an individual’s development (nurture), or to a combination of both factors. They could be, and often are, inherited through social interaction and made fit for purpose by cultural evolution.
Recommended Reading
Christiansen, M. H., Chater, N., & Culicover, P. W. (2016). (See References). An accessible overview of the history and current status of debate about the evolutionary origins of language.
Conway, J. R., Coll, M.-P., Cuve, H. C., Koletsi, S., Bronitt, N., Catmur, C., & Bird, G. (2020). Understanding how minds vary relates to skill in inferring mental states, personality, and intelligence. Journal of Experimental Psychology: General, 149, 1032–1047. doi:10.1037/xge0000704. Evidence that successful mentalizing involves much more than the ascription of thoughts and feelings.
de Klerk, C. C., Lamy-Yang, I., & Southgate, V. (2019). (See References). Compelling evidence from 4-month-old infants that imitation is culturally inherited.
Heyes, C. M. (2019). Précis of Cognitive Gadgets: The Cultural Evolution of Thinking. Behavioral and Brain Sciences, 42, Article e169. doi:10.1017/S0140525X18002145. A summary of Heyes’s (2018) book, published with commentary articles by psychologists, neuroscientists, philosophers, and evolutionary biologists and a response from the author.
Oostenbroek, J., Suddendorf, T., Nielsen, M., Redshaw, J., Kennedy-Constantini, S., Davis, J., . . . Slaughter, V. (2016). (See References). A rigorous, large-scale study that did not find any evidence of imitation in human newborns.
Footnotes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.