
Abstract
Severe acute respiratory syndrome coronavirus (SARS-CoV-2) first emerged in December 2019 in Wuhan, China, and has since spread rapidly worldwide. As researchers seek to learn more about COVID-19, the disease it causes, this novel virus continues to infect and kill. Despite the socioeconomic impacts of SARS-CoV-2 infections and likelihood of future outbreaks of other pathogenic coronaviruses, options to prevent or treat coronavirus infections remain limited. In current clinical trials, potential coronavirus treatments focusing on killing the virus or on preventing infection using vaccines largely ignore the host immune response. The relatively small body of current research on the virus indicates pathological responses by the immune system as the leading cause for much of the morbidity and mortality caused by COVID-19. In this review, we investigated the host innate and adaptive immune responses against COVID-19, collated information on recent COVID-19 experimental data, and summarized the systemic immune responses to and histopathology of SARS-CoV-2 infection. Finally, we summarized the immune-related biomarkers to define patients with high-risk and worst-case outcomes, and identified the possible usefulness of inflammatory markers as potential immunotherapeutic targets. This review provides an overview of current knowledge on COVID-19 and the symptomatological differences between healthy, convalescent, and severe cohorts, while offering research directions for alternative immunoregulation therapeutic targets.
Introduction
Severe acute respiratory syndrome coronavirus (SARS-CoV-2) and its associated pathology, COVID-19, have been of particular concern recently due to the worldwide disruption they have caused 1 . As the disease continues to spread, the development of prophylactic and therapeutic approaches is urgently required. The number of patients requiring intensive care is a critical point in this pandemic. A better understanding of the pathophysiology leading to disease progression is thus urgently needed, especially because the factors that drive severe lung pathology during infection with highly pathogenic human coronaviruses are poorly understood 2 .
The pathogen itself or an overwhelmed immune response can cause infected lesions 3 . Focusing on SARS-CoV-2, studies have observed that this virus can trigger immune responses that can be dysregulated in severe patients, leading to further injury to multiple organs 4,5 . An unbalanced immune response and exacerbated release of proinflammatory cytokines contribute to major complications of coronavirus infection, including acute respiratory distress syndrome (ARDS) and pulmonary fibrosis 6,7 . Yet the immune components that drive severe lung pathology during infection with highly pathogenic human coronaviruses are not clearly understood. Therefore, development of any potential treatments should focus not only on directly killing the virus or preventing infection with a vaccine, but also on managing immune and inflammatory responses 8 .
In this review, we investigated current knowledge on SARS-CoV-2 infection, focusing particularly on its immunologic features. We aimed to detail the mechanisms involving the host innate and adaptive immune responses against COVID-19 and to present information on recent COVID-19 experimental data. We found which immune components and secreted cytokines can induce systemic immune responses and histopathology in patients with COVID-19. Our findings elucidate the changes in immune mechanisms, the immune microenvironment, and immunopathogenesis upon SARS-CoV-2 infection. They provide a new research direction for developing alternative immunoregulation therapeutic targets and may assist in the discovery of potential immune-related hallmarks to define patient recovery and outcome.
Immune System Activation Under SARS-CoV-2 Infection
Humans have two types of immunities: innate (rapid and non-specific response) and adaptive (slow and specific response). In the innate immunity, myeloid cells, such as monocytes, macrophages, dendritic cells (DCs), and granulocytes, are critical in defending against foreign pathogens. Innate lymphocytes, such as natural killer (NK) cells and innate lymphoid cells (ILCs), also work in the innate immune response. In the adaptive immune system, B cells and T cells are crucial lymphocytes 9 . Immune response to SARS-CoV-2 involves both the innate and adaptive immunity 2 .
ACE2 Receptor-Mediated Inflammatory Response
At first contact with the respiratory mucosa, the SARS-CoV-2 virus infects cells via expression of the surface receptors angiotensin-converting enzyme 2 (ACE2) and type II transmembrane serine protease (TMPRSS2) 10 . Viral replication and release then occur, causing the host cell to undergo pyroptosis and release damage-associated molecular patterns (PAMPs), including viral nucleic acids and oligomers. The host’s initial innate immune function is the first line of defense against viral infection 11,12 . In the first days after infection, innate immune cells, including macrophages, DCs, neutrophils, and NK cells, initially activate by recognizing viral PAMPs via pattern-recognition receptors (PRRs) 13 .
Upon SARS-CoV-2 binding to the ACE2 receptor to enter the target cell, the renin–angiotensin system activates and angiotensin II (Ang II) level increase. Elevated Ang II levels are strongly associated with viral load and lung injury in patients with severe COVID-19 14,15 . In addition, studies have shown that the ACE2–Ang II axis can induce cytokine secretions, including interleukin 6 (IL-6), monocyte chemotactic protein 1 (MCP-1), vascular cell adhesion molecule 1 (VCAM-1), and selectin E, to induce macrophage infiltration, endothelial dysfunction, thrombin formation, and impaired fibrinolysis 14,16 –18 . ACE2 appears to play two roles in COVID-19. It first acts as a receptor for SARS-CoV-2 to enter the host and then, due to the increase in Ang II expression from the ACE2–AngII axis, it later recruits additional macrophages to the site of infection 14 .
Host Inflammatory Response
The activated innate immune system generates various pro-inflammatory cytokines and chemokines, including IL-6, CXCL10 (IP-10), macrophage inflammatory protein 1α (MIP1α), MIP1β, and MCP-1. These chemokines then attract monocytes, macrophages, and T cells to the infection site, promoting further inflammation and establishing a pro-inflammatory feedback loop, including the recruitment of NK cells 2 . In addition, innate antigen-presenting cells (APCs), such as DCs and macrophages, will also be at the infection site to present viral antigens to virus-specific T cells. This leads to activation of the body’s adaptive immunity, which is mediated by virus-specific B (humoral immunities) and T cells (cellular immunities) 8,19 .
In the adaptive immunity, cell-mediated immunity and antibody production have critical roles in COVID-19 infection 20 . Decreased absolute numbers of T lymphocytes (CD4+ and CD8+ T cells) occur in both mild and severe cases, with a more notable decrease observed in severe cases. In fact, a decrease in CD8+ T and B cells and an increase in the CD4+/ CD8+ ratio may be independent predictors of poor treatment outcomes 20 . Decreased IFN-γ expression by CD4+ T cells is also more notable in severe cases than in moderate ones 21 . Overall, the low innate and adaptive antiviral defenses and high pro-inflammatory cues lead to multi-organ damage and contribute to COVID-19 22 .
Gender Differences in COVID-19
While men and women have similar susceptibility to SARS-CoV-2 infection, evidence suggests that men with COVID-19 tend to exhibit more severe morbidity and mortality than do women 23,24 . Public meta-analyses have also shown that the odds of requiring admission to an intensive treatment unit for COVID-19 is three times higher among men. Furthermore, the mortality rate among men is 2.4 times higher 24 , and the case fatality rate (CFR) for men was shown to be 1.7 times higher than that for women across 38 countries 25 . In SARS patients, sex was observed to have had a role in mortality 24 .
To further understand differences between sexes, studies have shown that biological sex has direct effects on the immune components of innate and adaptive immunity by modulating genetic variants 26 –32 , transcription factors 33 , epigenetic modification 34,35 , sex hormones (including: estrogens 36,37 , progesterone 38 –42 , and testosterone 43 , and microbiome variances 44 . Therefore, sex may to some extent determine the host immune response to COVID-19 infection as well as the disease course and clinical outcome 44 . Given that women are at 8-9 times higher risk of developing autoimmune disease (AD) compared to men, it is possible that upon COVID-19 infection, their increased immune function may enhance their anti-inflammatory regulation and antiviral defense 45 –47 .
Clinical Symptoms
The abnormal and exaggerated cytokine storm that occurs in critically ill COVID-19 patients is a major factor of COVID-19 mortality. Development of a cytokine storm is also a sign of disease escalation in other viral diseases, such as SARS, Middle Eastern respiratory syndrome (MERS), and influenza 48 –50 . In COVID-19, hyperactivity of the immune system stimulates elevated cytokines including IL-6, IL-8, IL-1β, IL-2, IL-4, IL-7, IL-10, IFN-γ, TNF-α, MCP-1, GM-CSF, CCL2, CCL3, CCL5, and CXCL10 (IP-10). This results in a cytokine storm and lung immunopathology symptoms, including acute lung injury (ALI), systemic inflammatory response syndrome, and ARDS 51,52 . Moreover, intravascular coagulation, blood vessel thrombosis, and hyper-fibrinolysis coexist in COVID-19-induced ARDS and become more severe as the disease progresses 53 –55 . Changes in immune mechanisms, the immune microenvironment, and immunopathogenesis upon SARS-CoV-2 infection are described in the following sections and summarized in Fig. 1.
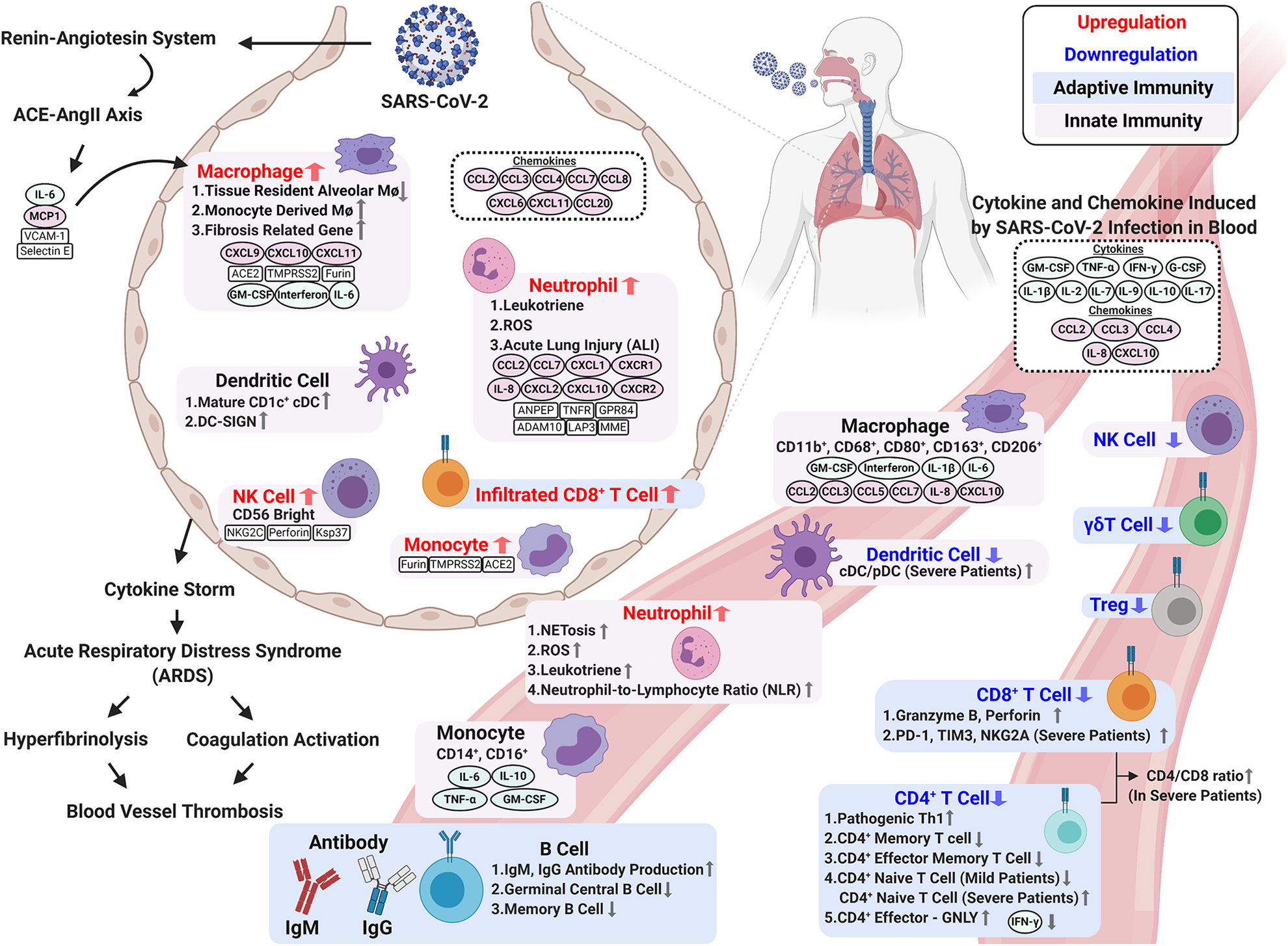
Changes in immune mechanisms, the immune microenvironment, and immunopathogenesis upon SARS-CoV-2 infection. Immune response to SARS-CoV-2 infection involves innate and adaptive immunity. Activated innate immune cells trigger a strong immune response to secrete cytokines, which cause a cytokine storm and ARDS. Elevated circulating cytokines (e.g., IL-1β, IL-2, IL-7, IL-9, IL-10, IL-17, G-CSF, GM-CSF, IFN-γ, and TNF-α) are present in patients with severe COVID-19. BALF samples from COVID-19 patients contain accumulations of various immune cell-attracting chemokines (e.g., CCL2, CCL3, CCL4, CCL7, CCL8, CCL20, CXCL6, IL-8, CXCL10 (IP-10), and CXCL11). A cytokine storm during SARS-CoV-2 results in ARDS, which induces intravascular coagulation and hyperfibrinolysis and causes high thrombus burden in COVID-19 patients. In terms of adaptive immunity, SARS-CoV-2 infection significantly decreases total adaptive immunity lymphocytes and impairs their ability to defend against the virus. Upon infection, CD4+ T cells differentiate less frequently into Th1 cells, and this is associated with the decreased IFN-γ production for antiviral response. Severe COVID-19 patients exhibit the exhausted phenotype CD8+ T cell with high PD-1 and Tim-3 expression. Interestingly, compared to mild cases, severe COVID-19 patients have higher counts of activated CD8+ T cells in circulation to produce cytotoxic granzyme B and perforin. The humoral response is less affected by the virus. The increase in activated B cells gives greater antibody production and better protection to eliminate the virus. Figure created with BioRender.
Innate Responses
The innate immune system is the first line of defense against infection from various pathogens, including viruses. Cells composing the innate immune system include myeloid cells (monocytes, macrophages, DCs, and granulocytes), cytotoxic lymphocyte NK cells, and ILCs (Group 1, Group 2, and Group 3) 12 . These cells orchestrate the direct clearance and eradication of pathogens and contribute to the generation of long-lived adaptive immune responses 56 .
The innate immune response to coronaviruses can be induced upon recognizing pathogen associated molecular patterns (PAMPs), including SARS-CoV-2 positive-sense RNA genomes, viral RNA in endosomes, and viral protein PRRs such as TLR-5 13 . This results in the excessive accumulation of monocytes, macrophages, and neutrophils in the lungs 57,58 . In the innate immunity, APCs such as DCs and macrophages can be used to process viral antigens and activate the host’s humoral and cellular immunities 8,19 . There are two major classes of MHCs involved in antigen presentation: MHC I and MHC II. SARS-CoV mainly depends on MHC I molecule presentation 59 ; by contrast, SARS-CoV-2 could influence the antigen presentation of SARS-CoV-2 variants to MHC II with a single mutation 60 . The innate immune cell subset changes and cytokine secretions in SARS-CoV-2 infection are described in the following sections and summarized in the Table 1.
Representative Innate Immune Cell Subset Changes and Cytokine Secretions in SARS-CoV-2 Infection.

Macrophages and Monocytes in COVID-19
Mononuclear phagocytes contribute substantially to innate and adaptive immunity by sensing and responding to microbial threats by producing inflammatory molecules that eliminate pathogens. Circulating monocytes are the largest type of leukocyte and can differentiate into tissue-resident macrophages and myeloid lineage DCs 85 . Resident subcapsular sinus macrophages and hilar lymph node macrophages are the majority of macrophages presenting viral antigens to activate specific T cells, which secrete cytotoxic perforin, granzyme B, and interferon-γ to kill infected cells. Thus, these macrophages play a protective role against viral infection by capturing viral particles 86 .
Upon COVID-19 infection, circulating monocytes and resident macrophages participate in all stages of SARS-CoV-2 87 . Human monocytes and macrophages seem to be a widespread target upon COVID-19 infection due to the expression of ACE2, TMPRSS2, and furin on both cells 64 . In vitro, both SARS-CoV and SARS-CoV-2 can equally infect type-I and -II pneumocytes as well as alveolar macrophages 88,89 . A highly proinflammatory macrophage microenvironment is present in the lungs of patients with severe COVID-19 61 , and autopsies on COVID-19 patients have revealed high infiltration of macrophages within the area of bronchopneumonia 70,90 . Moreover, with ACE2 and nucleoprotein (NP) antigen expression, CD68+ and CD169+ macrophages are highly infiltrating to the lymph node subcapsular sinus and splenic marginal zone of COVID-19 patients 58 . Furthermore, COVID-19 may selectively induce macrophages to produce IL-6, not TNF-α or IL-1β, to induce lymphocytopenia or lymphocyte necrosis 91 . Therefore, the depletion of tissue-resident alveolar macrophages and an abundance of inflammatory monocyte-derived macrophages are more associated with severe COVID-19 cases 58,61,70,92 .
Monocytes exhibit significant morphological and functional differences in patients with COVID-19, especially those requiring prolonged hospitalization and ICU admission 68 . The CD14+ CD16+ monocytes subset is found more in the peripheral blood of COVID-19 patients compared to healthy populations, and these larger-than-normal monocytes are easily detected on forward scatter in flow cytometry 68 . In severe COVID-19 cases, classical monocytes exhibit higher expressions of type I interferons (IFNs), TNF, and IL-1-driven inflammatory responses, but not with mild cases; this suggests that the exacerbating inflammatory cytokines may be critical in the progression to severe COVID-19 22 .
In addition to CD14+CD16+, larger-than-normal CD11b+, CD68+, CD80+, CD163+, CD206+ macrophages can also be found in the peripheral blood of patients COVID-19. Since CD80 is considered an M1 marker, while CD163 and CD206 are considered M2 markers, both M1 and M2 macrophages along with monocytes are present in the peripheral blood cells of patients with COVID-19 68 . Moreover, in moderate and severe COVID-19 cases, a relatively higher frequency of M1-like macrophages and higher concentration of chemokines, such as CXCL9, CXCL10 (IP-10), and CXCL11, can be found in the bronchoalveolar lavage fluid (BALF) compared to healthy individuals 61,93 .
Highlighting the effect of SARS-CoV-2 on macrophages, the cytokine storm in COVID-19 cases tends to fit the pattern of macrophage activation syndrome with lymphocytopenia. Immunohistochemical testing shows that infected macrophages produce high levels of IL-6 rather than TNF-α in COVID-19-infected spleen tissue and lymph nodes compared to healthy tissue. The SARS spike protein activates increased transcription of several cytokines in macrophages. These include granulocyte-macrophage colony-stimulating factor (GM-CSF), IL-6, CXCL10 (IP-10), IL-1β, IL-8, and IFNs 64 . However, SARS-CoV-2 does not activate any IFN response, including that of IFN type I, II, and III, from infected monocyte-derived macrophages (MDMs) 82 .
BALF samples from COVID-19 patients show accumulations of monocyte-attractant chemokines in the lungs, including CCL2, CCL3, CCL4, CCL7, CCL8, CCL20, CXCL6, and CXCL11 93,94 . In addition, the activated blood-circulating macrophages further release attractant chemokines (such as CCL2, CCL3, CCL5, CCL7, and CXCL10 (IP-10), as well as of the soluble form of the α-chain of the IL-2 receptor) to locally accumulated mononuclear macrophages 95 . Many other elevated circulating cytokines are also present in patients with COVID-19, especially in those requiring ICU admission, thus triggering severe cytokine syndrome and exacerbating the severity of the disease 58,85 . These include IL-1β, IL-2, IL-7, IL-9, IL-10, IL-17, G-CSF, GM-CSF, IFN-γ, TNF-α, IL-8, CXCL10 (IP-10), MCP-1 (CCL2), MIP1A (CCL3), and MIP1B (CCL4), and many of these cytokines and chemokines are secreted from monocytes and macrophages 93,96 .
Finally, single-cell RNA sequencing (scRNA-seq) and bulk RNA-seq analysis of the heterogeneity of macrophages in patients’ BALF with moderate and severe COVID-19 reveals abundant MDMs in severe cases 61 . Twenty macrophage subclusters have been identified 61 , with one subset of COVID-19-associated macrophages exhibiting higher tissue repair and fibrosis-related genes, similar to that seen in liver cirrhosis 65 . This suggests that the pathogenicity of infiltrating macrophages could extend beyond the promotion of acute inflammation and is in line with the fibrotic complications observed in patients under mechanical ventilation 97 .
Natural Killer Cells
NK cells, an innate cytotoxic lymphocyte, provide innate immune defense against acute viral infection and cancer. Compared to healthy people, patients with moderate or severe COVID-19 have low peripheral NK cell and lower total lymphocyte numbers 20,98 –102 . Yet compared mild cases, patients with severe COVID-19 have significantly lower total lymphocytes but no significant difference in NK cell numbers 20 .
The single-cell landscape of immune cells in the BALF of patients with COVID-19 exhibits increased NK cell numbers upon infection 12,61,103 . Flow cytometry reveals strong NK cell activation in the peripheral blood of patients with COVID-19, and single-cell RNA sequencing of BALF from COVID-19 patients shows two clusters of NK cells linked to disease status. Activated NK cell clusters show high levels of perforin, NKG2C, and Ksp37 expression, which is associated with disease severity, and the CD56bright NK cluster is present in severe hyperinflammation COVID-19 cases 12 .
For cytotoxic lymphocytes, such as NK cells and CD8+ T cells, NKG2A is an inhibitory receptor to recognize “self” MHC I on target cells 104,105 . As an inhibitory receptor, it induces NK cell and CD8+ T cell exhaustion in cancer and chronic viral infections 106,107 . In severe COVID-19 cases, increased NKG2A expression can induce NK and CD8+ T cell exhaustion and trigger cytotoxic lymphocytes to have impaired cytokine production 56,84 . Thus, the functional exhaustion of cytotoxic lymphocytes appears to be associated with SRAS-CoV-2 infection.
Neutrophils
The COVID-19 immunopathology can be exacerbated by neutrophil recruitment and related activity. Neutrophils are recruited by infected endothelial cells and form web-like chromatin structures known as neutrophil extracellular traps (NETs) as the forefront of innate immunity 108 . Patients with COVID-19 have elevated specific markers of NETs, including myeloperoxidase-DNA and citrullinated histone H3, proving that SARS-CoV-2 can activate NETosis 109,110 . In addition, compared with COVID-19 survivors, nonsurvivors have higher white blood cell (WBC) and neutrophil counts but lower lymphocyte and platelet counts 111 . The peripheral blood of patients with COVID-19 exhibits kinetic changes in WBCs, neutrophils, and monocytes, as well as in different lymphocyte subsets. Compared with mild cases, patients with severe COVID-19 show significant increases in total WBC counts at the time of onset (within 3 days) but not during the following period of disease progression 70,71 .
Upon viral infection, neutrophils are involved in early antiviral defense 112 . The initial pyroptotic of the infected endothelial airway can induce IL-1β overexpression and trigger neutrophils migration and T-cell activation 76,113 . The activated neutrophils further release leukotrienes and reactive oxygen species (ROS), thus triggering cytotoxicity such as ALI and a cytokine storm. Neutrophils may also cause endothelial injury, which can further promote viral systemic dissemination 76 –78,80 . Through degranulation and lysis, neutrophils can be cytotoxic 114 and can aggravate lung inflammation upon SARS-CoV-2 infection 115,116 . In addition, RNA-seq analysis of BALF samples shows that multiple neutrophil-attracting chemokine genes are upregulated in the COVID-19 infected lung microenvironment, and these include TNFR, IL-8, CXCR1, CXCR2, ADAM10, GPR84, MME, ANPEP, and LAP3 81 .
An increased peripheral neutrophil-to-lymphocyte ratio (NLR) can be seen in severe SARS-CoV-2 cases and is likely associated with an unfavorable prognosis 81,84,117 . This correlates with low expression of type I and III IFNs and high expression of pro-inflammatory factors and chemokines 22 . Increased serum level of the NLR ratio and lower percentages of monocytes, eosinophils, and basophils have been associated with disease severity and death. Severe cases tend to also have higher NLR and increasing inflammatory cytokines, chemokines, and other markers (including C-reactive protein, ferritin, and D-dimers) 58,72,75 . Furthermore, four contributing variables, namely WBC count, neutrophil count, neutrophil-to-CD8+ T cell ratio (N8 R), and NLR, are considered potential prognostic factors of the disease 70,71 . NLR appears to be the most useful prognostic factor affecting the prognosis in severe cases 117 .
The COVID-19 immunopathology can be exacerbated by neutrophil recruitment and related activity. Excessive upregulation of the neutrophil-attracting chemokines, including CXCL1, CXCL2, IL-8, CXCL10 (IP-10), CCL2 and CCL7, can be found in COVID-19 BALF samples 93,94 . From an anatomopathological perspective, patients who die from COVID-19 show marked lung infiltration by neutrophils 73 . In severe cases, microscopic examination of the autopsy lung tissues shows an increased number of neutrophils and histiocytes (acute bronchopneumonia) in the airspaces 71,90 .
Dendritic Cells
Dendritic cells (DCs) are a diverse group of professional APCs that provide an important bridge linking the innate and adaptive immune responses. DCs played a crucial role in two previous human coronavirus outbreaks: SARS and MERS-CoV 80,118 . In both diseases, like COVID-19, clinical manifestation includes rapid and progressive acute pneumonia with altered multiorgan functions. However, compared to MERS-CoV, which infects monocyte-derived DCs (moDCs) and rapidly generated high levels of IFN-γ 80,118 , SARS-CoV-2 does not activate any IFN response from infected moDCs 82 .
DCs comprise several subsets, including plasmacytoid DCs (pDCs) and conventional DCs (cDCs) 119 . Upon COVID-19 infection, total peripheral DC populations are reduced and their activity is impaired 80,83,120 . Decreased CD123+ pDCs and CD141+ DCs and increased CD1c+ cDCs have been reported in the lungs and bronchoscopy infiltrates 67,80 . In addition, when severe acute infection occurs, the ratios of CD123+ pDCs to CD11c+ cDCs decrease and significant functional impairment occurs 80,83 . Since pDCs are critical for type I IFN secretion and the initial antiviral response, the reduction in pDCs could contribute to acute COVID-19 pathogenesis and have implications for treatment response 83,120 .
Moreover, aging causes a reduction in plasmacytoid DC numbers, as well as their pathogen-sensing functions via TLRs 120,121 . In addition, the DC-specific intracellular adhesion molecule-grabbing nonintegrin (DC-SIGN) is a C-type lectin receptor present on the surface of both macrophages and DCs to recognize viral PAMPs. In older adults, the increased DC-SIGN expression may be involved in the pathogenesis of more severe SARS-CoV-2 infection. The combination of SARS-CoV-2 and age-mediated DC dysfunctions may be central to the increased susceptibility of severe infection and poor outcomes in older patients 120 .
Adaptive Immunity
Lymphocytes, especially T cells and B cells, carry out the adaptive immune response in the human body. Although they can take days or even weeks to become established, differentiated lymphocytes have an important role in controlling and shaping the immune response by providing various immune-related functions and long-lasting protection. The T cells can either kill an infected cell (cytotoxic CD8+ T cells or CTLs) or balance the immune response (CD4+ helper T cells), while the B cells are related to antibody production, also known as the humoral immunity. Each of cell type is critical for eliminating SARS-CoV-2 infection 122 . Cytotoxic CD8+ T lymphocytes, CD4+ effector-GNLY (granulysin), and NK cells are also necessary for controlling viral infection, and the functional exhaustion of cytotoxic lymphocytes correlates with disease progression 123 –125 .
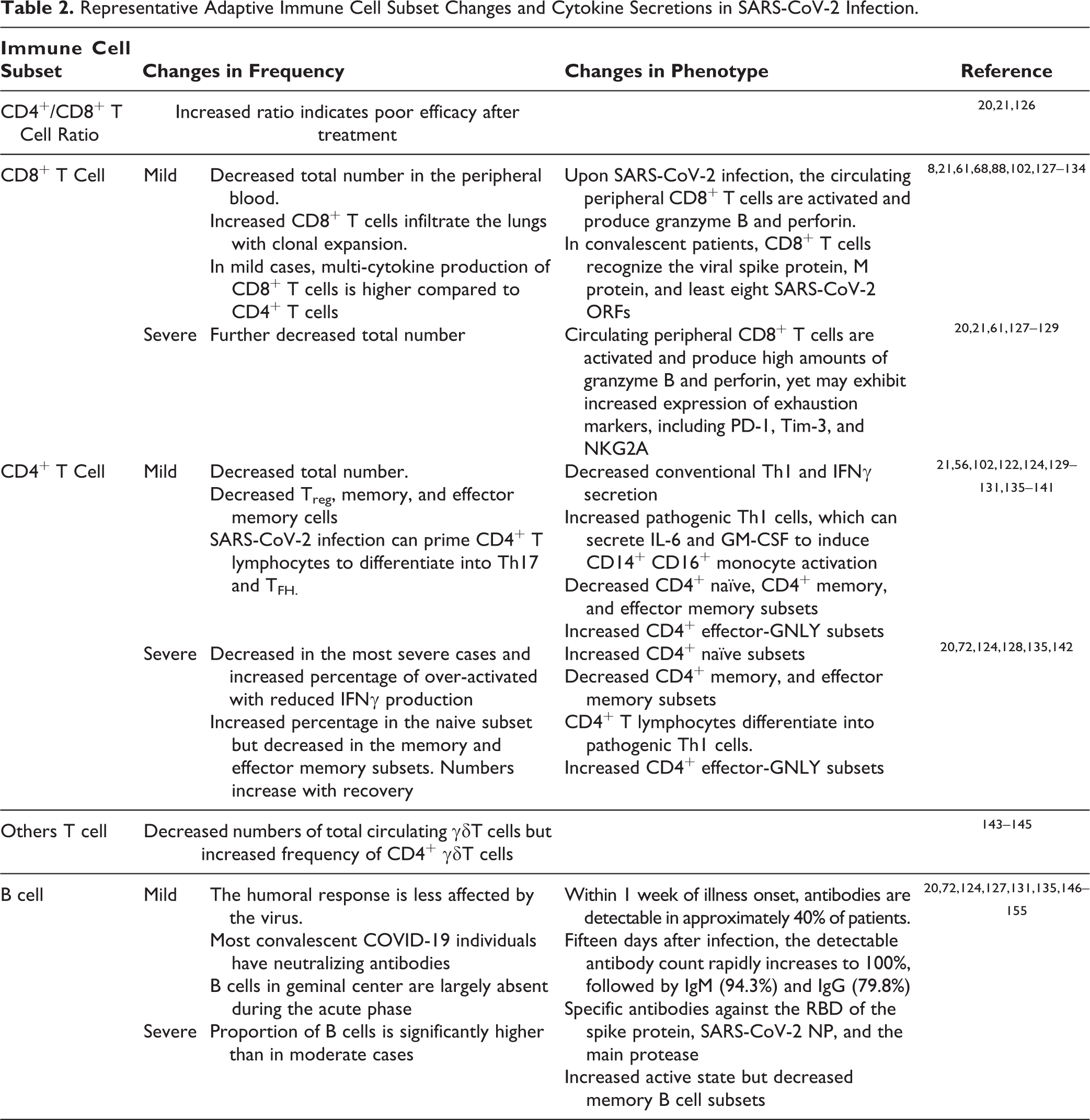
Adaptive immunity, including cell-mediated and humoral immunity, is critical in regulating COVID-19 infection 20 . Patients with COVID-19 exhibit a decrease in total lymphocytes and the mean counts of the three main lymphocyte populations (T, B, and NK cells), and this is more pronounced in severe cases. In particular, T and NK cells decrease markedly below normal levels, while B cells remain within the lower normal range 72 . Severe cases are not only associated with decreased total lymphocyte numbers, but also with increased naive T helper cells as well as decreased memory T helper cells and memory subsets 72 . Effector T cells, such as IFN-γ secreting CD4+ T cells, also tend to be lower in severe cases than in moderate cases. In addition, dysregulation of lymphocytes, including T cell exhaustion, occurs in most cases. Among all the adaptive immune cells, CD8+ T, CD4+ T, and B cells (and the CD4+/CD8+ ratio) are significantly associated with inflammatory status and are indicated as independent predictors of poor treatment outcomes 20,21 . The adaptive immune cell subset changes and cytokine secretions in SARS-CoV-2 infection are described in the following sections and summarized in the Table 2.
Representative Adaptive Immune Cell Subset Changes and Cytokine Secretions in SARS-CoV-2 Infection.
T Cells
Serious lymphopenia occurs when the absolute numbers of total T cells, CD4+ T cells, and CD8+ T cells are below the lower limit of normal. It can happen in patients with COVID-19, with more profound reductions observed in severe cases than in moderate cases 21,66,102,131,156 . Lymphopenia is correlated with p53 signaling overactivation 69 accompanied by atrophy of the lymph nodes and spleen 137 . CD26 and CD147 inducing activation-induced cell death may also be responsible for lymphopenia 88 . Moreover, the more profound the lymphopenia, the worse the prognosis 50,51 .
Although with a lower T cell total count, the pervasiveness of highly cytotoxic effector T cell subsets (e.g., CD4+ effector-GNLY, CD8+ effector-GNLY, and NKT CD160) is related to convalescence in cases of moderate COVID-19 124 . Present in COVID-19 convalescent individuals are virus-specific CD4 and CD8 T cells that can recognize multiple regions of the SARS-CoV-2 N protein and S protein epitopes as well as antibodies against the viral receptor-binding domain (RBD) 149,157 . In addition, patients with long-lasting memory T cells toward SARS are most likely to generate SARS-CoV-2-specific T cell immunity and recover from SAR-CoV-2 infection. Pre-existing T cells preferentially boost N-specific T cells, whereas individuals with no history of SARS tend to have NSP7-, NSP13-, and ORF1-specific T cells 157 .
Regarding the CD4+/CD8+ T cell ratio, some studies have reported no significant difference in the CD4+/CD8+ T cell ratio, indicating that both CD4+ and CD8+ T cells are depleted during SARS-CoV-2 infection 20,131 , while multivariate analysis results have suggested that the CD4/CD8 ratio is significantly higher in severe cases than in moderate cases, with a higher ratio indicating a worse inflammatory status and poorer efficacy posttreatment 20,126 .
Finally, the professional APCs process viral antigens to activate virus-specific T cells and cytotoxic T lymphocytes (CTLs), by major histocompatibility complexes (MHC; or human leukocyte antigen, HLA, in humans) 8,19 . Disease in patients infected with SARS-CoV with HLA-B*46:01 (MHC I molecules) genotypes is more severe compared to those with different genotypes, although this has not been clinically validated for SARS-CoV-2 8,9,158 –161 .
CD8+ T Cell Response in COVID-19
Analysis of lymphocyte subsets associates COVID-19 patients with lymphopenia. Although with lymphopenia, infiltrated lymphocytes can still be observed in the lungs infected with SARS-CoV-2, and scRNA-seq analysis of BALF from COVID-19 patients exhibits an increase in CD8 T cell infiltrate with proliferation and clonal expansion 61 . SARS-CoV-2-specific CD8+ T cell are present in approximately 70% of convalescent patients, and these cells can recognize the viral spike protein, M protein, and at least eight SARS-CoV-2 ORFs 130 .
There appears to be heterogeneity in the CD8+ T cell immune response between patients. Some studies have reported that upon SARS-CoV-2 infection, the circulating peripheral CD8+ T cells are activated and produce high amounts of granzyme B and perforin to increase their cytotoxic response. In addition, these CD8+ cells appear to be clonally expanded in the lungs of mild COVID-19 diseases 133 . In mild cases, patients have more multi-cytokine production CD8+ T cells than CD4+ T cell 132 . Other studies have reported that CD8+ T cells in severe COVID-19 patients become exhausted due to the increased expression of exhaustion markers, including PD-1, Tim-3, TIGIT, HLA-DR, CD38, CD25, and NKG2A 128,129 .
CD4+ T Cell Response in COVID-19
Upon infection, activated CD4+ T cells, also known as T helper (Th) cells, produce pro-inflammatory cytokines (IFN-γ), and antiviral cytokines, such as granzyme B and TNFκ 129,162 . Compared with healthy people, patients with COVID-19 exhibit a decrease in total CD4+ T cell numbers and IFN-γ production, with even lower IFN-γ levels occurring in severe cases than in moderate cases 21 . In mild cases, patients have lower proportions of the naive, memory and effector memory CD4+ T cell subsets. In severe cases on the other hand, patients have increased naïve helper CD4+ T cell, yet memory T cell subsets (CD3+CD4+CD45RO+) are decreasing 72,124,130 . In contrast to memory or naïve-state cells, the proportion of CD4+ effector-GNLY (granulysin) is elevated in patients with COVID-19 and convalescence patients 124 .
There are five types of CD4+: T helper (Th)1, Th2, and Th17; regulatory T cells (Treg), and follicular helper T cells (TFH) 138 . SARS-CoV-2 infection can prime CD4+ T lymphocytes to differentiate into pathogenic Th1 cells (nonconventional Th1), Th17, and TFH 21,135,139,141,163 . Normally, upon infection, activated Th1 effector T cells produce pro-inflammatory cytokines (IFN-γ) and antiviral cytokines, such as granzyme B, TNF-α, TNF-β, and TNF-κ 129,162 to stimulate innate and T cell immune responses. In COVID-19 patients, conventional Th1 effector cells show a decrease in the total amount 140,141 and lower IFN-γ production 21 . SARS-CoV-2 infection can also prime CD4+ T lymphocytes to differentiate into pathogenic Th1 cells and generate IL-6 and GM-CSF to promote activation of CD14+ CD16+ monocytes 139 . Furthermore, increased T cells (TFH) can help B cells to produce antibodies and drive CD8+ T cells’ cytotoxic response activation to kill virus-infected cells 138 . In addition, the count of Treg cells, which downregulate the induction and proliferation of effector T cells, is significantly lower in COVID-19 patients 21,137 , and this manifests as a lack of functional immunosuppression. Notably, a high NLR, reflecting a worsening of the inflammatory process, has been closely related with poor prognosis, disease severity, and death 164 .
Other T Cells
Patients with COVID-19 have lower total circulating γδT cell counts and higher frequency of CD4+ γδT cells. This is notable because the CD4+ subset of γδT cells produce Th2 cytokines, which can then induce TMPRSS2 expression (the SARS-CoV-2 spike protein priming protease). Th2 immune responses, while able to increase TMPRSS2 expression, also decrease ACE2 expression 143 –145
B Cell and Humoral Response
B cells are critical adaptive immune cells for antibody production and the humoral immune response 122 . Upon COVID-19 infection, T lymphocyte responses are impaired by the virus, while B cell activities are not as affected as T cells 135 . In severe cases, the proportion of B cells is significantly higher than in moderate cases (20% vs. 10.8%) 72,131,135 . In addition, patients with COVID-19 exhibit higher proportions of activating B subsets and decreased memory B cells compared with healthy individuals 124 . Moreover, most convalescent patients with COVID-19 have detectable cellular immune responses and neutralizing antibodies 135,149 .
Most of them also display antibody responses between 6 and 21 days after infection 150 –152 . Within 1 week of illness onset, antibodies are detectable in approximately 40% patients. Fifteen days after infection, the detectable antibody number rapidly increases to 100%, followed by IgM (94.3%), and IgG (79.8%). A strong positive correlation can also be observed between the Ab titer and clinical severity 15 days post infection 148 . In addition, SARS-CoV-2-specific antibodies against the RBD of the spike protein 149 , SARS-CoV-2 NP, and the main protease 135 .
The longevity of the antibody response remains unknown, yet most patients show high levels of SARS-CoV-2-specific memory B cells at 6-8 months post infection, suggesting that patients start to develop long-term protective immunity 154 . In the case of SARS or MERS-CoV infection, specific antibodies begin to wane at 12 to 52 weeks following infection and homologous re-infection can occur 153 . Serology testing is relatively ineffective for early detection because the detectability of virus-specific antibodies in the first 7 days from onset is below 40% 148 .
Moreover, the germinal center (GC) is a specialized area within the lymph nodes, where B cells can receive help from follicular T helper (TFH) cells to activate and become memory B cells or long-lived plasma cells. SARS infection can defeat GC responses, thus affecting B cell activity. As for COVID-19, GCs are also largely absent during the acute phase, and this absence is accompanied with a lack of BCL6-expressing B cells or TFH cells 155 .
Conclusions
This review investigated the various changes to immune mechanisms, the immune microenvironment, and COVID-19 immunopathogenesis. The COVID-19 pandemic is a current issue affecting people worldwide, and there is an urgent need to develop fundamental therapeutic interventions. Controlling the inflammatory response may be as important as targeting the virus. Thus, a better understanding of the immune processes and mechanisms is crucial for potential COVID-19 therapeutic drug development. Future research should consider focusing on identifying inflammatory biomarkers as potential immunotherapeutic targets and investigating the association between immune dysfunction and disease severity in SARS-Co-V-2 patients. The findings would be invaluable for vaccine development and evaluation.
Footnotes
Authors Note
Chih-Hung Ye and Wen-Lin Hsu contributed equally to this article.
Author Contributions
Conceptualization, S.-H. Yu, W.-L. Hsu, C.-H. Ye.; writing—original draft preparation, S.-H. Yu, W.-L. Hsu, C.-H. Ye.; writing—review and editing, S.-H. Yu, W.-L. Hsu, C.-H. Ye. All authors have read and agreed to the published version of the manuscript.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was financially supported by the Young Scholar Fellowship Program by Ministry of Science and Technology (MOST) in Taiwan, under Grant MOST 109-2636-B-002-012.