
Abstract
Idiopathic Parkinson’s disease (PD) is a progressive neurodegenerative disorder, clinically manifested by cardinal motor symptoms including tremor at rest, bradykinesia, and muscle rigidity. Transplantation of dopaminergic (DAergic) neurons is an experimental therapy for PD, however, it is limited by suboptimal integration and low survival of grafts. Pretreatment of donor tissue may offer a strategy to improve properties of transplanted DAergic neurons and thereby clinical outcome. We have previously shown that a combination of neurotrophin-4/5 (NT-4/5) and glial cell line-derived neurotrophic factor (GDNF) demonstrated additive effects on rat ventral mesencephalic (VM) tissue. The present study investigated the effects of NT-4/5 and GDNF as single factors, or in combination on DAergic neurons, in organotypic explant cultures of fetal human ventral mesencephalon. For that purpose, free-floating roller-tube cultures were prepared from VM and the equally sized pieces grown for 1 week in the presence or absence of neurotrophic factors. Both neurotrophic factors increased dopamine content in the culture medium and in the number of tyrosine hydroxylase immunoreactive neurons, most prominently after combined GDNF + NT-4/5 treatment. Culture volumes did not differ between groups while content of lactate dehydrogenase in the culture medium was moderately reduced in all treated groups. In conclusion, we identified that a combination of GDNF and NT-4/5 robustly promoted differentiation and survival of human fetal VM DAergic neurons, an observation with potential promising impact for cell replacement approaches in PD.
Introduction
Progressive loss of dopaminergic (DAergic) neurons in the substantia nigra pars compacta is critically implicated for the motor symptoms observed in patients suffering from Parkinson’s disease (PD). While drug therapies provide an effective means to substitute for the missing intrastriatal dopamine, long-term treatment often goes together with severe side effects. In this context, cell replacement strategies in PD have emerged as an experimental therapeutic concept. Thereby, the transplanted cells serve as a biological pump releasing dopamine. Human fetal nigral tissue can be transplanted safely, and it has been shown that grafts survived and reinnervated the host striatum of PD patients 1 –4 . This approach, however, is limited by suboptimal survival and differentiation of grafted DAergic neurons 1 .
Hence, alternative sources of transplantable DAergic neurons have been investigated including induced pluripotent stem cells (iPSC) and embryonic stem cells (ESC), but also their capacity to reinnervate the diseased striatum in rodents was found to be critical 5 . A number of neurotrophic factors have been described to increase the survival and growth of DAergic neurons including the glial cell line–derived neurotrophic factor (GDNF) and neurotrophin-4/5 (NT-4/5) 6 . GDNF belongs to the transforming growth factor-β (TGF- β) superfamily and mediates cellular responses by a multicomponent receptor complex composed of the rearranged during transfection (RET) proto-oncogene and GDNF family receptor α. In cultures of fetal midbrain, GDNF promotes survival and differentiation of DAergic neurons and increases high-affinity dopamine uptake 7,8 . NT-4/5 is a member of the NT family. Upon tropomyosin receptor kinase B (TrkB) receptor binding NT-4/5 initiates intracellular signaling cascades including the Rat sarcoma-phosphatidylinositol 3-kinase -Protein Kinase B (Ras-PI3K-Akt) and the phospholipase C-gamma 1 pathways 9,10 . In mesencephalic primary cultures, NT-4/5 has been shown to enhance the morphological complexity and to stimulate the survival of DAergic neurons 11,12 . In line with these reports, we have shown that GDNF and NT-4/5 administration resulted in a significantly higher survival rate of rat ventral mesencephalic (VM) tyrosine hydroxylase immunoreactive (TH-ir) neurons grown for 1 week as free-floating roller-tube (FFRT) cultures and also increased their 3,4-dihydroxyphenethylamine (dopamine; DA) release 6 .
Transient storage of donor tissue in FFRT culture offers the possibility of exposing cells with growth factors prior to grafting. We, and others, have previously shown that pretreated donor tissue resulted in a better outcome 13 . Support for growth factor pretreatment prior to transplantation is furthermore given by a recent study which demonstrated that fibroblast growth factor 2–mediated pregrafting expansion of primary VM precursor cells considerably improved DAergic cell survival and functional restoration in a rat model of PD 14 .
To the best of our knowledge, so far no studies have been performed analyzing the effects of a combined GDNF and NT-4/5 administration on VM tissue of human origin. Hence, in the present study, we aimed at investigating whether treatment with combined exposure to GDNF and NT-4/5 improves DAergic neuron survival and augments dopamine release as compared to a single-factor treatment.
Materials and Methods
Human Tissue
All experiments were carried out in accordance with the Ethics Committee of the Medical Faculty of the University of Bern and the Ethics Committee of the State Bern, Switzerland (study no. 181/07). Written consent was given by the women seeking abortion.
Tissue culture
FFRT cultures were prepared from VM derived from aborted human fetuses as described in detail by Spenger and colleagues 15,16 . The embryonic age of the specimen was 8- to 9-wks postconception for analysis of cell numbers and dopamine content and 7- to 10-wks postconception for Western blot analysis and lactate dehydrogenase (LDH) measurements as determined by the method of Evtouchenko and colleagues 17 . In brief, the VM tissue was dissected out and divided into two equally sized portions corresponding to the left and right side and then further divided into equally sized small pieces. Each piece was transferred into labeled conical plastic tubes (BD Falcon, Allschwil, Switzerland) containing 1 mL of culture medium and placed in a roller drum in an incubator for 1 wk 16 . The cultures were grown in medium containing 55% Dulbecco’s modified Eagle medium (DMEM) (Gibco, Reinach, Switzerland), 32.5% Hank’s balanced salt solution (HBSS, Gibco), 1.5% glucose, 10% fetal calf serum (Gibco), and 1% 0.01 M Hepes (Merck KGaA, Darmstadt, Germany). During the entire culture period, antibiotics (Gibco) were present. The culture tubes were randomly assigned to the GDNF (10 ng/mL; Promega AG, Dübendorf, Switzerland), NT-4/5 (10 ng/mL; Promega), the GDNF plus NT-4/5 group, and to the control group (n = 4–12 cultures per group for each of the 3 independent experiments). The medium was changed every other day. Neurotrophic factors were added at day 0 and then at each medium change. Control cultures were grown in medium with no trophic factors added.
Analysis of dopamine content in the culture medium by high-performance liquid chromatography (HPLC)
At the end of the culture period, the medium was collected and a dopamine-stabilizing solution (0.22 mg metabisulfite per 50 μl in 7.5% orthophosphoric acid) was added. Dopamine was determined using reversed phase HPLC with electrochemical detection as described in detail elsewhere 6 . Mean dopamine content in control cultures was 24.1 ± 4.4 pg/mL (mean ± standard error of the mean [SEM]).
Immunohistochemistry and morphometric analyses
Cultures were washed 2 times with ice-cold HBSS, pooled, and fixed in 4% paraformaldehyde in 0.1 M phosphate-buffered saline (PBS; pH 7.4) for 45 min at room temperature followed by cryoprotection in 20% sucrose in 0.1 M PBS containing 0.01% NaN3 for 24 hours. Cultures were sectioned at 20 µm on a freezing microtome (Frigocut 2800N, Reichert-Jung) and mounted on gelatin chrome alum–precoated glass slides (Sigma-Aldrich Chemie GmbH, Buchs, Switzerland). Sections were washed in PBS preincubated for 60 min in PBS containing 0.3% Triton X-100 and 10% horse serum and incubated for 48 h at 4 °C with primary antibodies against rabbit anti-TH (1:1,000, Pel-Freez Biologicals, Rogers, AR, USA) diluted in PBS containing 0.1% Triton X-100 and 2.5% horse serum. After washing, sections were incubated with biotinylated anti-rabbit antibodies (1:200; Vector, Burlingame, CA, USA). Bound antibodies were visualized with the avidin–peroxidase complex method and the metal enhanced 3,3′-diaminobenzidine-based kit (Pierce, Rockford, IL, USA). Slides were then washed, dehydrated, and covered with Eukitt (O. Kindler GmbH, Freiburg, Germany). Quantification of TH-ir neurons was performed by bright-field microscopy by means of a neuron tracing system using a 20× objective 6 . Cell numbers were corrected for double counting according to Abercrombie’s formula 18 and were normalized as TH-ir cell densities (number of TH-ir cells/mm2) and shown as percentage of controls. Mean TH-ir cell densities in control cultures were 71.3 ± 8.3 cells/mm2 (mean ± SEM). Culture volumes were assessed by computer-assisted image analysis as described elsewhere 6 . Mean culture volumes in control cultures were 0.47 ± 0.07 mm3 (mean ± SEM).
Analysis of LDH content in the culture medium
At the end of the culture period, the medium was collected and used for LDH-level measurements as described in detail earlier 6 . Values are expressed as percentage of controls of 4 independent experiments (n = 17–20 per group).
Western blotting
Proteins were analyzed by SDS-PAGE minigel electrophoresis as described previously 6 . Briefly, cultures were washed 2 times with ice-cold HBSS and then pooled for each group. The prepared lysates were subjected to a 12% SDS-PAGE separating gel, and the proteins blotted to a polyvinylidene difluoride membrane (Bio-Rad, Basel, Switzerland). After electrophoretic transfer, the blots were decorated with mouse anti-TH (1:500; Millipore, Billerica, MA, USA) or mouse anti-GFAP (1:500; Millipore) antibodies. Visualization of bound antibody was performed using anti-mouse antibody–peroxidase conjugates (1:10,000; Nordic Immunological Laboratories, Tilburg, The Netherlands) and the enhanced chemiluminescence method (Amersham, Glattbrugg, Switzerland). For loading control and densitometric analyses, membranes were reprobed with mouse monoclonal α–tubulin (1:20,000; Sigma-Aldrich Chemie GmbH, Buchs, Switzerland). Experiments were done in triplicate.
Statistics
For the statistical analysis, a commercially available software package was used (GraphPad Prism 6, La Jolla, CA, USA). The experimental groups were compared using the Kruskal–Wallis nonparametric analysis of variance test in combination with the Dunn’s multiple comparison post hoc analysis. Differences were considered statistically significant at P < 0.05. Values are presented as mean ± SEM.
Results
Immunohistochemical analysis of the cultures disclosed higher TH-ir neuron cell densities in the GDNF, NT-4/5, and GDNF plus NT-4/5 treated groups (3-fold, 5-fold, and 9-fold after GDNF, NT-4/5, and the combination treatment, respectively) as compared to untreated controls (Fig. 1A). Moreover, the combination treatment resulted in increased surviving TH-ir cell numbers as compared to all other groups (Fig. 1A). In contrast, culture volume did not differ significantly between control and experimental groups with slightly increased volumes found for the GDNF plus NT-4/5 group (by 22%) as compared to control groups (Fig. 1B). Similar to the outcome seen on cell densities of TH-ir neurons, significantly augmented TH protein levels in tissue lysate were observed in the growth factor treated groups as compared to control groups (Fig. 2A and B). We next reasoned whether the release of dopamine might have paralleled these effects on the TH-ir neurons. Indeed, the levels of dopamine in the culture medium were increased in all neurotrophic factor–treated groups (4.5-fold, 5.5-fold, and 6.5-fold after GDNF, NT-4/5, and the combination treatment, respectively) as compared to control groups. Accordingly, the treatment with NT-4/5 and GDNF + NT-4/5 exerted the same increase in dopamine release in the culture medium (Fig. 1C).
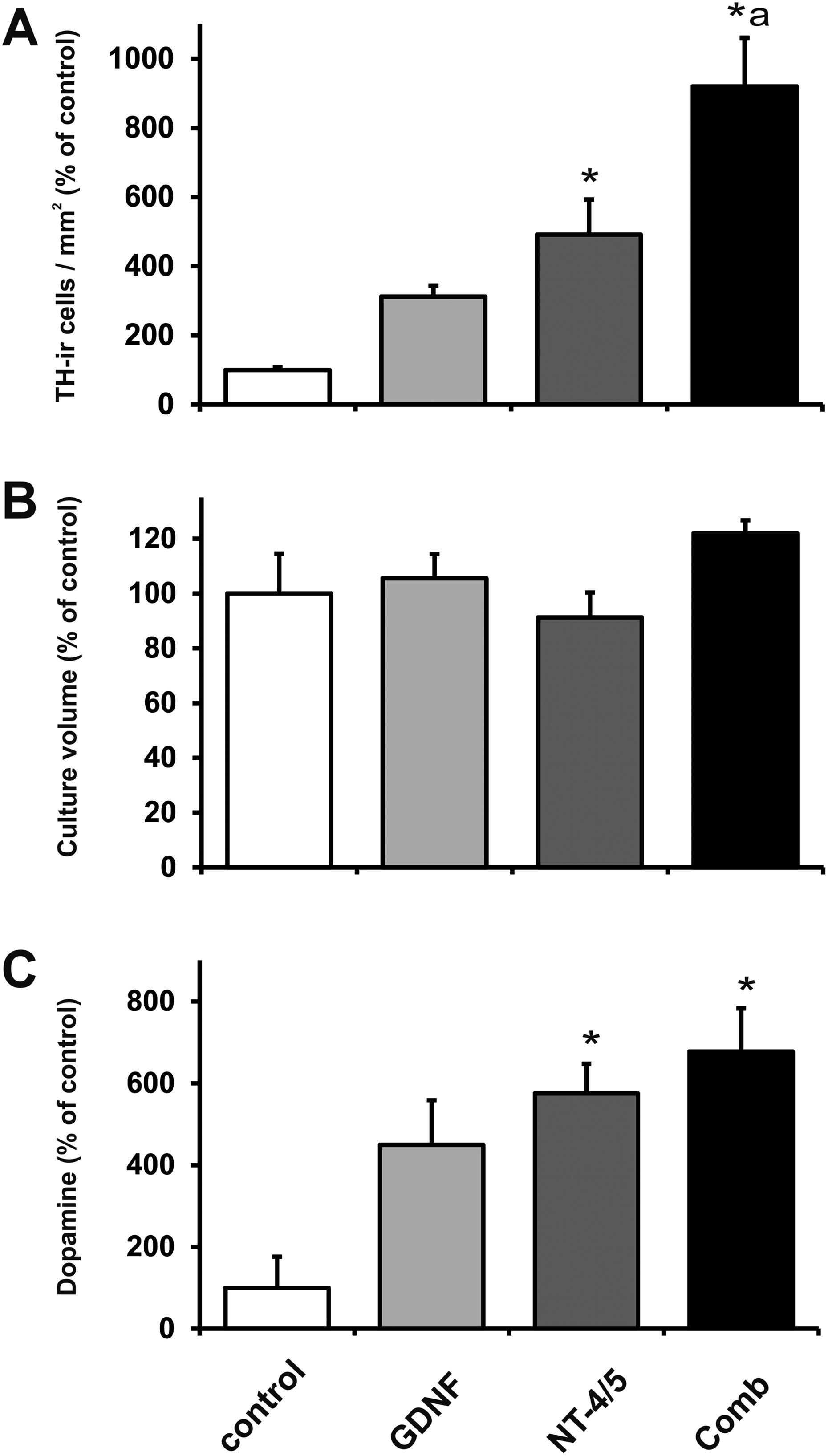
Effects of glial cell line–derived neurotrophic factor (GDNF; gray bars), neurotrophin-4/5 (NT-4/5; dark gray bars), and combined neurotrophic factor treatment (black bars) on tyrosine hydroxylase (TH) positive cell densities (A), culture volume (B), and dopamine content in the culture medium (C). Ventral mesencephalic (VM) free-floating roller-tube (FFRT) cultures were grown for 1 wk in absence (control, open bars) or presence of neurotrophic factors. Data in the bar graphs are given as a percentage of control values and are expressed as mean + standard error of the mean. *P < 0.05 versus corresponding control. a P < 0.05 versus all groups.

Representative microphotographs tyrosine hydroxylase (TH)-immunoreactive cells from human ventral mesencephalic (VM) free-floating roller-tube (FFRT) cultures grown for 7 d without (control; A) or with addition of neurotrophin (NT)-4/5 and glial cell line–derived neurotrophic factor (GDNF; comb; B). Scale bars: 100 µm and 50 µm (inserts). Representative Western blot analyses for TH (C) and glial fibrillary acidic protein (D) protein levels from human VM free-floating roller-tube FFRT cultures grown for 7 d without (control) or with addition of NT-4/5 and GDNF (comb). Note that the combined treatment resulted in a significantly increased TH signal intensity as compared to the control group (C), whereas no difference in GFAP signal intensities between groups was observed (D). Membranes were reprobed with α-tubulin as a loading control. *P < 0.05 versus corresponding control.
Western blot analysis for GFAP did not show increased levels in the neurotrophic factor–treated cultures as compared to controls (Fig. 2C and D). The level of LDH, released into the culture medium from dead or degenerating cells, was lower after growth factor treatment compared to untreated control cultures (100.0% ± 5.2%, 90.5% ± 1.3%, 91.4% ± 0.9%, and 90.2% ± 1.6% for control, GDNF, NT-4/5, and the combination treatment, respectively).
Discussion
In the present work, we demonstrate that the combination of NT-4/5 and GDNF has an additive effect on TH-ir cell numbers in organotypic explant cultures of fetal human ventral mesencephalon. Moreover, we have found that the combined treatment is superior to each of the single factors in improving the survival and the expression of structural and biochemical functional markers of cultured fetal DAergic neurons. Overall, these observations substantiate and extend our earlier report on VM-FFRT cultures of rat origin 6 .
Optimal growth, differentiation, and survival of DAergic neurons during development require a concerted action of different neurotrophic factors. In translational settings, several studies have addressed whether the neuronal supporting actions of different neurotrophic factors including GDNF and neurturin can be exploited to improve the efficacy of cell transplantation approaches for PD. Indeed, GDNF has already been reported to reduce apoptosis in DAergic neurons 19 and to promote DAergic fiber growth in fetal nigral grafts 20 . Application of NT-4/5 and GDNF furthermore protected DAergic neurons from toxin-induced cell death 21 , and GDNF was found to exert antiapoptotic effects on the DAergic neurons 19,22 . These observations suggest that the reduction in apoptosis events might be one of the mechanisms involved in the higher number of TH-ir neurons in the NT-4/5- and GDNF-treated samples. Indeed, this notion seems to be supported by the lower LDH levels measured after growth factor treatment. Notably and conversely to the additive actions observed on TH-ir cell densities, however, levels of released LDH did not differ among the growth factor-treated groups. Thus, this outcome does not support the idea that the higher TH-ir cell densities are the result of a substantial reduction in cell death; rather, these findings might be better explained by an enhanced specific maturation and/or differentiation of the TH-ir cells subpopulation. In agreement with this hypothesis, we have observed no significant differences in culture volume between groups. Moreover, in agreement with our previous study using cultures of rat origin 6 , GFAP protein levels were not affected by the treatments. Nevertheless, the mechanisms underlying the increased TH-ir cell densities were not investigated in the present study, and we cannot exclude that other neuronal phenotypes were also promoted by the treatments. In fact, it has been reported that GDNF and NTs stimulate the survival and growth of a wider range of neurons including striatal and cortical GABAergic neurons 9,23 –25 .
From a functional standpoint in the cell transplantation paradigm for PD, the capacity to release substantial amounts of dopamine is a crucial disease-changing pattern. We previously demonstrated that BDNF treatment increased the number of cultured DAergic neurons and elevated dopamine levels in the culture medium 26 . Moreover, we reported a moderate but beneficial survival-promoting effect of combined GDNF and BDNF treatment on rat fetal nigral tissue 20 . In the present study, HPLC measurements of dopamine in the culture medium disclosed pronounced effects after single factor treatment as compared to controls. Interestingly, only a tendency for additivity was detected. Although this outcome appears in contrast to the findings of Studer and colleagues, that is, that TH-ir cell numbers correlate with dopamine content in the culture medium 27 , it should be noted that in the mentioned study only cultures not exposed to growth factors were investigated. The mechanisms accounting for our observation need to be further explored and might involve differences in the degree of DAergic neuron maturation induced by NT-4/5 and GDNF possibly due to differences in the expression of the respective receptors. Importantly to note, even though Mendez et al. reported a substantial increased fluorodopa uptake in 2 parkinsonian patients receiving grafts exposed to GDNF, 28 it is clearly necessary to verify the effectiveness of the combined NT-4/5 and GDNF pretreatment regimen of human mesencephalic tissue in animal models of PD.
Conclusions
Recent evidence supports the view that the intrastriatal delivery of DAergic neurons might develop into a safe and effective therapeutic option for PD 29 . Although the human iPSC-derived DAergic cells hold a considerable potential due to the unrestricted availability, high purity, and minor ethical concerns, they appear to be less effective compared to the fetal-derived neurons. On the other hand, the limited availability of fetal donor tissue and the generally poor survival of the grafted DAergic neurons are still major issues to be solved for successful cell replacement interventions 29 . The present study demonstrates that exposure of human DAergic nigral neurons grown as FFRT cultures to a combination of GDNF and NT-4/5 significantly improved quality of donor tissue and that this combination is overall superior compared to the single-factor treatments. Further studies in PD will be needed to verify the significance of the preengraftment treatment described here. Nevertheless, it can be assumed that our findings likely also have impact for cell replacement approaches using iPSC- and/or ESC-derived donor cells.
Footnotes
Acknowledgments
We are grateful to Dr. Matthias Bischof, Tanja Bosnjak, Sandra Krebs, Inge Nielsen, and Dr. Alberto Perez-Bouza, Dr. Jürgen Sautter, and Dr. Christian Spenger for excellent technical assistance and their interest in the work. We are indebted to the Department of Gynecology, University of Bern, Switzerland, for their contribution.
Ethical Approval
This study was approved by the Ethics Committee of the Medical Faculty of the University of Bern and the Ethics Committee of the State Bern, Switzerland (study no. 181/07).
Statement of Human and Animal Rights
This study was conducted on deceased human VM tissue, therefore no statement of human and animal rights is applicable. VM tissue was obtained from aborted fetuses.
Statement of Informed Consent
Written consent was given by the women seeking abortion.
Declaration of Conflicting Interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was supported by the HANELA Foundation and the Swiss National Science Foundation (No. 3100A0-112529). MM was supported by the Danish Parkinson Association, IMK Almene Fond, Kirsten and Freddy Johansens Fond, and Hørslev Fonden.