
Abstract
Hepatocyte transplantation (HcTx) is a promising approach for the treatment of metabolic diseases in newborns and children. The most common application route is the portal vein, which is difficult to access in the newborn. Transfemoral access to the splenic artery for HcTx has been evaluated in adults, with trials suggesting hepatocyte translocation from the spleen to the liver with a reduced risk for thromboembolic complications. Using juvenile Göttingen minipigs, we aimed to evaluate feasibility of hepatocyte transplantation by transfemoral splenic artery catheterization, while providing insight on engraftment, translocation, viability, and thromboembolic complications. Four Göttingen Minipigs weighing 5.6 kg to 12.6 kg were infused with human hepatocytes (two infusions per cycle, 1.00E08 cells per kg body weight). Immunosuppression consisted of tacrolimus and prednisolone. The animals were sacrificed directly after cell infusion (n=2), 2 days (n=1), or 14 days after infusion (n=1). The splenic and portal venous blood flow was controlled via color-coded Doppler sonography. Computed tomography was performed on days 6 and 18 after the first infusion. Tissue samples were stained in search of human hepatocytes. Catheter placement was feasible in all cases without procedure-associated complications. Repetitive cell transplantations were possible without serious adverse effects associated with hepatocyte transplantation. Immunohistochemical staining has proven cell relocation to the portal venous system and liver parenchyma. However, cells were neither present in the liver nor the spleen 18 days after HcTx. Immunological analyses showed a response of the adaptive immune system to the human cells. We show that interventional cell application via the femoral artery is feasible in a juvenile large animal model of HcTx. Moreover, cells are able to pass through the spleen to relocate in the liver after splenic artery infusion. Further studies are necessary to compare this approach with umbilical or transhepatic hepatocyte administration.
Keywords
Introduction
Orthotopic liver transplantation is still the treatment of choice for various advanced chronic and acute liver diseases. However, due to the shortage of donor organs this treatment is unavailable for many patients. Hepatocyte transplantation (HcTx) has previously been evaluated as a possible alternative to liver transplantation. HcTx allows using organs which are not eligible for transplantation 1 as well as partial livers to maximize the number of recipients from one liver 2 . HcTx has been evaluated for the treatment of acute liver failure 3 , with mixed results in adults and children 4,5 . The biggest potential, however, is seen in congenital metabolic liver diseases 6 . Here, encouraging results have been shown for Crigler–Najjar syndrome Type I 5 –7 , urea cycle disorders 5,6,8 , and Factor VII deficiency 5,9 .
Although various cell application routes are being explored, such as infusion through the hepatic vein 10 or intrahepatic injection 11,12 , the most common application route for hepatocyte administration to the liver remains the portal vein 13 . Nonetheless, the portal vein access has shown instances of portal vein thrombosis following HcTx 14,15 .
The spleen is a common ectopic site for HcTx 13 , and is primarily seen as an option in patients with impaired liver structure 16 . This may be due to findings that HcTx into the spleen could induce hepatization of the spleen 17,18 , which leads to a lower risk of pulmonary embolism otherwise posed by portocaval collaterals 16 . Other research suggests that cells infused into the splenic artery relocate into the liver 18,19 as commonly observed in rodents—though it is not clear whether or not this really happens in humans. The transfemoral access to the splenic artery for HcTx has been well evaluated in adults 18,20,21 , and has the potential to be a safe and easily accessible application route. Previous trials have suggested a lesser risk for thromboembolic complications after cell infusion to the spleen and, possibly, subsequent translocation to the liver 22 . However, there are few clinical experiences in the young infant or neonate 23 . Since congenital metabolic liver diseases are primarily diseases of neonates and infants, more research on this application route is warranted. Therefore, preclinical trials using a suitable animal model to evaluate the feasibility and efficacy of transfemoral intrasplenic HcTx via the splenic artery are needed.
The juvenile Göttingen minipig is a suitable preclinical model for the human infant because of the similarities in human and porcine physiology 24 –26 , as well as a vascular system that allows for transfemoral catheterization 27 . At 11 weeks old a Göttingen minipig weights 4–5 kg, which is approximately the weight of the human infant 28 .
Using a juvenile large animal model of Göttingen minipigs and human hepatocytes, which can be clearly followed by immunohistochemistry, we aimed to evaluate the feasibility of transfemoral catheterization of the splenic artery with multiple cell infusions, providing insight on engraftment, translocation, and long-term viability as well as thromboembolic complications.
Materials and Methods
Animal Studies
Experiments were performed on four female Göttingen minipigs weighing 7.3 kg (pig 1) 5.3 kg (pig 2), 6.1 kg (pig 3), and 12.6 kg (pig 4). All experiments were performed with full approval by the state Office of Health and Local Affairs (LAGeSo, Berlin, Germany; Reg No. G 0260/13) and under supervision by the local welfare office. The animals were kept in the animal care facility of the Department of Experimental Medicine of the Charité – Universitätsmedizin Berlin (certified by ISO 9001:2008). Two animals were sacrificed directly after transplantation, one animal on day 2 after the intervention, and one animal was sacrificed on day 18 after the first intervention.
Catheter Intervention
All catheter interventions were performed by the same interventional radiologist (P.P.) of the Department of Radiology, Charité – Universitätsmedizin Berlin. The common femoral artery was punctuated under ultrasound guidance and a 3 French polyurethane sheath was placed in the femoral artery. A 3 French C2-catheter was placed in the splenic artery according to the individual anatomy of the animals. Placement of the catheter was confirmed via digital subtraction angiography. When angiography showed a prominent gastroepiploic artery, it was closed with microcoils during the initial intervention to prevent cell translocation to the gastric wall. Once the catheter was placed, it remained in place until the end of the cell application protocol.
Cell Preparation and Handling
Human hepatocytes were provided by Cytonet GmbH & Co. KG, Weinheim, Germany as a suspension of heterologous human hepatocytes, representative for cell suspensions used in clinical trials in children aged 0–5 years with urea cycle disorders (CCD02 in Germany and CCD05 in USA/Canada).
The suspensions were prepared from non-transplantable donor organs using a two-step collagenase perfusion of the liver tissue (further details cannot be given due to intellectual property protection). Donors had a negative serology for HIV, Hepatitis B and C, and other transmittable diseases. All suspensions used were tested negative for microbiological growth. Cells were transported cryopreserved in liquid nitrogen. The suspensions used were thawed in a water-bath at 40°C, counted, and checked for viability using the trypan blue exclusion test, transferred into syringes without resuspension and immediately infused. The viability aimed for in cell suspensions after thawing was 50% to 65% viable cells, and the cut off for infusion was viability below 45%. Reference values were chosen in respect of cryopreservation decreasing the ability of hepatocytes to exclude trypan blue by up to 30% 29 : adhering to the clinically required viability of 60% or above would have likely led to a sizable number of cells being discarded. Since human hepatocytes were used, a lower threshold on viability has been set for ethical and economic reasons. The cut off was chosen to be able to consistently reach the goal for viable cells without surpassing the safety limit. No batch fell below the cut off after thawing, with observed cell viabilities ranging from 50.7% to 76.5%.
Cell Application
Each intervention cycle consisted of two cell infusions. The upper limit for cell count per infusion had been set to 1.00E08 cells per kg body weight, based on safety aspects. Viable cell count was aimed at 3.30E07 to 5.00E07 cells. The infusion rate was set to 0.5–1.0 ml/min. The time between infusions was initially set to 8 h but revised to 2 h after the first experiment (pig 1, sacrificed after 2 days). One animal underwent two cell infusion cycles, with the third cell infusion on day 4 after the first infusion. Total infused volume varied by cell density of thawed batches, viability, and animal weight. Total volume per infusion ranged from 49.8 ml to 101.9 ml.
Immunosuppression
On days of cell administration, immunosuppression was induced immediately prior to cell application using i.v. Methylprednisolone at 10 mg/kg body weight (Urbason, Sanofi-Aventis Deutschland GmbH, Frankfurt, Germany). Immunosuppression was continued using 10 mg prednisolone once daily as well as tacrolimus 0.1 mg/kg body weight p.o. twice daily (targeted trough level 10–12 ng/ml), starting from the day prior to the first infusion. The serum blood levels of tacrolimus were determined at blood withdrawals at days 0, 4, 6, 11, and 18. Dosage was adjusted to match desired blood levels of 10–15 mg/ml. Calculated antibiotic and anti-viral regimes were administered to minimize the risk of infection. Tulathromycin 2.5 mg/kg body weight was administered at days 0 and 6 subcutaneously as mycoplasma infection prophylaxis.
Imaging Studies
During each infusion cycle, color-coded Doppler sonography (CCDS) was performed to monitor portal venous and splenic artery blood flow. CCDS was performed directly before and directly after each infusion.
One week prior to all interventions a computed tomography (CT)-exam (Light speed VCT, General Electric Healthcare, Milwaukee, WI, USA) was performed in all animals. Additional CT exams were performed on pig 4 on day six as well as day 18 (i.e., 2 days and 14 days after the second cell infusion).
Anesthesia
Catheter interventions and CT scans were performed under anesthesia. Premedication was administered with an intramuscular injection of 3 mg/kg azaperone (Stresnil; Janssen Cilag, Neuss, Germany), 25 mg/kg ketamine (Ursotamine; Serumwerk, Bernberg, Germaniy), 3.5 mg/kg xylazine hydrochloride (Rompun; Bayer Vital GmbH, Leverkusen, Germany) and 0.03 mg/kg atropine sulfate (Atropin sulfate, B Braun, Melsungen, Germany). The animals were orally intubated (ID 4.0 with cuff, Mallinckrodt, Mallinckrodt Medical, Athlone, Ireland). The animals were then ventilated using standard ventilators (Draeger Zeus Infinity Empowered, Lübeck, Germany) and were kept under general anesthesia with continuous administration of 4–6 mg/kg propofol (Fresenius Kabi AG, Bad Homburg, Germany) and 15–25mg/kg ketamine (Actavis Group PTC EHF, Munich, Germany). Balanced crystalloids were infused as necessary.
Interventional Monitoring
Each animal had a 4.2 French broviac catheter implanted before the first infusion. Using this catheter, blood samples were taken for clinical chemistry and blood gas analysis during cell infusions. From pig 4, additional blood samples were harvested on day 4, 6, 11, and 18 (Fig. 1).

Study design and time of interventions. Experimental set-up. Animals received hepatocyte infusions at time points 0 and, in one case, additional infusions at time point 4. Computed tomography (CT) and color-coded Doppler sonography (CCDS) was performed in the follow-up until sacrifice.
Blood samples for clinical chemistry were handled according to the instructions provided by synlab.vet (Berlin, Germany), who also performed analyses. Blood gas analysis was done using a clinical grade reader (ABL 800 Flex, Radiometer, Krefeld, Germany).
Sacrifice and Tissue Sample Harvesting and Examination
Animals were sacrificed in deep narcosis and organs were harvested following heart arrest. Samples were taken directly after macroscopic examination. Tissue samples of the spleen, liver, and lungs were harvested using a standardized systematic approach, including multiple samples of each liver lobe, each pulmonary lobe, and multiple samples of each region of the spleen. Samples of other organs were harvested at sight of macroscopic anomalies.
Immunohistochemistry
All tissue samples were embedded in paraffin. Human hepatocytes were stained using either human-specific Ab7797 anti-CK18 (Abcam, Cambridge, UK) antibodies in the liver, lungs, or spleen, or liver-specific OCH1E5 antibodies (Santa Cruz Biotechnology, Dallas, TX, USA) in the lung or spleen.
The anti-CK18 antibody and the OCH1E5 antibody were diluted 100-fold. Both were then incubated overnight at 4°C. Antigen retrieval was performed prior to incubation with DAKO pH 9 antigen retrieval solution (Agilent Technologies, Santa Clara, CA, USA) or DAKO high PH antigen retrieval solution (Agilent Technologies) and cooked in a pressure cooker for 5 min. Stains were performed using the alkaline phosphatase–anti-alkaline phosphatase method using DAKO Fuchsin-Chromogen (Agilent Technologies) for the staining reaction. The counterstaining was done using Mayer’s Hematoxylin (AppliChem, Darmstadt, Germany). Hepatocyte count was scaled from grade 0 to grade 3.
Quantification of Anti-Donor-Specific Antibodies
Production of anti-donor IgM and IgG antibodies in recipient serum samples was determined by flow cytometry at the Institute of Medical Immunology (Charité – Universitätsmedizin Berlin). Briefly, donor hepatocytes were incubated with different serum dilutions for 20 min followed by incubation with fluorescein isothiocyanate (FITC)-conjugated goat anti-pig-IgM H/L (Abcam, Cambridge, UK) or FITC-conjugated goat anti-pig-IgG H/L (AbD Serotec GmbH, Puchheim, Germany). Samples were washed and acquired on a BD LSRFortessa (BD Biosciences, San Jose, CA, USA). Data analysis was performed using FlowJo software version 10.1 (FlowJo, LLC, Ashland, OR, USA). The amount of produced antibodies is presented as the geomean shift between serum samples and negative controls.
Statistical Analysis
Statistical analysis was performed using GraphPad Prism 8.0 (GraphPad Software, Inc., La Jolla, CA, USA). Pearson’s correlation was performed to analyze the correlation of transaminase and tacrolimus levels. A p-value less than 0.05 was considered significant.
Results
Transfemoral Access of the Splenic Artery is Feasible
In all cases, angiographic catherization of the splenic artery was successful. The femoral artery was accessible transdermal or via open surgical exploration (Fig. 2). Cell application was feasible, with no signs of stenosis or decreased perfusion in the splenic artery or portal vein in the CCDS during or immediately following cell infusions.

Interventional catheter placement in the splenic artery. A: Catheterization of the celiac trunk with depiction of the (1) common hepatic artery, (2) right gastric artery, (3) splenic artery. B: Contrast accumulation of the spleen (arrow). C: Situation after coiling of the gastroepiploic artery (arrow).
Hepatocytes Translocate from the Spleen to the Liver
On the day of the intervention as well as 2 days later, hepatocytes could be seen in a large number of splenic vessels by immunohistochemistry. Also, in smaller numbers, cells were detected within the parenchyma of the spleen. Immunohistochemical staining of the liver on the day of the infusion as well as 2 days after infusion proved translocation of human hepatocytes from the spleen to the liver. While the largest part of the cells remained in the spleen, a large fraction of cells could be found in the hepatic vessels. In addition, small amounts of cells translocated into the liver parenchyma (Fig. 3). The observed spleen and liver parenchyma showed no signs of necrosis or ischemia, and the structure of the spleen seemed to be largely unaffected. A smaller number of cells were able to pass through the liver into the lungs, where they persisted for up to 14 days after the last infusion. The semi-quantitative evaluation of cell count showed hepatocytes in a sizable fraction of tissue samples observed (Fig. 4).
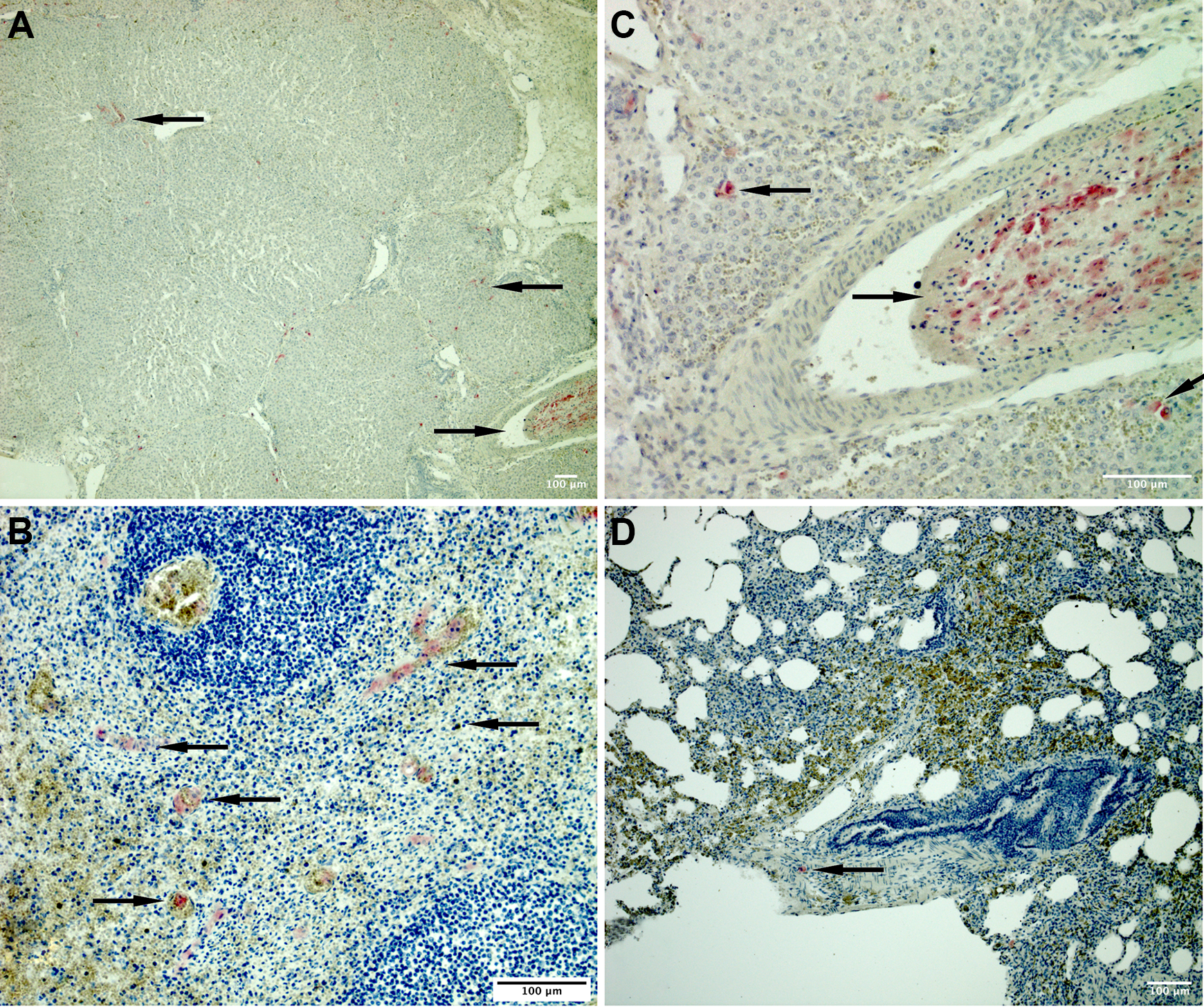
Immunohistochemistry of days 0 and 2 after infusion. A: Liver, day 0, Intraparenchymal human hepatocytes (←) and intravascular human hepatocytes in the portal venous system (→). B: Liver, day 0, Intraparenchymal human hepatocytes (←) and intravascular human hepatocytes (→). C: Spleen on day 2, Intraparenchymal human hepatocytes (←) and intravascular human hepatocytes (→). D: Lung, day 0, Intraparenchymal human hepatocytes (←)
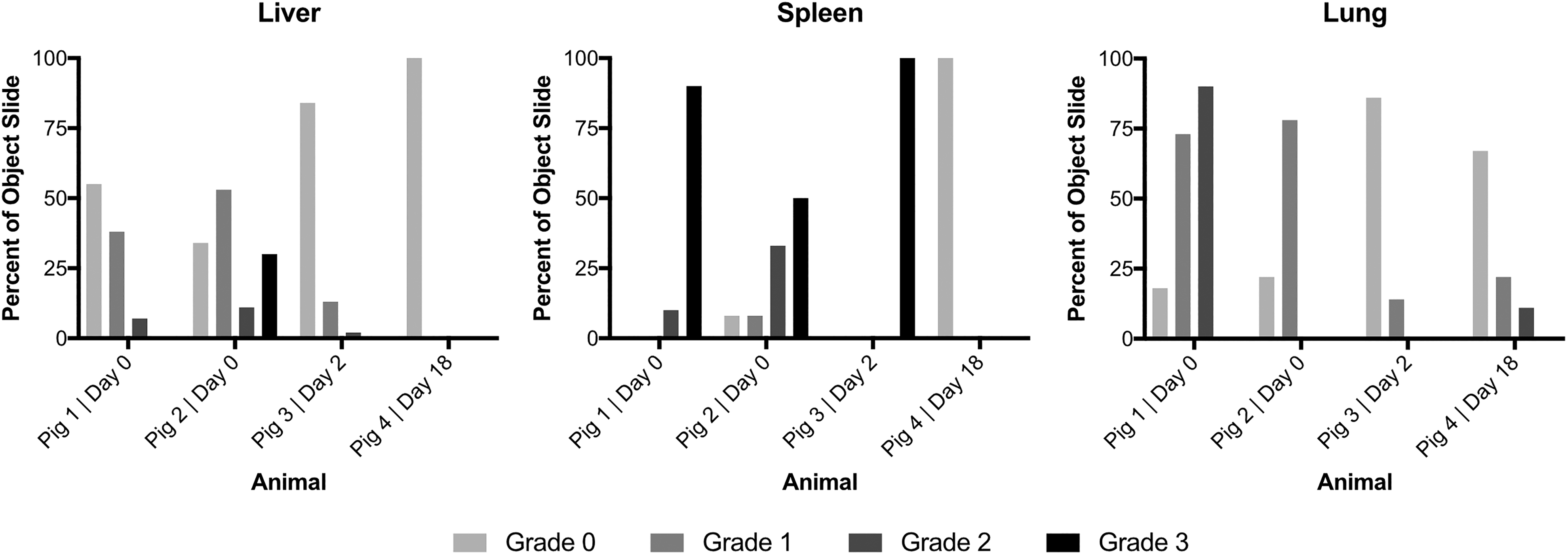
Semi-quantitative cell count by organ and animal. Detection of human hepatocytes, in percentage of slides observed. Slides are assigned grades based on following categorization:
Two Weeks after Infusion, Hepatocytes can be Neither Found in the Spleen Nor in the Liver
Immunohistochemistry failed to detect transplanted human liver cells in the liver 2 weeks after the last cell infusion. Although no hepatocytes were found in the spleen of the animals, the lungs showed residual hepatic cells (Fig. 5).

Immunohistochemistry, day 18 after infusion. At 14 days after infusion, human hepatocytes can neither be found in the spleen nor the liver, but in the lungs. A: Liver, no human hepatocytes visible. B: Spleen, no human hepatocytes visible. C: Lung, OCH1E5 staining. Intraparenchymal hepatocytes (←) and intravascular hepatocytes (→).
Efforts to quantify metabolic activity of human liver cells were cut short due to methodological problems: An enzyme-linked immunosorbent assay test quantifying human albumin was established, and a peak of human albumin levels was observed after the second cell infusion with a subsequent drop, most likely due to contamination of the cell suspension with human albumin (data not shown).
Clinical Chemistry
The clinical chemistry showed a stepwise rise of transaminases in the animal that was sacrificed 2 weeks after the fourth hepatocyte infusion. Maximum serum aspartate aminotransferase (AST) levels were reached at day 4 (408 U/l), highest alanine aminotransferase (ALT) levels were measured at day 6 (430 U/l) of the experiment. A steep rise in gamma-glutamyltransferase was also observed, with levels rising from 40 U/l on day 4 to 981 U/l on day 6 and 1437 U/l on day 11. However, all liver function parameters were close to the normal serum threshold on day 18. Bilirubin stayed within normal range over the course of the experiment. The tacrolimus trough levels dipped below the target levels on day 4 of the experiment (3.12 µg/l) but recovered after adjustment of dosage to 10.80 µg/l at day 6. Over the subsequent period tacrolimus levels remained in therapeutic range, above 10.00 µg/l. By Pearson correlation, a non-significant negative correlation was found between serum tacrolimus levels and transaminase levels (–0.61 for AST, p=0.27; –0.20 for ALT, p=0.74).
Procedure-related Complications
Severe complications occurred in two animals: Both sustained hemodynamically relevant bleeding complications after sheath removal. Subsequently, the femoral artery was ligated following sheath removal, which was tolerated well by the animals. In one case the bleeding caused a drop of hemoglobin levels from 99 g/l to 37 g/l, but the levels normalized a week later.
One animal suffered from necrosis of the gastric wall due to a large number of cells migrating into the gastroepiploic artery and subsequently occluding the left gastric artery. This complication was avoided in all further animals by coiling of the gastroepiploic artery before cell infusion. The coiling procedure had no noticeable adverse effects.
On day 18, signs of acute tubular necrosis were found in the left kidney; however, the procedure itself seems an unlikely cause, since histopathological analysis of the fibrosis of the kidney was determined to be older than 2 weeks.
Moreover, contrast-enhanced CT imaging showed signs of occlusion of the terminal splenic artery on day 6, which was spontaneously resolved by day 18 (Fig. 6).

Contrast CTs of the abdomen, portal venous phase. A: Homogenous spleen 7 days before first cell infusion. B: Inhomogeneous contrast enhancement of the caudal spleen (→) on day 6. C: Homogenous perfusion of the caudal spleen (→) on day 18.
Pathology
The pathological examination of the spleen 18 days after the first intervention showed neointimal proliferation in the arterioles. The media was broadened by myxoid material and vacuolization of the smooth muscle cells. In the vessel walls, fibrinoid necrosis was found with inflammatory infiltrations. Hepatocytes could not be identified in the spleen 18 days after the first transplantation.
The lungs, the liver, the stomach, and the spleen had a macroscopically normal appearance. The left kidney was atrophic, the right kidney presented normally. The histopathological analysis revealed disseminated infectious lesions in the primary and secondary target organs of the hepatocyte infusion (liver, spleen, and lungs), which cannot clearly be linked with the study as such lesions can occur spontaneously in minipigs (personal communication with pathologist). In smaller parts of the lungs, macrophages were found, and the alveolar walls occasional had round cell infiltrations. In the liver, moderate portal and interstitial infiltration of inflammatory cells were discovered with perivascular inflammatory extravasation. The left kidney showed acute tubular necrosis of unknown origin (fibrosis older than 2 weeks).
Generation of Donor Hepatocyte-specific Antibodies
In order to understand why the hepatocytes could not be detected 14 days after infusion we tested whether they elicited an adaptive immune response. Therefore, we tested consecutively collected serum samples for the presence of anti-IgM and anti-IgG antibodies specific to the human hepatocytes used for transplantation. Infusion of hepatocyte from two independent donors (batch #1 und #2) induced production of IgM and IgG antibodies in time-dependent manner (Fig. 7). Clearly, different hepatocyte batches vary in their immunogenicity, as batch #1 elicited a higher increase in antibody production than batch #2.
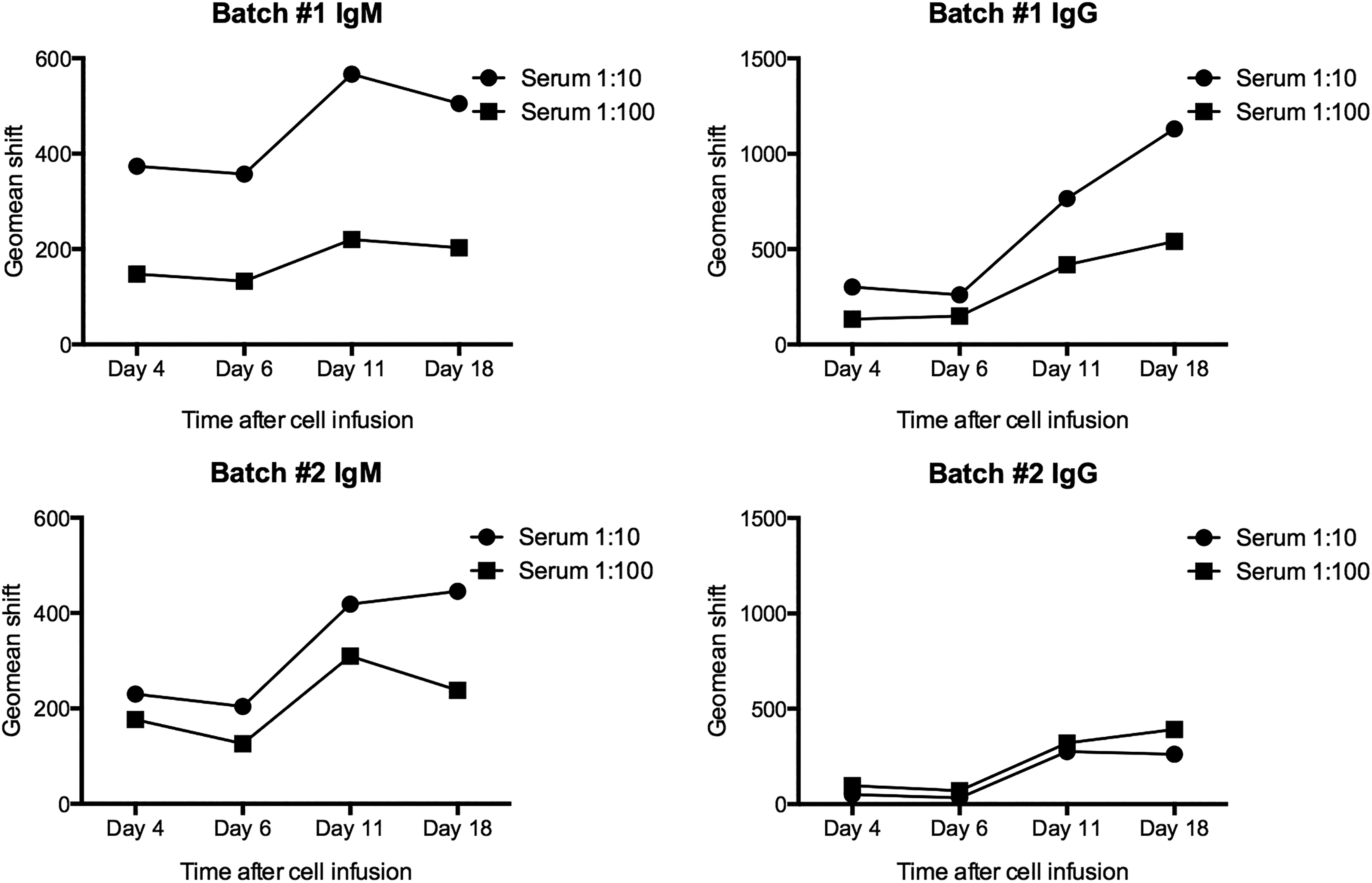
Antibody production. Infusion of hepatocyte induced cell-specific IgM and IgG production in a time-dependent manner (batches #1/2).
Discussion
The large animal model has been important in establishing new techniques in surgery and transplantation. The feasibility of the infant pig model for hepatocyte transplantation has already been shown by Enosawa et al. 30 . The transfemoral catheterization of the splenic artery for hepatocyte transplantation, however, has not been evaluated in a juvenile large animal model. We had previously used this route in a large animal study of the adult landrace pig 31 . However, cell translocation to the liver had not been satisfactorily evaluated due to the experimental protocol, which was based on sacrifice of the animals immediately after transplantation.
The results of our study show that interventional catheter placement through the femoral artery is possible in juvenile pigs, which are similar to human infants in regard to vascular diameter of the respective visceral vessels. We also show that cell application into the splenic artery is feasible. One minor thromboembolic event occurred, which was completely reversible. The pathological examination showed no tissue damage caused by the occlusion.
In addition, cell translocation to the portal venous system and liver was clearly demonstrated, with human hepatocytes detected by immunohistochemistry in up to 67% of tissue samples observed. While for the first days after intervention cells predominantly remained in the spleen, a sizable fraction dislocated to the liver, with a smaller amount surpassing the liver sinuses to engraft in the lungs. Even though a previous study using Indium-111-labeled hepatocytes has shown no significant cell translocation to the lungs following intraportal infusion 32 , cell translocation into the lungs has previously been proven by using the same application route by Bilir et al. 33 . Our experiment shows that cell translocation to the lung was not prevented by the intrasplenic approach, possibly attributed to further migration of hepatocytes from the liver through portocaval collaterals.
However, the total amount of cells migrating to the liver and lungs can only be estimated from the systematic histopathological analysis performed in our study, for lack of quantification assays. Therefore, further studies on migration and long-term survival of liver cells in ectopic sites are needed. In past experiments, iron oxide particles and radiolabeling have been used to quantify cell distribution 22,32 . Limited spatial resolution may limit the sensitivity of nuclear imaging to detect small amounts of cells in ectopic sites. Hepatocytes labeled with micrometer-sized iron oxide particles have been shown not to be negatively affected by the magnetic field of magnetic resonance imaging systems 34 , and should therefore be considered for further experiments. Both techniques necessitate prior marking of cells ex vivo, which was not part of the experimental protocol of in this study.
Since the liver is the natural host of hepatocytes and bile secretion of transplanted hepatocytes could in principle be realized by connection to the biliary system, a translocation to the liver could be beneficial for the long-term engraftment of the cells. However, cell engraftment has proven to be an issue: In the long-term follow-up, the observed number of surviving cells deviated from the desired result, concurring with the findings of Baumgardner and Orosz who have shown that hepatocytes are strongly antigenic 35 . Up to 70% of the cells may be eliminated by the innate immune system, with macrophages clearing the portal vessels of non-engrafted cells 36 . However, a T-cell-mediated reaction of the adaptive immune system seems to be a limiting factor in long-term survival 37 . This in accordance with our immunological analysis, which shows a reaction of the adaptive immune system, despite immunosuppressive treatment. A non-significant negative correlation was observed between transaminase and serum tacrolimus levels. Different factors might play a role in the rise of transaminase levels, such as infusion of necrotic hepatocytes, reaction of the innate immune system, and the time of blood sampling, allowing no clear cause–effect relation from our data.
It is likely that the immune response was amplified by the study design as a xenotransplantation model. Human hepatocytes were chosen to allow detection of the cells via immunohistochemistry without prior marking. Since inter-species transplantation is expected to amplify the immune response, a higher rate of surviving cells may be achieved in an allotransplantation setting. Furthermore, the use of cell batches from different donors in one animal may have additionally led to a higher immunological response: Antibody production differed between cell batches, with one batch eliciting a higher increase in antibody production. It is likely that donor–recipient HLA-matching could therefore improve long-term survival. The relatively high number of necrotic hepatocytes in the cell suspensions (viabilities between 50 to 77% after thawing prior transplantation) could have further boosted the immune response.
Different approaches tackling cell rejection are currently being evaluated. Common approaches are radiation preconditioning of the host organ 38 , gene therapy 39,40 , and microencapsulation 41,42 . With advances in modulating the immune response, the outcome of HcTx should improve. This may lead to higher demand for the procedure, which would favor application routes with a higher clinical availability, such as the one evaluated.
The splenic artery application route has shown no severe thromboembolic complications, and the catheterization of the splenic artery and coiling of the gastroepiploic artery was without adverse effects. The study design, however, was susceptible to other complications: Since the cell application must occur under anticoagulation, the animals were prone to bleeding after catheter removal. While this posed no problem in two of the animals, two other animals suffered hemodynamically relevant bleeding from the puncture site. Standard issue closure systems could not be applied due to the small diameter of the punctured vessel. This is a safety concern for future experiments on the subject. The complications observed are strongly associated with the experimental model: Catheter removal had to be undertaken in non-sedated animals, which prevented sufficient compression of the arterial vessel. Thus, this problem would most likely not have occurred in humans. However, cyanosis of the foot with loss of peripheral pulses has previously been described after transfemoral hepatocyte transplantation in the human infant. In the case described in literature, perfusion and peripheral pulses reverted back to normal over the course of 48 h 18 .
Given the complications observed in the study, as well as those previously described, it cannot be concluded that the femoral artery infusion route is equivalent or superior to the portal vein or transhepatic access. However, since lethal portal vein thrombosis has been described in portal venous access for hepatocyte transplantation 14 , the assessment of alternative routes for hepatocyte infusion is important for the safety in future pediatric hepatocyte transplantation trials. Other application routes such as catheterization of the umbilical vein have been used in clinical studies and might by less prone to the bleeding complications observed in our study 43 ; however, only very limited data are currently available. Therefore, no appropriate comparison between the splenic approach and other application routes can be made so far.
Another issue is monitoring of the metabolic activity of the cells. The measurement of human albumin can give an idea of metabolic cell activity. However, the half-life of human albumin has been reported at 20 days 44 . This has rendered the measurement unreliable, since the transfused cells are immersed in an albumin-rich cell buffer. Due to the small number of surviving cells observed on day 18 after transplantation, no relevant metabolic activity was to be expected.
Our study was designed to be a pilot study for cell transplantation via the splenic artery in the infant, and the possible translocation of the cells. The number of the animals used in the experiment was limited (n=4). Moreover, our study was not designed to exactly quantify the biodistribution of the infused cells. Many of the problems and complications which were seen originate from the animal model, and should not occur in human subjects; since minipigs are very active at this young age, it was necessary to keep them sedated for the entirety of the application protocol. This led to the animals being in anesthesia for up to 13 h. The most invasive part of the protocol was the catheterization itself. There would be no need to continue the anesthesia in human infants once the catheter is secured. The valve should also be able to remain in the femoral artery, making repeated catheterization unnecessary, and therefore lowering the risk of bleeding complications. This should eliminate two major pitfalls of the performed experiment.
In conclusion, we have shown that hepatocyte administration into the splenic artery via the femoral artery is feasible in a juvenile minipig model. We were also able to clearly demonstrate translocation of donor hepatocytes from the spleen to the liver in this preclinical large animal model—thereby verifying the very first findings by Strom et al. who reported that hepatocytes might have migrated from the spleen to the liver in humans after intraarterial infusion to the spleen. However, Strom and colleagues had concerns regarding their findings, which were based on PCR-based detection of the Y gene in the liver, which might have been due to leukocytes transferred with the donor hepatocytes 18 . We here have not been able to ensure long-term engraftment of the cells, and bleeding complications have proven to be an issue. Nevertheless, many of the problems experienced are immanent to the pig model used. With projected advances in preventing rejection of the transplanted cells, the splenic artery route should be further investigated for hepatocyte transplantation, especially regarding safety and efficacy compared with other routes used for cell administration in hepatocyte transplantation.
Footnotes
Acknowledgments
We thank the teams from the Research Institution of Experimental Medicine and of the Department of Radiology of the Charité – Universitätsmedizin Berlin for help with animal care and imaging.
Ethical Approval
All experiments were performed with full approval by the state Office of Health and Local Affairs (LAGeSo, Berlin, Germany; Reg No. G 0260/13).
Statement of Human and Animal Rights
All animal studies were performed in accordance with the federal law and after approval by the local animal welfare agency and under supervision by the local welfare office. The animals were kept in the animal care facility of the Department of Experimental Medicine of the Charité – Universitätsmedizin Berlin (certified by ISO 9001:2008).
Statement of Informed Consent
There are no human subjects in this article and informed consent is not applicable.
Declaration of Conflicting Interests
The author(s) declared the following potential conflicts of interest with respect to the research, authorship, and/or publication of this article: The study was funded by Cytonet GmbH & Co. KG, Weinheim. Dr. Dr. Wolfgang Rüdinger and Dr. Sabine Kafert-Kasting were employed by Cytonet during the study period.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: Nathanael Raschzok is participant of the BIH Charité Clinician Scientist Program funded by the Charité – Universitätsmedizin Berlin and the Berlin Institute of Health. This work was supported by institutional funding of the Charité – Universitätsmedizin Berlin, and by Cytonet GmbH & Co. KG, Weinheim.