
Abstract
The limited availability of human islets has led to the examination of porcine islets as a source of clinically suitable tissue for transplantation in patients with diabetes mellitus. Islets from porcine donors are commonly used in both in vitro and in vivo experiments studying diabetes mellitus. However, there are significant differences in quality and quantity of islet yield depending on donor pig age, as well as substantial differences in the costs of pancreas procurement in adult versus neonatal and juvenile pigs. In this study, we compared the total cost per islet of juvenile pig pancreata with that of neonatal and adult pigs. Although adult porcine pancreata yield, on average, more than five times the amount of islets than do juvenile and neonatal pancreata, we found that the high price of adult pigs led to the cost per islet being more than twice that of juvenile and neonatal islets (US $0.09 vs $0.04 and $0.02, respectively). In addition, neonatal and juvenile islets are advantageous in their scalability and retention of viability after culture. Our findings indicate that isolating neonatal and juvenile porcine islets is more cost-effective and scalable than isolating adult porcine islets.
Introduction
Development of the Edmonton Protocol in 2001 and demonstration of the viability of islet cell transplantation as treatment for type 1 diabetes mellitus has led to increased interest in optimization of human islet transplantation for treatment of this disease 1,2 . However, limited availability of viable human islets and costs of human islet isolation and characterization creates an obstacle for widespread use of islet transplantation for diabetes treatment and has prompted the examination of porcine islets as a clinically suitable substitute 1 .
At present, adult pigs are considered superior in many respects to young (neonatal and juvenile) pigs as a source of islets for xenotransplantation. Young porcine pancreata contain islet-like cell clusters which contain sparse insulin-producing cells in comparison to mature islets 3 . Thus, separation of insulin-producing cells from exocrine cells is more difficult in young pigs 4 –6 . Previous studies have demonstrated that adult porcine islets have greater insulin secretion rates than neonatal and juvenile porcine islets 7 . However, there are potential benefits to utilizing young pig islets, including reduced immunogenicity, increased cell proliferation ability post-transplantation, and increased in vitro stability versus adult islets 8 –10 . Furthermore, neonatal islets have been shown to be effective in restoring normoglycemia after transplantation into diabetic mice 11 . Isolation of adult porcine islets is also labor-intensive and expensive, especially when considering the prolonged husbandry costs required to produce adult pigs. Therefore, utilization of young pigs may potentially mitigate some of these costs 2,12 .
No previous study has compared the cost-effectiveness of neonatal, juvenile, and adult porcine islet isolations. Smith et al. had previously compared the oxygen demand, membrane integrity, and cell function and proliferation between neonatal, juvenile, and adult porcine islets 7 . However, the pricing to carry out each group of experiments was not considered. Therefore, there is a need to understand the cost-effectiveness and scalability of each age group of pigs to facilitate understanding of their roles in islet isolation and xenotransplantation.
To accomplish this objective, we developed a scalable protocol for porcine islet isolation using 18- to 24-day-old juvenile pigs that has shown promise in in vivo transplant studies in rodent models of diabetes. The aim of this study was to determine the actual costs to produce islets from our pre-weaned 18- to 24-day-old juvenile pigs and compare these with the costs of producing islets in neonatal (3 days old) and adult pigs, as reported in the current literature. We reviewed several studies investigating islet yield in adult and neonatal pigs and determined the price per viable islet equivalent (IEQ) of each group of pigs in the context of their respective advantages and disadvantages.
Materials and Methods
Islet Isolation of Juvenile Pigs
Islets were isolated from 18- to 24-day-old male Yorkshire swine (n = 12; S&S Farms, Ramona, CA, USA) using procedures we described previously 13 . Briefly, the pancreas was harvested using rapid surgical procurement (<5 min) and placed in Organ Preservation Solution (Corning Cellgro, Manassas, VA, USA). Cold ischemia time was limited to <30 min. The pancreas was then washed in cold (4°C) Hanks balanced salt solution supplemented with HEPES (Corning Cellgro, Manassas, VA, USA), and trimmed of surrounding adipose and lymphatic tissue in a sterile biosafety hood. The pancreatic tissue was then minced into 2–3 mm2 pieces and digested at 37°C using Sigma Type V Collagenase (2.5 mg/mL in Hanks balanced salt solution; Sigma Aldrich, St. Louis, MO, USA). The islet tissue clusters (50–500 μm) isolated using this method were allowed to mature into islet cell clusters during in vitro culture at 37°C, 5% CO2 first in Recovery Maturation Media (University of California Irvine, Irvine, CA, USA) supplemented with 215 mM aprotinin (Sigma-Aldrich), 0.5 mM Pefabloc (Sigma-Aldrich), 417 mM dornase alfa (Genentech, San Francisco, CA, USA), and 10% porcine serum (Omega Scientific, Tarzana, CA, USA), and 48 h later, in a novel maturation media (University of California Irvine, Irvine, CA, USA) supplemented with 10% porcine serum (Omega Scientific, Tarzana, CA, USA) as outlined previously 13 .
Yield, Viability and Function Determination
Juvenile pigs
Islet yield was calculated from the previous 12 juvenile pig islet isolations. At the end of the 7-day culture period, islet quality control assessment was performed. Islet count and IEQ were determined by staining a 100-μL aliquot with 1 mL dithizone (MP Biomedicals, Santa Ana, CA, USA), and observing islets at 25x on a standard stereomicroscope (Max Erb, Santa Ynez, CA, USA) containing a 10x eye piece graticule. Islet viability was analyzed using fluorescein diacetate (FDA) and propidium iodide (PI) (Invitrogen, Carlsbad, CA, USA), imaged using fluorescence microscopy (Nikon LSM510; Thornwood, NY, USA), and quantified with a Microplate reader (Tecan Infinite F200, Magellan V7; Tecan, Männedorf, Switzerland).
Islet function was determined using glucose-stimulated insulin release; 100–150 porcine islets were incubated for 1 h, in corresponding order, in low glucose (2.8 mM), high glucose (28 mM), high glucose plus 3-isobutyl-1-methylxanthine (28 mM, 50 μM), and then low glucose (
Adult pigs
Average yield and viability of adult islets were estimated from the literature. Four studies composed of nine subgroups and a total of 131 adult pigs were analyzed (Table 1) 4–5,14–15 . One study reported viability for each of its three subgroups, and average viability was calculated from this study. SI, as measured by glucose-stimulated insulin release, was reported in two studies.
Average Yield, Viability, Stimulation Index (SI), and Cost of Adult Porcine Islets.
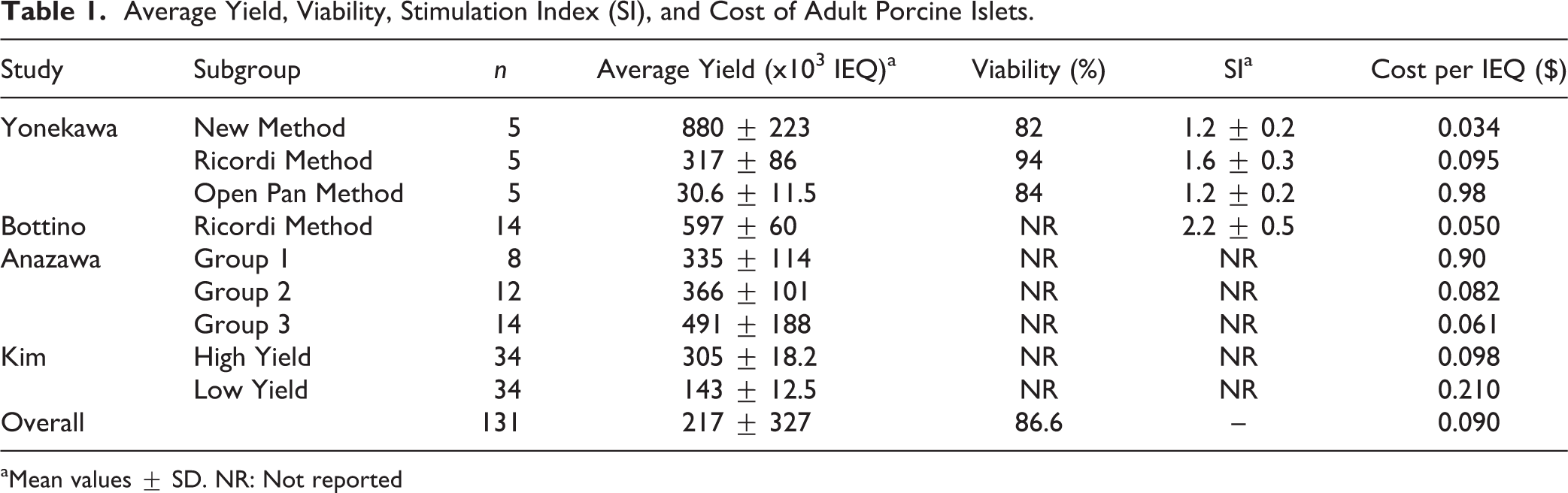
aMean values ± SD. NR: Not reported
Neonatal pigs
Average yield and viability of neonatal islets were estimated from the literature. One study comprised of three subgroups and 136 pigs was analyzed 16 . Average SI, as measured by glucose-stimulated insulin release, was reported for each subgroup (Table 2). Viability data was not available and thus not considered for neonatal pigs in this paper.
Average Yield and Stimulation Index of Neonatal Porcine Islets.

aMean values ± SD. NR: Not reported
Cost Determination
For neonatal, preweaned juvenile and adult pigs, islet isolation cost was divided into three categories. Animal source, husbandry, and veterinary costs were grouped together as “Husbandry”, surgical procurement and organ preservation including operating room costs were grouped as “Surgical”, and isolation and tissue culture costs were grouped as “Isolation”. All categories included applicable materials as well as labor cost. Isolation and surgical costs were calculated using values provided by the University of California, Irvine (CA, USA). An institutional tax of 29.8% was applied across all categories (Table 3).
Neonatal, Juvenile, and Adult Islet Isolation—Complete Costs Per Pig.

Data Analysis
The primary statistical endpoints of this study were islet yield, viability, and SI of each population. Pooled means and standard deviations (SDs) were appropriately calculated when the given population was homogeneous. For instance, data for juvenile pigs was obtained from a single protocol and population, and data for neonatal pigs was obtained from a single study. Thus, these populations were each treated as homogeneous. For juvenile pigs, cost per islet was determined using this pooled mean and the calculated cost of juvenile islet procurement. Similarly, viability as well as mean and SD of SI was determined from pooled data for the 12 trials of this study.
For neonatal pigs, a pooled mean and SD was calculated under the assumption that subgroups from the single analyzed study originated from a homogeneous population. Cost per islet was determined using this pooled mean and the calculated cost of neonatal islet procurement. Similarly, mean and SD of SI was determined from pooled data for the three subgroups of this study. Viability data was not available.
Data regarding islet yield in adult porcine pancreata was extracted from previous studies 4–5,14–15 . However, because studies were conducted under differing conditions and samples were not homogenous, pooled means and SDs are limited in their usefulness of analyzing this data 17 . Instead, each study was evaluated individually and the cost per islet was determined based on the number of islets procured in each study and the calculated cost of adult porcine pancreata (Table 1). For illustrative purposes, and in order to broadly compare average islet yields in adult versus juvenile and neonatal pigs, a simple weighted average of islet yield was calculated to account for differences in sample size among the included studies. Viability was calculated using simple weighted averages and pooled SDs, as these values arose from a single study and thus the population was deemed to be homogenous. SI was reported in two studies, and thus pooling was not appropriate. Indices are reported in Table 1.
Results
Yield, Viability and Function of Islets in Juvenile, Adult, and Neonatal Pigs
Islet yield after 7 days of in vitro culture was 30.4 ± 1.20 x 103 IEQ per juvenile pig, with average viability of 87.5%. Glucose-stimulated insulin response gave an average value of 1.77 ± 0.49.
Values from nine protocols and 131 animals were used to determine the range of islet yields from adult porcine pancreata. Islet yield averages ranged from 30 x 103 to 880 x 103 IEQ per pig with viabilities ranging from 82% to 94%, and SIs ranging from 1.2 to 2.2 (Table 1). For illustrative purposes, average islet yield was determined to be 333 ± 129 x 103 IEQ per adult pig. Average reported viability was 86.7% (Table 1).
Values from one protocol (three subgroups) and 136 animals were used to determine the average islet yield from neonatal porcine pancreata. Average islet yield was determined to be 38.1 ± 3.0 x 103 IEQ per adult pig with an average reported viability of 87.5%. Average SI was calculated to be 4.7 ± 0.58 (Table 2).
Cost Comparison of Juvenile, Adult, and Neonatal Pig Islet Procurement
There were no differences in isolation or surgical costs for neonatal or juvenile pigs at our institution. Costs of adult pig husbandry were US $9958 (costs for raising pigs raised to 2 years of age) compared with approximately $96 for neonatal and $576 for 18- to 24-day-old pre-weaned juvenile pigs (Table 3). Surgical costs were also moderately less expensive for neonatal and juvenile pigs ($30.50 per animal) compared with adult pigs ($228.77 per animal). Notably, up to 12–15 juvenile pig pancreases can be recovered in the same time it would require to complete 1 adult pig pancreas procurement (1.5–2 h). Isolation costs for neonatal and juvenile pigs were calculated at $642.76 per isolation, including 7 days in vitro tissue culture, compared with approximately $16,946 for adult pig isolation including gradient purification and in vitro tissue culture. Therefore, the total cost of neonatal islet isolation was calculated as $769 per pancreas, juvenile islet isolation as $1249 per pancreas, and adult islet isolation as $27,133 per pancreas. The price per IEQ was therefore determined to be $0.02 for neonatal pigs and $0.04 for juvenile pigs. Price per IEQ for adult pigs ranged from $0.03 to nearly $1.00. An average price per IEQ of $0.09 was estimated for adult pigs (Table 4, Supplemental Table 5).
Average Yield, Price, Viability and Stimulation Index by Age of Donor.

aMean values ± SD.
Discussion and Conclusion
Study Limitations
There were a few notable limitations to this study owing to its retrospective nature. Because our laboratory does not isolate islets from adult or neonatal porcine pancreata, data regarding these subgroups had to be estimated from the literature. Although we were able to find data from multiple studies examining adult porcine islet yields, there were few studies that reported this data in neonatal piglets, and thus sample size was limited in this group. Furthermore, although pooling multiple studies on adult islet isolation allowed for a more complete analysis of islet yields in this population, it also limits the efficacy of a pooled mean and SD in this group, as the populations of these studies cannot be assumed to be identical. Since the studies all sampled adult pigs, and we were not concerned with specific method of isolation, we were able to generate a pooled mean for limited illustrative purposes. However, the cost per IEQ of each individual study is a more exact measure of this value (Table 1).
Cost Effectiveness and Scalability of Juvenile, Neonatal, and Adult Porcine Islets
In this study, we evaluated the cost-efficiency and scalability of neonatal, juvenile, and adult porcine islets, accounting for husbandry, surgery, and isolation costs for each group. Our findings indicate that neonatal ($0.02/IEQ) and juvenile ($0.04/IEQ) porcine islets are more cost-effective than adult porcine islets ($0.09/IEQ, range $0.03 to $0.98). Looking solely at cost, juvenile porcine islets cost less than half of the price of adult islets while having nearly identical viability and SI measurements. Neonatal porcine islets had more than double the SI of juvenile and adult islets at only a fraction of the cost. However, there was limited literature that specified neonatal porcine islet yield, and no study reported viability in the data we gathered. Nevertheless, previous studies have confirmed higher stimulation indices in neonatal porcine islets versus adult porcine islets, attributing their findings to the lower basal secretion of neonatal islets and the minimal damage sustained during the less aggressive isolation procedure. The 7-day culture of neonatal and juvenile islets also may allow for better cell recovery and function, since the adult islets are not cultured prior to secretion assays 18–19 . Moreover, while islet yields in young pigs have been shown to increase throughout a culture period, likely due to further differentiation of duct cells into islet cells, studies have shown a 30–50% decrease in yield after culture of adult islets 14,19 . Furthermore, studies suggest that neonatal islets continue to secrete insulin beyond the time that the secretory function of adult islets begins to decline 20 .
The reproducibility and scalability of juvenile and neonatal porcine islet isolations is superior to that of adult porcine islets. Adult pig isolations had highly variable total islet yields (Table 1), whereas neonatal and juvenile pig isolations had relatively consistent total islet yields, thereby indicating their increased reproducibility. Part of the variability of the adult porcine islet yields can be attributed to differing isolation conditions; the success of such adult isolations have been shown to depend on morphological screening of the pancreas, warm ischemia time, and endotoxin content, among other factors 21 . In terms of scalability, the relative simplicity of the juvenile and neonatal porcine islet isolation procedure maximizes islet yield per hour of labor, since the procedure essentially consists of removing the pancreas and chopping it into small fragments, followed by enzymatic digestion and subsequent cell culture. However, adult porcine islet isolation is much more complicated. In short, one must first inject collagenase selectively through the pancreatic duct and later break down fibrous tissue by placing the pancreas within a Ricordi chamber and continue to vigorously shake it while also being careful not to damage the islets 22 . This aggressive isolation procedure can limit islet yield, function, and viability through islet destruction. Recent studies have begun using an Oxford chamber—a digestion and filtration chamber similar to the Ricordi chamber—because it results in reduced tissue destruction and greater islet yields 23 . Nevertheless, the use of both chambers requires more training and a more optimized protocol due to the many variables that can alter isolation success, like collagenase injection pressure, digestion time, and chamber temperature 24,25 .
As a result of the relative ease of the juvenile and neonatal pig isolations, under our juvenile pancreatic isolation procedure, as many as 24 pancreases have been procured over a 4-h period, illustrating the improved scalability of the procedure when compared with adult pig pancreas isolations. Even at this heightened pace, juvenile islet yields were similar to results obtained with standard processing of two to four pancreases over the same period of time. Juvenile islet isolation also requires less extensive training and decreased labor hours, taking 20 h to isolate from four juvenile pigs, but 58 h to isolate from 1 adult pig. These islet isolation times were determined from our isolation data of juvenile pigs and data provided by the University of California, Irvine. This disparity was also accounted for in our cost calculation per IEQ for each porcine islet group.
Our findings indicate that isolating neonatal and juvenile porcine islets is more cost-effective and scalable than isolating adult porcine islets. Neonatal and juvenile porcine islets demonstrated similar or superior viability and SI values when compared with adult porcine islets. Furthermore, the adult porcine islet isolation procedure is more difficult and labor-intensive to successfully conduct and produce consistent results, making the juvenile and neonatal porcine islet isolations more scalable for future experimentation.
Supplemental Material
Supplemental_Table_1 - Cost and Scalability Analysis of Porcine Islet Isolation for Islet Transplantation: Comparison of Juvenile, Neonatal and Adult Pigs
Supplemental_Table_1 for Cost and Scalability Analysis of Porcine Islet Isolation for Islet Transplantation: Comparison of Juvenile, Neonatal and Adult Pigs by Rachel Vanderschelden, Mayilone Sathialingam, Michael Alexander and Jonathan R. T. Lakey in Cell Transplantation
Footnotes
Ethics Approval
All procedures in this study were conducted in accordance with University of California Irvine Institute of Animal Care and Use Committee, approved protocol #AUP-17-129.
Statement of Human and Animal Rights
All procedures were performed according to protocols approved by the University of California Irvine Institute of Animal Care and Use Committee.
Statement of Informed Consent
Informed consent is not applicable because there are no human subjects in this study.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.
Supplemental Material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.