
Abstract
To address the lack of effective treatments for germinal matrix hemorrhage-intraventricular hemorrhage (GMH-IVH), a devastating condition in preterm infants with poor prognosis, this study investigated the neuroprotective potential and underlying mechanism of human umbilical cord-derived mesenchymal stem cells (HUCMSCs). Using a collagenase VII-S-induced GMH neonatal rat model and lipopolysaccharide-stimulated BV2 microglial cells, in vivo and in vitro experiments were performed. In vivo results showed that intraventricular HUCMSCs administration alleviated cerebral injury (reduced ventricular enlargement and cortical damage) and improved motor, memory, and cognitive functions in GMH rats. Both HUCMSCs and MSC-conditioned medium (MSC-CM) modulated microglial polarization toward the pro-resolving phenotype, suppressed pro-inflammatory polarization, and decreased TNF-α and IL-6 expression. Mechanistically, MSCs downregulated ROCK2 protein levels; the ROCK agonist PA abrogated MSC-CM’s anti-inflammatory effects, while the ROCK inhibitor Y27632 mimicked MSC-CM’s pro-M2 polarization. Collectively, this study demonstrates that HUCMSCs exert neuroprotection against GMH by suppressing ROCK2 to regulate microglial polarization, thereby mitigating neuroinflammation and promoting nerve repair, highlighting MSCs as a promising therapeutic strategy for GMH-IVH.
Introduction
Germinal matrix hemorrhage (GMH) is a form of neonatal cerebral hemorrhage that originates from the germinal matrix within the subventricular ependymal region. It primarily affected preterm infants born before 32 weeks of gestation or weighing less than 1500 g, particularly those born very prematurely before 28 weeks of gestation or with extremely low birth weights below 1000 g 1 . The prognosis for severe intraventricular hemorrhage (IVH) complicated by GMH was unfavorable, and effective treatment strategies remained lacking in clinical practice. Therefore, the development of treatment for GMH-IVH was an urgent priority.
Cerebral injury following GMH comprises primary and secondary injury, with the latter being the primary determinant of prognosis2,3. Neuroinflammation stood out as a pivotal contributor to secondary cerebral injury 4 . Microglial cells, as the foremost glial cells that reacted to brain trauma, possessed the capacity to generate an array of detrimental cytokines when in an overactivated state, thus fueling inflammation. Depending on the microenvironment, microglia can exhibit three distinct phenotypes: homeostatic microglia, pro-inflammatory microglia, and pro-resolving microglia 5 . Targeted therapeutic strategies post GMH-IVH focused on restraining pro-inflammatory microglia activation and fostering pro-inflammatory to pro-resolving transition during the acute phase6,7.
Stem cells therapy had emerged as a predominant approach in the field of therapeutic research for treating brain injuries due to its effectiveness in tissue and organ repair. Recent studies had demonstrated that human umbilical cord–derived mesenchymal stem cells (HUCMSCs) possess self-renewal capacity, high proliferation rate, multi-lineage differentiation potential, and immune-modulating properties, making them a promising alternative source of mesenchymal stem cells (MSCs) for both basic research and clinical applications 8 . The therapeutic mechanism of stem cells transplantation for central nervous system injuries involved immunomodulation, secretion of trophic factors, and enhancement of local microcirculation, indicating a comprehensive reparative mechanism9–11. Despite advancements in translating MSC therapy for GMH into clinical practice, variations in efficacy still existed12–14, underscoring the importance of elucidating the precise mechanisms underlying MSCs treatment. This study utilized a D7 neonatal rat GMH model to investigate the impact of intraventricular HUCMSCs injection on microglial polarization phenotype regulation, aiming to uncover the potential mechanisms involved in this process and provide a solid scientific and theoretical foundation for HUCMSC-based GMH treatment.
Materials and methods
HUCMSC preparation and identification
Following written informed consent from all pregnant women, human Wharton’s jelly–derived MSCs were isolated and expanded. This study was approved by the Ethics Committee of the Affiliated Hospital of Guizhou Medical University (approval No. 631, approved on 13 July 2021). Umbilical cords were processed by washing with PBS, removal of blood and vessels, cutting into 1 mm3 fragments, and culturing in α-MEM (Gibco, Gibco, Grand Island, NY, USA) with 10%–15% FBS (Sigma-Aldrich, Gibco, Grand Island, NY, USA), penicillin, and streptomycin. Cells were maintained in a 37°C incubator with 5% CO2, with medium changes every 3 days. After 5–7 days, cells migrated from tissue fragments and proliferated, and after 14 days, cells were harvested using pancreatic enzyme for expansion and characterization. HUCMSCs were identified through flow cytometry analysis of cell surface markers (CD34, CD73, CD90, CD45, and CD11b). Flow cytometry analysis was performed using a Beckman CytoFLEX flow cytometer (Beckman Coulter, Brea, CA, USA). The gating strategy was set as follows: first, a forward scatter (FSC) versus side scatter (SSC) dot plot was used to gate the viable cell population and exclude cell debris; subsequently, single cells were gated using FSC-H vs. FSC-A to eliminate cell aggregates; finally, the expression of surface markers was analyzed in the gated single viable cell population. For flow cytometry analysis, the sample size was set as n = 3 per group, and HUCMSCs were isolated from two independent umbilical cord samples. For each sample, at least 10,000 events were recorded and analyzed using CytExpert software (version 2.4, Beckman Coulter). P4 to P10 HUCMSCs were utilized in subsequent experiments.
Harvest and concentration of MSC-CM
HUCMSCs were cultured in serum-free α-MEM for 24 h to generate MSC-CM. The CM was filtered using a 0.22-μm filter to eliminate cellular debris, and then concentrated threefold in a 10-kD Millipore ultrafiltration tube (Millipore, Billerica, MA, USA) by centrifugation at 5000 rpm for 15 min in a refrigerated high-speed centrifuge. The concentrated supernatant was aliquoted into a centrifugal tube for further applications. The MSC-CM derived from 2 × 106 MSC cells yielded 5 ml of medium.
Labeling of HUCMSCs
Selected P4 or P5 HUCMSCs transduced with green fluorescent protein (GFP) using a replication-incompetent adenovirus carrying GFP. Upon achieving a GFP expression rate of approximately 70%, the cells were harvested for transplantation.
Induction of germinal matrix hemorrhage (GMH)
A total of 132 healthy Sprague-Dawley rat pups, aged 7 days (P7), weighing approximately 15 ± 2 g, were obtained from the Guizhou Medical University Animal Center. The rats were randomly assigned to either the sham or GMH groups, and no rats were excluded from the study due to intraoperative death. All rat pups were housed in temperature-controlled rooms with ad libitum access to food and water. GMH induction was performed on unsexed P7 rats using collagenase VII-S (Sigma-Aldrich, St. Louis, MO, USA) infusion. In brief, the rats weighing between 13 and 17 g were anesthetized with 3%–5% isoflurane (RWD Life Science, Shenzhen, China), and anesthesia was maintained with 1%–2% isoflurane during the surgery. The rat pups were secured in a stereotaxic frame, and the scalp was sterilized. A burr hole (1 mm) was drilled at 1.5 mm right lateral and 1.6 mm rostral relative to the bregma, after which a 10-µl Hamilton syringe filled with 0.3 U/µl collagenase solution was attached to an infusion pump. The needle was inserted 2.7 mm below the dura with the bevel facing the midline. The collagenase VII-S solution [0.3 U in 1 μl volume of TES-Calcium Chloride buffer (TESCA)] was infused at a rate of 1 μl/5 min, and the needle remained in place for 5 min before being removed at a rate of 1 mm/min to prevent leakage. The burr hole was sealed with bone wax, and the incision was sutured closed with 7-0 silk. After the surgery, the rats were placed on a 37°C heating pad to recover from anesthesia, and once fully conscious, they were returned to their dams. The sham surgery followed the same procedure as described above and infusion of 1 μl TESCA. The average duration of the surgery for each animal was approximately 20–30 min.
HUCMSCs intraventricular transplantation
Cell transplantation was conducted 24 h after successful modeling, with HUCMSCs resuscitated 48 h in advance. HUCMSCs from the P4–P10 generation, exhibiting favorable cell morphology and robust proliferation capability, were chosen. The cells were administered to the animals within 1 h. The rats were randomly divided into sham group, vehicle group, and MSCs groups, anesthetized with 3%–5% isoflurane, and 1%–2% maintained under anesthesia on the operating table. Local skin was disinfected using iodophor and alcohol, the fontanel was exposed, and a brain stereotaxer was used to target the same coordinates as the model (coordinates: bregma as the origin, mediolateral (ML) 1.5 mm, anterior-posterior (AP) 1.6 mm, dorsoventral (DV) 2.7 mm), which corresponded to the lateral ventricle-adjacent germinal matrix of the right hemisphere (consistent with the collagenase injection site for modeling). Then, 5 μl of cells (cell count: 5 × 105/5 μl) were injected at a rate of 1 μl/min. Following completion of the injection, the needle was held steady for 5 min, slowly withdrawn to prevent liquid leakage, the small hole sealed with bone wax, and the skin sutured after disinfection. Post-surgery, the rats were placed on a 37°C bed until they were cared for by the mother rats upon awakening. The vehicle group underwent the same procedure but was injected with 5 μl of sterile PBS.
Negative geotaxis test
7 days after HUCMSCs transplantation, pups were subjected to a 45° head-down tilt test to measure the time taken to complete a 180° rotation. Each trial had a maximum duration of 60 s, with three trials conducted per pup per day. The average time to complete the rotation was calculated based on the values obtained from the three trials.
Rotarod tests
Prior to the formal experiment, the neonatal pups in each group underwent a 3-day period of continuous training. The test was carried out 21 days after HUCMSCs transplantation. The rats were positioned on a rotating rod apparatus (ZSDichuang, Guangzhou, China), with the rotational speed increasing from 20 to 60 rpm over a maximum duration of 5 min. Three trials were conducted, during which the time and distance of falls were documented, and the average values were calculated for statistical analysis.
Water maze test
Morris water maze experiments were conducted at 21 days after HUCMSCs transplantation to evaluate cognitive function and memory in animals, following established protocols. The apparatus consisted of a pool (110 cm in diameter) accompanied by a small platform (11 cm in diameter), in which the pups could climb to escape the water. Swim distance and latency were recorded digitally and analyzed using tracking software. In the cruise tests, the rats were manually guided to the platform if they had difficulty in locating it in 60 s. In the spatial probe tests, the platform was removed and the rats’ movement trajectory and residence time in the target quadrant were recorded.
Brain section
The brains were taken directly after euthanasia using isoflurane anesthesia. Using a brain matrix, continuous coronal sections of the brain were taken caudally at the beginning of optic chiasm. The section thickness was 1.5 mm. The slices were placed on a clean plane for observation and photography. Image-J analysis software was used to calculate the area of the obtained images, ventricle volume(mm3) = area of each ventricle (mm2) ×1.5 mm.
In vitro experiment
Concentrated MSC-CM interfered with BV2 cells
The mouse microglia (BV2) cells were obtained from Procell (China, Wuhan) and cultured in BV2 special medium (CM-0493A, Procell) at 37°C with 5% CO2. The cells were seeded at a density of 5 × 104/well in 6-well plates. Four hours after cell adhesion, the group is divided into Control group, Lipopolysaccharide (LPS) group, and LPS+MSC-CM group according to whether LPS (100 ng/ml) and concentrated MSC-CM were added or not, after 24 h, BV2 cells were harvested for Western blotting and immunofluorescence analysis. Furthermore, to confirm the regulatory role of MSCs on microglial polarization via ROCK regulation, a ROCK agonist (PA, 5 μM; MCE) and a ROCK inhibitor (Y27632, 20 μM; MCE) were utilized. The BV2 cells were categorized into Control group, LPS group, LPS+MSC-CM group, LPS+PA group, LPS+PA+MSC-CM group, and LPS+Y27632 group for intervention.
Transwell experiment
The BV2 cells were seeded in the lower compartment at a density of 5 × 104/well in 6-well plates. Four hours after cell adhesion, the group is divided into LPS group and LPS + MSCs group to whether LPS (100 ng/ml) and HUCMSCs seed in the upper compartment (BV2 count:HUCMSCs count = 1:1), after 24 h, BV2 cells were harvested for Western blotting.
Immunofluorescence
Brain tissue samples were fixed in 4% paraformaldehyde overnight. Dehydrated in 30% saccharose, embedded in optimal cutting temperature compound (OCT) and cut into 15-μm-thick slices. The BV2 cells and brain slices underwent permeabilization with 0.05% Triton X-100 (Sigma-Aldrich, St. Louis, MO, USA) for 10 min at room temperature, followed by blocking with 5% bovine serum albumin at 37°C for 1 h, and incubation with the primary antibody overnight at 4°C. Subsequently, the slices were exposed to fluorescence secondary antibody goat anti-mouse FITC (1:100; Biopm, Shanghai, China) or goat anti-rabbit CY3(1:200; Biopm, Shanghai, China) at room temperature for 1 h. Finally, the nuclei were counterstained by DAPI for 10 min at room temperature. Imaging was conducted using a fluorescence microscope and analyzed with Image-J software under consistent acquisition parameters to minimize background interference. Primary antibodies utilized in the study included CD206 (1:50; Santa, USA), iNOS (1:100; Proteintech, Wuhan, China), and Arg-1 (1:100; Proteintech, Wuhan, China).
Nissl’s staining
Twenty-eight days after HUCMSCs transplantation, rats anesthetized with isoflurane underwent intracardial infusion of normal saline and paraformaldehyde, fixed at 4°C with 4% paraformaldehyde for 24 h, then dehydrated, paraffin-embedded, sliced (5 μm), stained with Nissl’s staining kit (ServiceBio, China), and observed the brain structure of each group under a microscope.
Western blot analysis
Brain samples and BV2 cells were homogenized in Phenylmethylsulfonyl fluoride (PMSF) and Radio-Immunoprecipitation Assay (RIPA) buffer (PMSF:RIPA = 1:99, Solarbio, Beijing, China). Following quantification through Bicinchoninic acid (BCA) assay, the proteins were separated on Sodium Dodecyl Sulfate (SDS) gels and transferred to Polyvinylidene Fluoride (PVDF) membranes (Millipore, USA). The membranes were blocked with 5% Bovine Serum Albumin (BSA) for 1 h at room temperature and incubated with primary antibodies overnight at 4°C. Subsequently, peroxidase-conjugated secondary antibodies goat anti-rabbit (1:3000; Proteintech, China) were utilized for 1 h at room temperature. Protein bands were detected by the imaging system (Tanon, China) and quantified by Image-J software. Primary antibodies utilized in the study included β-actin (1:5000; Bioworld, China), GAPDH (1:1000; ServiceBio, China), iNOS (1:4000; Proteintech, China), Arg-1 (1:3000; Proteintech, China), and ROCK2 (1:1000; Affinity, China).
Quantitative real-time PCR analysis
Total RNA was extracted from BV2 cells using an RNA extraction kit (Mei5 Bioservices Co. Ltd, Wuhan, China). Subsequent to reverse transcription into cDNA using a cDNA synthesis kit (Yeasen, Shanghai, China). Quantitative PCR (qPCR) was conducted using synthetic primers and SYBR Green (Yeasen, China) on a CFX96 Detection System. After incubation at 95°C for 5 min, samples were subjected to 40 cycles of 95°C for 10 s and 60°C for 30 s. Normalization of final results was based on the division of target values by a reference value of “1.” The expression of IL-6, TNF-α, and GAPDH were determined by qPCR using the followed primers:
IL-6, forward: TGGAGTTCCGTTTCTACCTGG, reverse: GGTCCTTAGCCACTCCTTCTGT;
TNF-α, forward: CTCTTCAAGGGACAAGGCTG, reverse: CGGACTCCGCAAAGTCTAAG;
GAPDH, forward: GCAAGTTCAATGGCACAG, reverse: GCCAGTAGACTCCACGACA.
Statistical analysis
All data were analyzed using IBM SPSS 22.0 or GraphPad Prism 8 software and expressed as the mean ± standard deviation (SD). All continuous variables were tested for normality and homogeneity of variance prior to group comparisons. For normally distributed data, one-way analysis of variance (ANOVA) was used for multiple group comparisons, followed by Tukey’s post hoc test for pairwise comparisons. For non-normally distributed data, the Kruskal–Wallis test was applied, and Dunn’s test with Bonferroni correction was used for post hoc analysis to control Type I error. Repeated-measures data were analyzed by repeated-measures ANOVA with Greenhouse–Geisser correction when sphericity was violated. A value of P < 0.05 was considered statistically significant.
Results
Isolation and identification of HUCMSCs
HUCMSCs were isolated using the tissue block adherent method. Following 7 days of cultivation, cells migrated out of the tissue blocks, and were passaged after 14 days. Microscopic examination revealed that the cells displayed elongated spindle shapes, grew adherently in clusters, and formed vortex-like structures, consistent with the morphological characteristics of MSCs (Figure 1a). Flow cytometry analysis was performed on purified HUCMSCs, showing expression of CD90 (99.15% ± 0.25%), CD73 (100%), CD45 (0.09% ± 0.017%), CD34 (0.133% ± 0.0058%), and CD11b (0.13% ± 0.02%) (Figure 1b). These findings demonstrated that cells obtained through the tissue block adherent method exhibited positive expression of CD90 and CD73 on the cell surface, and negative expression of CD45, CD34, and CD11b, meeting the international standards of HUCMSCs. This validated the successful isolation of high-purity HUCMSCs using the tissue block adherent method, suitable for subsequent experiments.

Isolation and identification of HUCMSCs. (a) HUCMSCs were isolated using the tissue block adhesion method, and the microscope images were captured at 7, 9, 11, 14 days and P1 after culture. Scale = 100 μm. (b) Detection of cell surface markers for identification: CD90, CD73 were positive, while CD45, CD34, and CD11b were negative. Red is the homologous control group, green is the sample group.
Tracking of GFPHUCMSCs at different time points in vivo
To study the survival time of HUCMSCs transplanted into the ventricle, GFP-labeled HUCMSCs were utilized for tracking. After transplanting HUCMSCs carrying GFP into the rat brain, the brain tissues were collected for frozen sections at 3, 7, 10, 14, and 28 days post-transplantation. The findings demonstrated a progressive decline in green fluorescence intensity within the ventricle over time, by the 28 days post-transplantation, there were a small amount of green fluorescence was observed around the ventricular wall (Figure 2), indicating that the number of transplanted exogenous HUCMSCs gradually decreased with the extension of time, and only a small number of cells survived at 28 days, which also suggest that the main mechanism of HUCMSCs is derived from its secretion, rather than cell replacement.

Trace of transplanted GFPHUCMSCs in vivo. (a) Ventricle level fluorescence imaging at 3 days post-transplantation, with an enlarged image provided in (c). Scale = 200 μm. (b) Hippocampus level fluorescence imaging at 3 days post-transplantation with enlarged images in (d–e). Scale = 200 μm. (f–i) Fluorescence imaging surrounding the ventricle at 7, 10, 14, and 28 days post-transplantation. Scale = 25 μm. Green is GFP, blue is DAPI shows nuclei, red arrow indicates transplanted HUCMSCs cells.
HUCMSCs improved ventricular volume enlargement, cerebral cortex reduction, and neural function following GMH
The GMH model was established successfully through the injection of collagenase VII-S, resulting in visible right ventricle hemorrhage at 24 h and right ventricle enlargement at 14 days (Figure 3a), effectively replicating the pathophysiology of neonatal GMH. Brain tissue samples were collected at 3, 7, and 10 days post-HUCMSCs transplantation. Histological analysis revealed asymmetrical cerebral cortex dimensions in the vehicle group compared with the sham group, with a reduction observed at the right injection site. In contrast, the MSCs group exhibited improved cerebral cortex morphology (Figure 3c), suggesting a beneficial effect of HUCMSCs on post-GMH cerebral cortex injury. The examination of rat brain tissue slices 28 days post-treatment revealed a significant enlargement of the right ventricle and increased ventricle volume in the vehicle group compared with the sham group (P < 0.001). Conversely, the MSCs group displayed a significant reduction in ventricle volume (P < 0.01 vs vehicle group) (Figure 3d, g). Nissl’s staining results also revealed that right ventricle dilation was observed in the vehicle group compared with the sham group, whereas ventricle dilation was significantly reduced in the MSCs group (Figure 3b), suggesting that HUCMSCs can improve the hydrocephalus complicated after GMH. The negative geotaxis test conducted 7 days post-treatment indicated a significantly prolonged latency in the vehicle group compared with the sham group (P < 0.05), whereas the latency in the MSCs group was significantly reduced (P < 0.01 vs vehicle group) (Figure 3h). The results of the rotarod test 21 days post-transplantation showed that compared with the sham group, the distance at drop in the vehicle group was significantly reduced (P < 0.0001), while the distance at drop in the MSCs group was higher than that in the vehicle group (P < 0.05) (Figure 3j), indicating that HUCMSCs enhanced the motor coordination capacity of GMH rats. In the water maze test, the latency of MSCs group was significantly reduced compared with the vehicle group on the third and fourth day of constant speed cruising (third day: P < 0.05, fourth day: P < 0.05). Similarly, in the spatial search test, the latency of the MSCs group was significantly lower than that of the vehicle group (P < 0.05) (Figure 3e, f, i), suggesting that HUCMSCs can enhance the cognitive function and memory performance of GMH rats. These beneficial effects are likely mediated via paracrine or protective mechanisms rather than direct cell replacement (for causes explained later in the article).

Impact of HUCMSCs on mitigating cerebral damage in GMH rats. (a) Comparison of the sham group and the vehicle group at 24 h and 14 days post-modeling, red arrow indicates the right ventricle. Scale = 1 mm. (b) Nissl’s staining of brain tissue 28 days post-transplantation in three groups. Scale = 2 mm. (c) The gross morphology of brain tissue in the three groups at 3, 7, and 10 days post-transplantation, red arrow indicates the cerebral cortex at the right injection site. Scale = 5 mm. (d) Right brain tissue slices 28 days post-transplantation in three groups. (e) Statistical analysis of latency in the cruise trials conducted on three groups. Scale = 1 cm. (f) The spatial probe testing route map in three groups. (g) Statistical analysis of ventricle volume 28 days post-transplantation in the three groups. (h) Statistical analysis of negative geotaxis test latency 7 days post-transplantation in the three groups. (i) Statistical analysis of latency in spatial probe trials in three groups. (j) Statistical analysis of the drop distance in three groups 21 days post-transplantation. Data are expressed as mean ± standard deviation. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001; Compared with the vehicle group, #P < 0.05.
MSCs promoted microglial polarization to pro-resolving state and decreased polarization to pro-inflammatory state in vivo
In order to further investigate the regulatory impact of MSCs on microglial polarization phenotype, tissue immunofluorescence analysis revealed a significant increase in the pro-inflammatory state marker (iNOS) in the vehicle group compared with the sham group (P < 0.001), with no significant alteration in the pro-resolving state marker (CD206) (P > 0.05). In the MSCs group, iNOS fluorescence intensity notably decreased (P < 0.05 vs vehicle group), while CD206 fluorescence intensity significantly increased (P < 0.0001; Figure 4a–d). Similar results were also verified by Western blot experiment, the protein expression level of iNOS in the MSCs group was significantly decreased (P < 0.05 vs vehicle group), while the protein expression level of pro-resolving state marker (Arg1) was significantly increased (P < 0.001; Figure 4e and f). In conclusion, in vivo experiments had shown that MSCs can promote the polarization of microglia in rat brain tissue to pro-resolving state, reduce the polarization to pro-inflammatory state, reduce inflammation, and promote nerve repair, which may be one of the important mechanisms of MSCs playing a protective role in the brain after GMH.

MSCs regulate microglia polarization in vivo. (a) iNOS and DAPI expression in periventricular brain tissues was detected using immunofluorescence. iNOS was shown in red, DAPI in blue for the nuclei, LV for lateral ventricle, PWM for periventricular white matter. Scale = 50 μm. (b) CD206 and DAPI expression in periventricular brain tissues was detected using immunofluorescence. CD206 was shown in green, DAPI in blue for the nuclei. Scale = 50 μm. (c) Statistical analysis of fluorescence expression of iNOS in three groups. (d) Statistical analysis of fluorescence expression of CD206 in three groups. (e) Western blot detection of iNOS and Arg1 protein expression in three groups. (f) Statistical analysis of iNOS protein relative expression in three groups. (g) Statistical analysis of relative expression of Arg1 protein in three groups. Data are expressed as mean ± standard deviation. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
MSC-CM promoted the polarization of BV2 cells toward pro-resolving state and decreased the polarization toward pro-inflammatory state in vitro
The efficacy of MSCs in modulating microglial polarization has been validated through in vivo experiments. To further substantiate this finding, we will conduct additional validation at the in vitro level. Since the beneficial effects of MSCs may be related to their secretory activity, the cells were first treated with concentrated MSC-CM. Immunofluorescence analysis revealed a significant increase in iNOS fluorescence intensity in the LPS group compared with the control group (P < 0.0001), along with a decrease in Arg1 fluorescence intensity (P < 0.001). In contrast to the LPS group, iNOS fluorescence intensity was significantly reduced in the LPS+MSC-CM and MSC-CM groups (LPS+MSC-CM: P < 0.0001; MSC-CM: P < 0.0001), while Arg1 fluorescence intensity was notably upregulated (LPS+MSC-CM: P < 0.05; MSC-CM: P < 0.0001; Figure 5a, b, f, g). The Western blot analysis revealed a significant upregulation of iNOS protein expression in the LPS group compared with the control group (P < 0.001). Conversely, the iNOS protein expression levels were significantly reduced in the LPS+MSC-CM and MSC-CM groups compared with the LPS group (LPS+MSC-CM: P < 0.001; MSC-CM: P < 0.0001). In addition, the expression of Arg1 protein was significantly increased in the LPS+MSC-CM and MSC-CM groups (LPS+MSC-CM: P < 0.001; MSC-CM: P < 0.0001; Figure 5c–e). In conclusion, MSC-CM can promote the polarization of LPS-stimulated BV2 cells toward a pro-resolving state and reduce the polarization toward a pro-inflammatory state, as evidenced by the in vivo experimental findings.

MSC-CM decreased BV2 cell polarization to pro-inflammatory state and increased polarization to pro-resolving state in response to LPS. (a) The expression of iNOS and DAPI in each group was detected by immunofluorescence, iNOS was shown in red, DAPI in blue for the nuclei. Scale = 50 μm. (b) The expression of Arg1 and DAPI in each group was detected by immunofluorescence. Arg1 was red and DAPI was blue, indicating the nuclei. Scale = 50 μm. (c) Western blot detection of iNOS and Arg1 protein expression in each group; (d) Statistical analysis of iNOS protein in each group; (e) Statistical analysis of Arg1 protein in each group; (f) Statistical analysis of fluorescence expression of iNOS in each group. (g) Statistical analysis of expression of Arg1 in each group. Data is expressed as mean ± standard deviation. *P < 0.05,***P < 0.001,****P < 0.0001.
MSCs promoted the polarization of microglial to pro-resolving state and decreased the polarization to pro-inflammatory state by down-regulating ROCK2
As Rho/ROCK plays an important role in the polarization of microglia, we first detected the protein expression level of ROCK2 in the brain tissues of three groups, and the results showed that compared with the sham group, the protein expression level of ROCK2 in the vehicle group was significantly increased (P < 0.01). However, the expression level of ROCK2 was significantly down-regulated in the MSCs group (P < 0.01; Figure 6c, i). In order to further verify that MSCs can regulate the expression level of ROCK2 in microglia and regulate the polarization direction of microglia by regulating ROCK2, this study was conducted in vitro. The immunofluorescence results of cell slides revealed that that there was no significant alteration in CD206 fluorescence intensity in the LPS group compared with the control group (P > 0.05), while iNOS fluorescence intensity showed a significant increase (P < 0.0001). The results were consistent between the LPS+PA group and the LPS group. Compared with LPS group, fluorescence intensity of CD206 in LPS+MSC-CM group was increased (P < 0.0001). In the LPS+MSC-CM group, there was a notable elevation in CD206 fluorescence intensity (P < 0.0001) and a significant decrease in iNOS fluorescence intensity (P < 0.01) compared with the LPS group. Similar trends were observed in the LPS+Y27632 group and LPS+MSC-CM group (Figure 6a, b, e, f). The Western blot analysis revealed a significant upregulation of ROCK2 and iNOS protein expression in the LPS group compared with the control group (ROCK2: P < 0.05; iNOS: P < 0.05), with no significant change in Arg1 expression (P > 0.05). In contrast, the LPS+MSC-CM group exhibited a significant decrease in ROCK2 and iNOS expression levels (ROCK2: P < 0.001; iNOS: P < 0.05) and a significant increase in Arg1 expression (P < 0.01) compared with the LPS group. The expression patterns in the LPS+PA group were similar to the LPS group, but were reversed upon addition of MSC-CM. The results of the LPS+Y27632 group had the same trend as those of LPS+MSC-CM group (Figure 6d, j–l). To further investigate the regulatory impact of MSC-CM on pro-inflammatory mediators, TNF-α and IL-6 expression levels were assessed using reverse transcription–polymerase chain reaction (RT-PCR). Results demonstrated a significant increase in mRNA expression of pro-inflammatory factors (TNF-α and IL-6) in the LPS group compared with the control group (TNF-α: P < 0.001; IL-6: P < 0.0001); Expression levels of LPS+PA-induced proinflammatory factors in the LPS+PA group were consistent with those in the LPS group (TNF-α: P < 0.0001; IL-6: P < 0.0001 vs Control group). LPS+MSC-CM treatment led to a downregulation of TNF-α and IL-6 mRNA expression compared with the LPS group (TNF-α: P < 0.01; IL-6: P < 0.0001). In addition, IL-6 expression in the LPS+Y27632 group significantly decreased compared with the LPS group (P < 0.01). Furthermore, the LPS+PA+MSC-CM group exhibited a significant reduction in TNF-α and IL-6 mRNA expression levels compared with the LPS+PA group (TNF-α: P < 0.0001; IL-6: P < 0.0001); TNF-α expression in the LPS+Y27632 group was notably decreased as well (P < 0.05; Figure 6g, h). In conclusion, ROCK2 protein is upregulated in the GMH model, while MSCs can downregulate its expression. Both ROCK2 and iNOS proteins are upregulated in LPS-stimulated BV2 cells. The ROCK agonist PA can enhance pro-inflammatory state polarization and elevate pro-inflammatory factor mRNA levels in BV2 cells, synergizing with LPS, but this effect can be counteracted by MSC-CM. Conversely, the ROCK inhibitor Y27632 promotes pro-resolving state polarization in BV2 cells, aligning with the effects of MSC-CM, indicating that MSCs can modulate BV2 cell polarization toward pro-resolving state and diminish pro-inflammatory state polarization by suppressing ROCK2 protein expression.
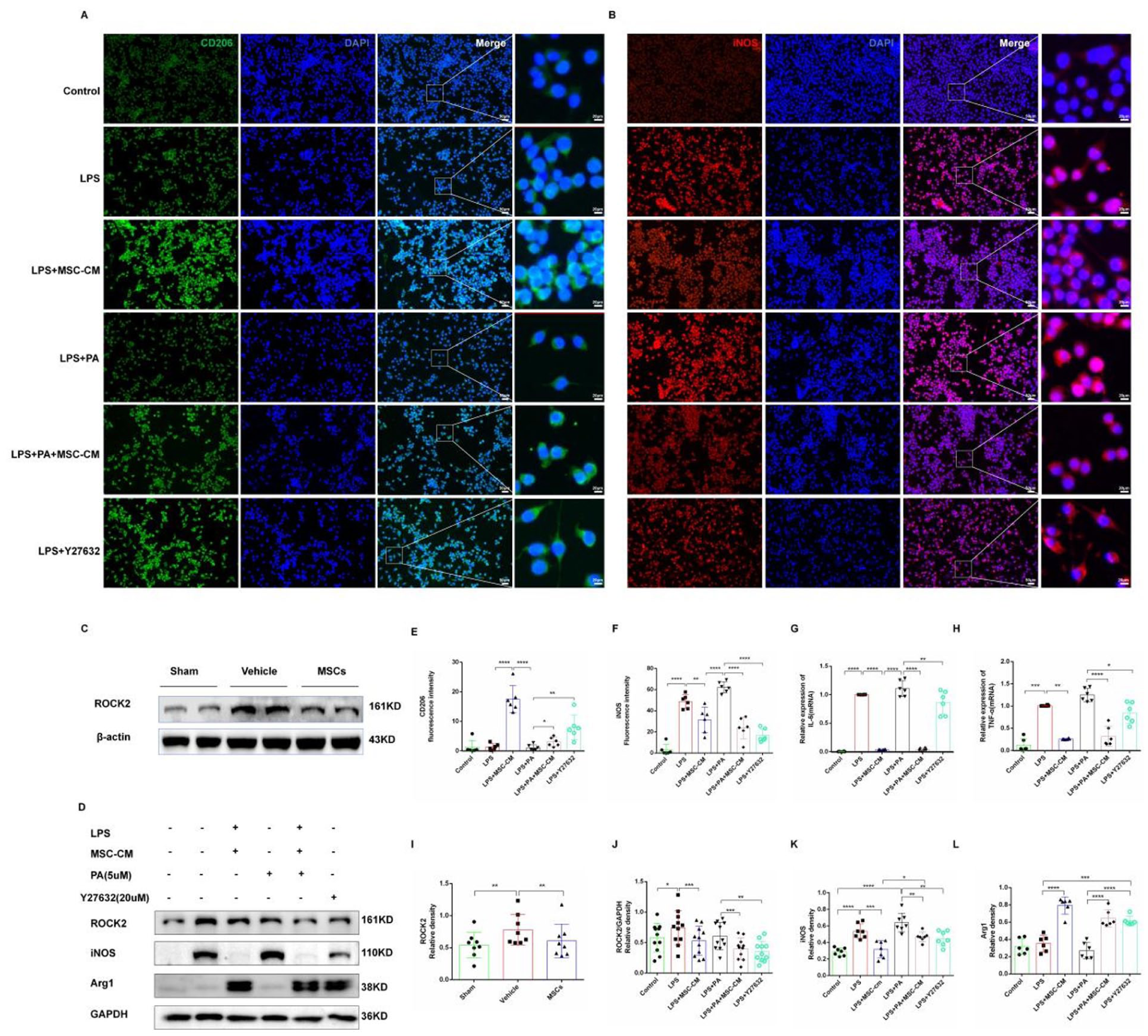
MSCs regulate microglia polarization by down-regulating ROCK2. (a) Immunofluorescence detected CD206 and DAPI expression in each groups. CD206 was green; DAPI was blue, indicating the nuclei. Scale = 50 μm. The picture on the right was the enlarged picture in the white box. Scale = 20 μm. (b) Immunofluorescence detected iNOS and DAPI expression in each group. iNOS was red, and DAPI was blue, indicating the nuclei. Scale = 50 μm. The picture on the far right was the enlarged picture in the white box. Scale = 20 μm. (c) Western blot detection of ROCK2 protein expression in the brain tissues of rats in three groups. (d) Western blot analysis of the expression of ROCK2, iNOS, and Arg1 in BV2 cells. (e) Statistical analysis of CD206 expression in each group. (f) Statistical analysis of iNOS expression in each group. (g) The relative quantitative statistical analysis of TNF-α in each group was detected by RT-PCR. (h) Relative quantitative statistical analysis of IL-6 in each group was detected by RT-PCR. (i) Statistical analysis of ROCK2 protein expression in the brain tissue of three groups. (j) Statistical analysis of ROCK2 protein expression in each group. (k) Statistical analysis of iNOS protein expression in each group. (l) Statistical analysis of Arg1 protein expression in each group. Data are expressed as mean ± standard error. *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
Discussion
Severe IVH complicated with GMH is likely to lead to hydrocephalus and other neurological sequelae. However, there is still a lack of effective treatment methods to improve brain injury and prevent the development of Posthemorrhagic Hydrocephalus (PHH) after severe GMH-IVH, and surgical treatment is only performed in severe hydrocephalus. Therefore, there is an urgent need to develop new treatment strategies to improve the prognosis of severe GMH-IVH in preterm infants. MSCs have positive migration ability, neuroprotection, immunomodulatory and paracrine effects, which have attracted much attention in the treatment of nervous system diseases. Studies have shown that MSCs show great neuroprotective potential in various brain injuries, including ischemic stroke 15 , multiple sclerosis 16 , traumatic brain injury 17 , Alzheimer’s disease 18 , Huntington’s disease 19 , Parkinson’s disease 20 , spinal cord injury 21 , and intravenicular hemorrhage 22 .
In our study, MSCs were observed to have a certain brain protective effect on GMH, which is in agreement with other studies. The GMH model was constructed by collagenase VII-S injection. Brain sections showed hemorrhage in the right germinal stroma and ventricle, and ventricle enlargement was seen 2 weeks later, similar to secondary hydrocephalus (PHH) after GMH-IVH in newborns, indicating that collagenase injection is one of the more objective methods to construct GMH rat models. In our study, the neuroprotective effect of MSCs on GMH rats was verified from the structure and function of brain tissue, respectively. Brain sections and histology proved that MSCs could reduce the increase of cerebral ventricle volume after GMH. Negative geotaxis test, rotary rod test and water maze test proved that MSCs could improve the motor, learning and memory ability of rats after GMH. These results tentatively suggest that MSCs are a potential therapeutic modality to help mitigate brain damage caused by GMH.
After the occurrence of GMH-IVH, the main components of blood such as red blood cells, white blood cells, macrophages, platelets and plasma proteins (thrombin, complement, plasminase, etc.) can enter the ventricle through the ependymal cells, causing primary brain injury and secondary brain injury23,24. Microglia will be the first glial cells to respond. They play an important role in the neuroinflammatory response after intracerebral hemorrhage and are also an important cause of secondary injury. Studies have shown that activated microglia can be found around hematoma within 1–4 h of intracerebral hemorrhage 25 . Activation of microglia is thought to play a dual role in cerebral hemorrhage. The main functions of activated microglia include phagocytosis of hematoma and cell debris to maintain tissue homeostasis and promote nerve function recovery 26 . However, in this process, activated microglia also release a variety of harmful cytokines, such as pro-inflammatory factors, chemokines, reactive oxygen species, protease, NO synthase, prostaglandin, etc.27–29, thereby increasing the degree of brain injury after cerebral hemorrhage; therefore, regulating the polarization of microglia is a key target of current therapy 30 . Current studies have shown that MSCs have immunomodulatory effects, including the effect on the polarization of macrophages/microglia, which is manifested as promoting the polarization of anti-inflammatory pro-resolving state and reducing the polarization of pro-inflammatory state 31 . In our study, it was also found that MSCs can regulate the polarization of microglia after GMH, which is manifested as an increase in the number of pro-resolving microglial cells and a decrease in the number of pro-inflammatory microglial cells. A similar effect was observed when concentrated MSC-CM was applied to BV2 cells, suggesting that MSCs can regulate microglial polarization by secreting substances rather than by direct contact with the cells.
Under physiological conditions, microglia are usually in a homeostatic state, highly branched, with tertiary and quaternary branching structures, but they are rapidly activated under pathological conditions, and their cell morphology will also undergo corresponding changes, showing that the cell body is enlarged, the process is shorter, and the cell shape is round or rod-shaped32,33. The stability and plasticity of the cytoskeleton is one of the key factors in the polarization of microglia 34 . When the cytoskeleton is regulated, the morphology and function of microglia will change accordingly. Cytoskeleton influences the polarization of microglia by regulating the change of cell morphology and the regulation of intracellular signaling. The regulation of cytoskeleton involves several signaling pathways, including Rho GTPases signaling pathway 35 , PI3K/Akt signaling pathway 36 , MAPK/ERK signaling pathway 37 , Wnt signaling pathway 38 , and extracellular matrix signaling pathway 39 , among which Rho GTPases signaling pathway is an important regulatory mechanism. Rho-associated kinase (ROCK), a key kinase in the serine-threonine kinase family, is the main downstream effect on of small GTPase RhoA, containing two subtypes ROCK1 and ROCK2, which have similar structure and function. While ROCK2 dominates in brain and muscle. Rho/ROCK signaling pathway plays an important role in cell growth, development, differentiation and migration, especially in neurological diseases such as neurodegenerative diseases, stroke, neuroinflammation and so on40–42. In recent years, ROCK, as a molecular target, has been considered to be the key to controlling microglia morphology, migration and the release of inflammatory factors 43 . The study showed that the uptake activity of BV2 cells increased after blocking ROCK, and the cell morphology changed, which was manifested as the cell volume increased, many small dendrites appeared, and the shape was more irregular. Some studies have suggested that inhibiting the expression of ROCK protein can promote pro-resolving state polarization of microglia. ROCK is highly expressed in activated macrophages, showing pro-inflammatory state and neurotoxicity 42 . ROCK is an upstream molecule of the NF-κB pathway 44 , which can positively regulate the expression of pro-inflammatory microglial cytokines and negatively regulate the expression of pro-resolving microglial cytokines by inducing the activation of NF-κB 45 . Y-27632, an inhibitor of ROCK, can prevent the activation of NF-κB 46 , thus affecting the expression and secretion of pro-inflammatory microglial and pro-resolving microglial markers. ROCK has been shown to be an effective target for neurodegenerative diseases, and inhibitors of ROCKs have shown promising therapeutic effects in neurodegenerative diseases. RhoA/ROCK is activated when stimulated by histamine, thrombin, vascular endothelial growth factor, lipopolysaccharide, and mechanical action47,48. The RhoA/ROCK pathway may also be activated in the intracerebral hemorrhage model49,50. In the rat cerebral hemorrhage model, ROCK2 increased at 6 h, peaked at 3 d, and decreased at 7 d, but it was still higher than the pre-intervention level. In a LPS-induced BV2 cell model, the Rho/ROCK pathway is thought to play an important role in inducing inflammation 51 . In our study, it was also observed that ROCK2 protein expression was up-regulated in the GMH model, and down-regulated in the MSCs group. The expression level of ROCK2 protein was also up-regulated after LPS was used to stimulate BV2 cells, and down-regulated after the addition of concentrated MSC-CM, which was consistent with the experimental results in vivo. Moreover, the mRNA levels of pro-inflammatory microglial markers (iNOS) and pro-inflammatory factors (TNF-α and IL-6) increased after the addition of PA, the agonist of ROCK, while the effect of Y27632, the inhibitor of ROCK, was reversed, suggesting that MSCs regulate microglia polarization through down-regulation of ROCK2.
It is worth mentioning that in our study, although intraventricular injection of MSCs greatly increased the number of MSCs homing to intraventricular cells, exogenous MSCs still could not survive in vivo for a long time. In vivo tracking of MSCs using GFP showed that the fluorescence intensity of GFP gradually decreased with the extension of transplantation time. By day 28 of transplantation, almost negligible green fluorescence was seen only around the ventricles. Therefore, we speculated that the intravenial environment may not be suitable for the long-term colonization and infinite proliferation of exogenous HUCMSCs, especially the existence of adverse environment such as hemoglobin, thrombin, a large number of inflammatory factors and chemokines in the intravenial brain after hemorrhage exacerbates the death of exogenous HUCMSCs, which also proves that the cell replacement role of HUCMSCs is not the main mechanism of its role, but its strong paracrine role may be one of the mechanisms of its main role. It should be noted that the presence of transplanted cells in the bilateral hippocampi, especially in the contralateral hemisphere, may not solely represent active migration of transplanted cells. Some degree of passive diffusion during the stereotaxic injection procedure could also contribute to the distribution of cells in the contralateral side, which is difficult to completely avoid in intracerebral transplantation. This potential limitation should be considered when interpreting cell distribution patterns. HUCMSCs cannot survive in vivo for a long time and proliferate indefinitely, which may be the reason for their low tumorigenic risk. However, improving the survival rate and survival time of HUCMSCs in vivo may enhance their neuroprotective effect and greatly improve long-term prognosis and nerve repair, which is a direction for future research.
It should be noted that tissue fixation, processing, and sectioning can alter brain morphology and lead to shrinkage or distortion, which may affect the quantitative assessment of ventricular volume. Although we attempted to standardize the histological procedures, the precision of the 1.5 mm sectioning technique may still introduce some variability. Compared with in vivo magnetic resonance imaging, which allows for accurate and noninvasive volumetric measurements, histological methods are inevitably affected by processing-induced artifacts. Therefore, these technical limitations should be taken into consideration when interpreting the present results.
Conclusion
In conclusion, HUCMSCs regulate microglia polarization by down-regulating ROCK2 protein level, regulate neuroinflammation, and have neuroprotective effects on GMH rats, providing a new idea for the therapeutic mechanism of HUCMSCs.
Footnotes
Acknowledgements
Not applicable.
Ethical considerations
Animal experiments: The animal study was reviewed and approved by The Animal Care and Use Committee of Guizhou Medical University (approval No. 2101208, approved on 8 July 2021). All procedures involving animals were conducted in accordance with the relevant ethical guidelines to minimize animal suffering. Tissue collection from women: The collection of human umbilical cord tissue (for Wharton’s jelly-derived MSCs isolation) was reviewed and approved by the Ethics Committee of the Affiliated Hospital of Guizhou Medical University (approval No. 631, approved on 13 July 2021). All procedures were performed in accordance with the Declaration of Helsinki.
Consent to participate
Written informed consent was obtained from all pregnant women prior to umbilical cord tissue collection.
Author contributions
M.Y. designed the research and wrote the manuscript. J.X. prepared and completed the BM-MSCs’ isolation and culture. S.W. helped complete the animal experiment and collected the data and performed the statistical analysis. M.C. provided useful suggestions on experiment design and reviewed the manuscript, provided funds collection. X.Z. assisted with reviewing, editing the manuscript, and provided expertise and feedback.
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by the Science and Technology Department of Guizhou Province (ZK [2022] General 415), the Health Commission of Guizhou Province Scientific Research Project (gzwkj 2022-136) and the project for Ministry of Education on Birth Defects and Related Maternal and Child Diseases (2025KF03).
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Data availability statement
The datasets generated for this study are available on request to the corresponding author.
Statement of human and animal rights
This article does not contain any studies with human or animal subjects.
Statement of informed consent
There are no human subjects in this article and informed consent is not applicable.