
Abstract
Pancreatic islets isolated for transplantation are disconnected from their vascular supply and need to establish a new functional network posttransplantation. Due to poor revascularization, prevailing hypoxia with correlating increased apoptosis rates in experimental studies can be observed for months posttransplantation. Endothelial progenitor cells (EPCs) are bone marrow–derived cells that promote neovascularization. The present study tested the hypothesis that EPCs, isolated from human umbilical cord blood, could be coated to human islet surfaces and be used to promote islet vascular engraftment. Control or EPC bioengineered human islets were transplanted into the renal subcapsular space of nonobese diabetic/severe combined immunodeficiency mice. Four weeks posttransplantation, graft blood perfusion and oxygen tension were measured using laser Doppler flowmetry and Clark microelectrodes, respectively. Vessel functionality was also assessed by in vivo confocal imaging. The vascular density and the respective contribution of human and recipient endothelium were assessed immunohistochemically by staining for human and mouse CD31. Islet grafts with EPCs had substantially higher blood perfusion and oxygen tension than control transplants. Furthermore, analysis of the vascular network of the grafts revealed that grafts containing EPC bioengineered islets had a superior vascular density compared with control grafts, with functional chimeric blood vessels. We conclude that a simple procedure of surface coating with EPCs provides a possibility to improve the vascular engraftment of transplanted human islets. Established protocols are also easily applicable for intraportal islet transplantation in order to obtain a novel directed cellular therapy at the site of implantation in the liver.
Introduction
The metabolically active pancreatic islets are highly vascularized, with a blood perfusion regulated to match the different metabolic demands for oxygen and nutrient supply of the endocrine cells 1 . The islet vasculature also supports β cell growth, differentiation, and function by paracrine interactions 2 –6 and provides drainage of secreted hormones to the central circulation.
Pancreatic islets isolated for transplantation are disconnected from their vascular supply and need to establish a new functional network posttransplantation. Due to poor revascularization, prevailing hypoxia with correlating increased apoptosis rates in experimental studies can be observed for months posttransplantation, not only at the renal subcapsular site 7,8 but also at the clinically used intraportal site 9,10 . It is therefore desirable to find a therapeutic alternative that allows for better revascularization shortly posttransplantation.
Endothelial progenitor cells (EPCs) were first described in 1997 by Asahara et al. 11 . These proangiogenic cells are recruited from the bone marrow in response to a vascular injury and contribute to neovascularization by incorporation into the damaged vasculature 12 –14 , as well as through the release of different angiogenic and proinflammatory factors 15 –18 . Peripheral mobilization of EPCs and other bone marrow–derived cells by granulocyte-macrophage colony-stimulating factor were previously shown to enhance pancreatic islet revascularization and function after intraportal transplantation 19 . However, this technique has not been implemented clinically, possibly in fear of evoking an immune rejection of the graft. More recently, EPCs have instead been mixed together with rodent 20,21 or porcine 22 islets and shown to increase islet vascular density and β cell function posttransplantation in rodents.
The present study tested the possibility to improve engraftment of transplanted human islets by EPCs. In order to establish a clinically useful protocol, which is also applicable to the intraportal site, a simple coating procedure was developed, where EPCs attached to the islet surface. Moreover, in order to exclude that the previously observed increased revascularization by EPCs merely reflected dysfunctional blood vessels, in vivo imaging, blood perfusion, and oxygen tension measurement techniques were applied for the first time to quantify their functional relevance.
Materials and Methods
Animals
Male nonobese diabetic/severe combined immunodeficiency mice (Taconic M&B, Ejby, Denmark) weighing 25 to 30 g were used in all experiments. The animals were housed under standardized conditions (12 h light/12 h dark cycle) and had free access to water and pelleted food.
Islet Isolation and Culture
Human islets were obtained from the Nordic Network for Islet Isolation. Islets were isolated from 11 brain-dead donors (8 males, 3 females; see Table 1 for donor characteristics) and transplanted within 6 to 14 d after isolation. The human islets were cultured at 37 °C and 5% CO2 in Connaught Medical Research Laboratories (CMRL) 1066 media (Gibco, Grand Island, NY, USA) containing 10% (vol/vol) fetal calf serum (Sigma-Aldrich, St. Louis, MO, USA), 50 mmol/L
Characteristics of Donors Used in This Study.
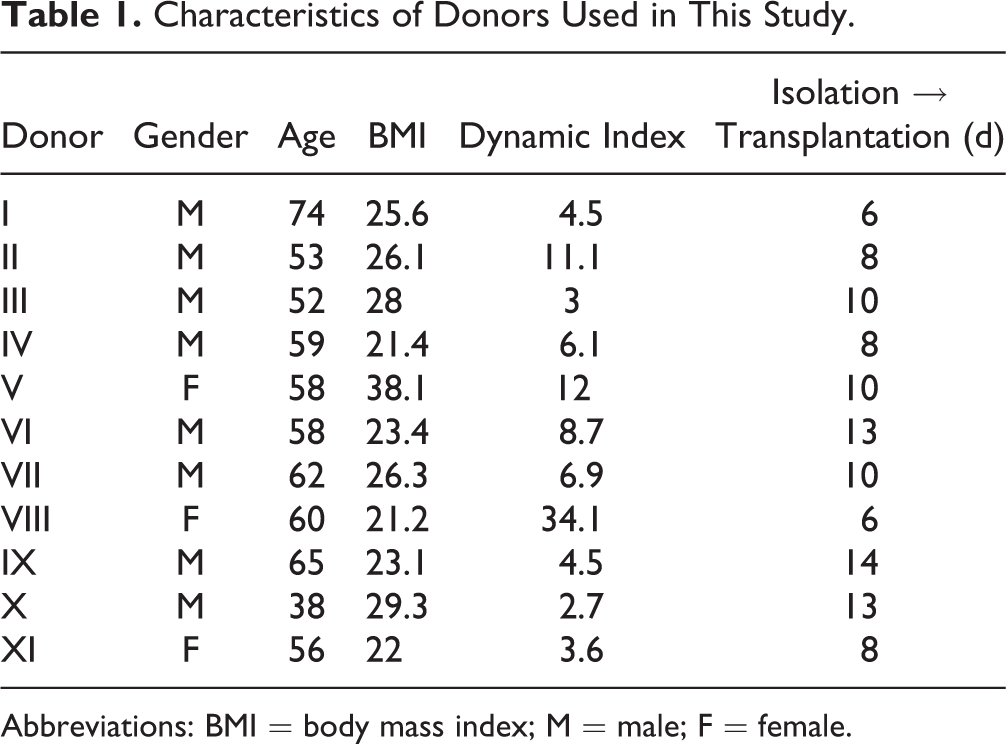
Abbreviation: BMI = body mass index; M = male; F = female.
EPC Culture and Coating Procedure
EPCs from human umbilical cord blood (AMS Biotechnology, Abingdon, United Kingdom) were used at passages 6 and 7. The obtained cells had been characterized by their expression of CD146, CD34, vascular endothelial growth factor receptor 2, von Willebrand factor, low-density lipoprotein uptake, and lectin binding. The cells were defined as late EPCs, since they did not express CD133. Their functional capacity was confirmed by tube formation and migration assays by AMS Biotechnology. Cells in our lab were cultured in T75 flasks (Corning Life Sciences, Corning, NY, USA), with endothelial progenitor outgrowth cell (EPOC) media (AMS Biotechnology). In order to coat EPCs to the islet surface prior to transplantation, the islets were incubated at 37 °C with 10,000 EPCs per islet for 1 h under gentle shaking. This number of EPCs per islet was chosen based on pilot experiments, where full coverage of the human islet surfaces was observed within 1 h of coincubation.
Cell Tracking
In order to determine the fate of the EPCs, 1 batch of cells were labeled using Qtracker 625 cell-labeling kit (Life Technologies, Carlsbad, CA, USA) prior to transplantation.
Islet Transplantation
Animals were anesthetized by an intraperitoneal injection of Avertin (0.015 mL/g body weight of a 2.5% (vol/vol) solution of 10 g 97% (vol/vol) 2,2,2-tribromoethanol [Sigma-Aldrich] in 10 mL 2-methyl-2-butanol [Kemila, Stockholm, Sweden]]. A small incision was made at the left subcostal flank, exposing the kidney. Two hundred control or EPC bioengineered human islets were transplanted into the renal subcapsular space. In separate experiments, control and EPC bioengineered human islets were instead transplanted into the abdominal external oblique muscle (for technique, see the study by Christoffersson et al. 23 ).
Blood Flow and Oxygen Tension
One-month posttransplantation, the mice were again anesthetized by Avertin and maintained at body temperature on a surgical table. A polyethylene catheter was inserted into the right carotid artery and thereafter connected to a blood pressure transducer (ADInstruments, Dunedin, New Zealand). Another incision was made at the left subcostal flank. The kidney was freed from its surrounding tissues and immobilized in a plastic cup, with a clear view of the graft. Graft and kidney blood perfusion were measured using laser Doppler flowmetry with a needle probe (Transonic BLF 21 Series, probe diameter 1.2 mm; Transonic, Ithaca, NY, USA). Clark microelectrodes (external tip diameter = 2 to 5 µm; Unisense, Aarhus, Denmark) were used to measure the oxygen tension of the graft as well as the kidney cortex. The same microelectrode was used for all experiments. Six to 10 measurements were performed in each graft or kidney, and the calculated mean in each organ was considered to be one experiment. All measurements were performed according to previously established protocols 24 .
Ten minutes prior to sacrifice, some animals received an intrajugular injection with 100 μg soybean agglutinin lectin (Life Technologies) to study perfused, and thus functional, vessels within the grafts. The kidney was thereafter removed and snap-frozen in liquid nitrogen.
In Vivo Imaging
Mice transplanted with control or EPC bioengineered human islets to striated muscle in the abdominal wall were at 2 wk posttransplantation anesthetized by isoflurane and maintained at body temperature on a surgical table. An intravenous injection of Alexa Fluor 488-conjugated rat anti-mouse CD31 (BioRad, Hercules, CA, USA) and APC-conjugated mouse anti-human CD31 (eBioscience, San Diego, CA, USA) was performed. The antibodies were allowed to circulate for a minimum of 5 min prior to imaging. An incision was made to the skin to expose the graft, and the muscle was immobilized for imaging using a vacuum window 25 . A Zeiss LSM 5 LIVE microscope (Carl Zeiss AG, Oberkochen, Germany), with a Plan-Apochromat 20×/0.8 objective, was used for all in vivo imaging.
Immunohistochemistry
Cryosections were made with a thickness of 8 µm. Sections were air-dried and thereafter fixed in cold (4 °C) acetone for 5 min. Avidin block solution (Vector Laboratories, Burlingame, CA, USA) was added to each slide for 20 min, followed by biotin block solution (Vector Laboratories) for an additional 20 min. The sections were thereafter covered in Tris-NaCl-blocking (TNB) buffer (PerkinElmer, Waltham, MA, USA) for 30 min.
The slides were incubated with primary antibodies (rabbit anti-human IA2 [anti-Protein Tyrosine Phosphatase, Receptor Type N {PTPRN}, 1:200 in TNB buffer; Atlas antibodies, Stockholm, Sweden]), biotin conjugated mouse anti-human CD31 (1:300 in TNB buffer; eBioscience), and rat anti-mouse CD31 (Clone ER-MP12, 1:100 in TNB buffer; AbD Serotec, Kidlington, UK) at 4 °C overnight. Slides were washed 3 times with washing buffer (Dako, Glostrup, Denmark), followed by a 60 min incubation at room temperature with the secondary antibodies (Alexa Fluor 555 goat anti-rat [1:200 in TNB buffer; Life Technologies] and Alexa Fluor 647 donkey anti-rabbit [1:200 in TNB buffer; Jackson ImmunoResearch Laboratories, West Grove, PA, USA]). The secondary antibodies were removed and slides were washed 3 times with washing buffer. Streptavidin-HRP (1:150 in TNB buffer; PerkinElmer) was added for 30 min, followed by washing and incubation with Fluorophore Tyramide (1:80 in amplification buffer; PerkinElmer) for 8 min. The nuclei were stained with Hoechst (1:10,000 in PBS; Life Technologies) for 5 min in room temperature. The slides were dried and mounted with Fluoromount-G (SouthernBiotech, Birmingham, AL, USA).
Confocal Imaging and Islet Vascular Density Assessment
Confocal imaging was performed using a laser scanning confocal microscope Zeiss LSM 780 (Carl Zeiss AG) with a Plan-Apochromat 20×/0.8 M27 objective. Zeiss Zen 2012 Blue edition software, version 1.1.1.0 (Carl Zeiss AG) was used for all image analyses. Islet vascular density was assessed by measuring human and mouse CD31-positive areas within IA2-positive areas (Islet Antigen-2 [IA2] is a ubiquitous marker for islet endocrine cells 26 ).
Statistical Analysis
All data are presented means ± Standard error of the mean (SEM). Paired and unpaired Student’s t-test was used for comparison between control and bioengineered grafts. For all comparisons, a P value of <0.05 was considered significant. All statistical analyses were carried out using GraphPad Prism Version 6.0 (GraphPad Software, San Diego, CA, USA).
Results
EPC Bioengineered Human Islets
By incubating islets together with EPCs for 1 h under gentle shaking, the surface of the islets became covered with EPCs. After prolonged culture (48 h) of some of the islets, the EPCs were still attached to the surface, indicating a strong cell-to-surface binding (Fig. 1A and B).

Human islets coated with endothelial progenitor cells (EPCs). A majority of the islets (yellow = insulin) that had been coated with EPCs (green = human CD31, indicated by white arrows) still had a layer of EPCs covering the islet surface, indicating a strong cell-to-surface binding 48 h later (A and B). Furthermore, these images also show remnant donor endothelial cells (also in green, indicated by red arrows), confirming that human islet endothelial cells survive in culture. All scale bars correspond to 50 µm.
Human Islets Contained Endothelial Cells at the Time of Transplantation
The images of human islets in vitro showed that many of these islets still contained remnant endothelial cells after 8 to 16 d of culture (Fig. 1A and B). There was no difference in remnant donor endothelial cells between the groups at the time of transplantation.
Human Islets Coated with EPCs Had a Higher Vascular Density
EPC bioengineered grafts at the renal subcapsular site had an increased islet vascular density when compared with control transplants at 1 mo posttransplantation (Fig. 2A to E; n = 8 in both groups). This reflected a doubled area of blood vessels of both mouse (recipient) and human origin.

Vascular density of control and endothelial progenitor cell (EPC) bioengineered grafts. Vascularization of control (A and B) and EPC-coated human islets (C and D) at 1 mo posttransplantation. Contribution of ingrowing recipient (mouse) endothelial cells was evaluated by mouse CD31 staining (red), whereas human endothelial cell contribution was assessed by human CD31 staining (green). The endocrine mass was visualized by the ubiquitous islet endocrine marker IA2 (yellow) in (A) and (C). (E) Quantification of vascular density and the respective contribution of mouse and human endothelial cells in the grafts of control (closed bars) and EPC-coated human islets (open bars) are shown (n = 8 in both groups). (F) At 1 mo posttransplantation, many of the transplanted EPCs (yellow = Qtracker 625) were incorporated (indicated by white arrows) into functional vessels (green = bound soybean agglutinin lectin). Scale bars correspond to 200 µm (A), 100 µm (C), and 50 µm (F). All values are given as means ± standard error of the mean for 8 transplanted animals in each group. *P < 0.05 when compared to control islet grafts.
EPCs Were Incorporated into Functional Blood Vessels
By labeling EPCs prior to transplantation, these cells could be traced and imaged within the renal subcapsular grafts. The observed overlap between labeled EPCs and lectin confirmed that EPCs were incorporated into functional blood vessels at 1 mo posttransplantation (Fig. 2F; n = 3).
Grafts Containing EPCs Had Significantly Higher Blood Perfusion and Oxygen Tension
In order to assess the functionality of the newly formed graft vessels, blood perfusion and oxygen tension were measured within the human islet grafts and surrounding kidney tissue. The blood perfusion in EPC bioengineered grafts (n = 8) was more than double that of control grafts (n = 9) 1 mo posttransplantation (Fig. 3A). Moreover, the EPC containing grafts had a 3-fold increase in oxygen tension when compared with control grafts (Fig. 3B). Meanwhile, the blood perfusion and oxygen tension of the kidney cortex did not differ between the groups (Fig. 3C and D).

Blood perfusion and oxygen tension. Blood perfusion (A) and oxygen tension (B) in 1-mo-old transplants of control (closed bars; n = 9) or endothelial progenitor cell (EPC)-coated human islets (open bars; n = 8). Blood perfusion and oxygen tension were also recorded in the adjacent renal cortex of the same animals (C and D, respectively). All values are given as means ± standard error of the mean for 8 to 9 transplanted animals in each group. ***P < 0.001 when compared to control islet grafts.
Functional Chimeric Vessels Were Present in EPC Bioengineered Grafts
To further assess the functionality of the newly formed vessels, islets were transplanted to the striated muscle of the abdominal wall in 7 animals. This transplantation site enabled for easy visualization of the islet grafts within the living animals. This imaging confirmed that the donor (human) and recipient (mouse) vessels connect within the grafts to form functional (blood perfused) vessels (Fig. 4A and B). The EPC bioengineered grafts also contained functional chimeric (Fig. 4B and C) blood vessels (i.e., vessels of both human and mouse origin), further supporting the finding that EPCs incorporate into ingrowing blood vessels.

In vivo imaging of control and endothelial progenitor cell (EPC) bioengineered grafts. Human islets were transplanted to the striated muscle of the abdominal wall and visualized by in vivo microscopy 2 wks later. By in vivo imaging, it was confirmed that human (red = human CD31) and mouse (green = mouse CD31) vessels connect within the grafts (indicated by white arrows) to create functional blood vessels (A and B). The EPC bioengineered grafts also contained blood-perfused chimeric vessels (indicated by yellow arrows), further supporting the finding that EPCs incorporate into functional blood vessels (B and C). All scale bars correspond to 50 µm.
Discussion
Poor engraftment of the implanted tissue constitutes a major challenge for successful clinical islet transplantation. In the present study, we describe a simple procedure of surface-coating islets with EPCs prior to transplantation and show this to be a highly effective mean to improve the revascularization of human islet grafts. The formed blood vessels were found highly functional and more than doubled the blood perfusion and oxygen tension in the tissue at 1 mo posttransplantation.
Previous studies of transplanted murine islets have shown that islet endothelial cells disappear within the first days during islet culture 27,28 . In contrast, the results of the present study indicated that human islet endothelial cells remain even after prolonged culture of the human islets, thereby providing a possibility for these endothelial cells to participate in the newly formed vascular network of the islets posttransplantation. The reason for this striking difference is presently obscure. However, besides the species difference, studies of murine islet endothelial cells in culture indicate that they are highly susceptible to glucose toxicity and that their apoptosis rates could substantially be decreased by lowering the glucose concentration during culture 29 . Similarly, glucose has been shown to inhibit angiogenesis of isolated human islets 30 . It is noteworthy that the clinical protocols, as well as the present study, used a lower glucose concentration (5.6 mmol/L) for islet culture than that commonly applied for rodent islet cultures (11 mmol/L) 28 .
The EPCs used in the present study did not express CD133 and are thus considered to be late EPCs, also known as EPOCs 31 , in contrast to the originally described early EPCs obtained from circulating adult peripheral blood nuclear cells 11 . It has been proposed that early and late EPCs contribute to the neovascularization in different ways; early EPCs are suggested to release angiogenic cytokines that are thought to attract more endothelial cells to the site of injury, while late EPCs mainly provide the area with sufficient numbers of endothelial cells for the neovasculogenesis to occur and do so by differentiating into mature endothelial cells 12 . Nevertheless, also, late EPCs produce a multitude of angiogenetic factors, including matrix metalloproteinase 2 and vascular endothelial growth factor (VEGF) 17 , and cord blood–derived late EPCs have recently been observed to promote angiogenesis by mere paracrine factors 32 . Indeed, the higher number of blood vessels in EPC bioengineered islet grafts in the present study was observed to result not only from an incorporation of the EPCs into the capillary networks but also from an increased ingrowth of recipient vessels (mouse) and proliferation of residual human endothelial cells. Previous studies indicate that EPCs may trigger angiogenesis in rodent and porcine islets by either stimulating the VEGF-A expression in the transplanted β cells 22 or the secretion of VEGF-A directly from the EPCs 20 . Another mechanism may be that microvesicles from EPCs transfer RNA stimulating angiogenesis in tissue 33 . Whereas Oh et al. 20 , similar to us, reported that both donor and recipient endothelium, as well as the transplanted EPCs, contributed to the newly formed blood vessels, the inclusion of EPCs in blood vessels was not observed by Kang and colleagues 22 . Discrepancies between studies are likely explained by different characteristics of the transplanted EPCs, but perhaps also species differences. Importantly, in the present study of human islets, we show that the grafts with EPCs acquire a much higher vascular density than control grafts, and that inclusion of EPCs does not only enhance the speed of revascularization as reported previously 22 .
One month posttransplantation, the area of blood vessels in islet grafts containing EPCs was almost doubled when compared with control islet grafts. Although this has also been reported for murine islets with murine EPCs 20 , those and our findings could have been explained by expansion of transplanted EPCs or remnant endothelial cells in the tissue, without the formation of functional perfused blood vessels. We therefore investigated the formed blood vessels for functionality by different techniques. Firstly, that the vascular structures, including those with incorporated EPCs and remnant endothelium, were perfused and were confirmed by both intravenous lectin infusion and in vivo imaging. Secondly, the recordings of blood perfusion by laser Doppler flowmetry showed that the blood perfusion levels in the grafts of EPC-coated islets were much higher than in control grafts and similar to those in the highly blood-perfused renal cortex. In this context, it may be noted that at least in rodents, the blood perfusion of native islets is similar to that in the renal cortex 1,34 . Thirdly, the oxygen tension levels in the grafts of EPC-coated islets was restored to that of native rodent islets 35 , suggesting optimal oxygen delivery to the transplanted β cells.
In the chosen study design, the islets were transplanted beneath the kidney capsule and to striated muscle to enable functional evaluation of the formed vasculature by in vivo microscopy and direct blood perfusion and oxygen tension measurements. However, although not tested in the present study, the established protocols are also easily applicable for intraportal islet transplantation in order to obtain a novel directed cellular therapy at the site of implantation in the liver. Normoglycemic and not hyperglycemic recipient mice were used in the study, which may have influenced the results. However, in the clinical situation, efforts are applied to obtain strict metabolic control posttransplantation, and, at least for the revascularization of murine islets, the process is independent of the glycemic state of the recipient 36 . Native EPCs of patients with diabetes are essentially defective in both numbers and function 37,38 , which provide an important rationale for their substitution in islet transplantation, although they, at least in animal models, in lower numbers may home to transplanted islets 39 . EPCs for bioengineering pancreatic islets can easily be obtained from peripheral blood of adult patients or, as in the present study, from umbilical cord blood. Heightened HLA sensitization may occur if including EPCs with islets, but this may be solved by using islet donor- or recipient-derived EPCs.
Footnotes
Authors’ Note
Prior publication of this study is in abstract form. Part of this article was presented as an abstract at the International Pancreas and Islet Transplantation Association Meeting in Monterey, CA, in 2013.
Acknowledgments
The skilled technical assistance of Dr. Zhanchun Li and Mrs. Lisbeth Sagulin, Department of Medical Cell Biology, Uppsala University, is gratefully acknowledged. We are also grateful to Professor Asahara and Tokai University for providing the opportunity for Liza Grapensparr to visit and learn EPC culture and expansion techniques as well as to the Nordic Network for Clinical Islet Transplantation for their generous support with human islet preparations.
Ethical Approval
All animal experiments were approved by Uppsala’s Ethical Committee on Animal Experiments (C124/15). All experiments involving human islets were approved by the Regional Ethical Review Board in Uppsala (Ups 02/290).
Statement of Human and Animal Rights
All experiments were conducted in accordance with Swedish human and animal protection laws.
Statement of Informed Consent
This study made use of human islets from the pancreas of deceased brain-dead donors. The organs were provided to the Nordic Network for Clinical Islet Transplantation for clinical or research use according to the Swedish laws for organ donation.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This work was supported by the Swedish Research Council, EXODIAB, StemTherapy, Torsten Söderberg Foundation, the EFSD/JDRF/Novo Nordisk Programme 2012, the Swedish Diabetes Association, the Swedish Child Diabetes Foundation, Diabetes Wellness Sverige, the Novo Nordisk Foundation, Olle Engqvist Byggmästare Fund, and AFA.