
Abstract
Background
Lung cancer is the most common malignancy, and its mortality ranks first among malignancies. Non-small cell lung carcinoma (NSCLC) is the most common pathological subtype of lung cancer. It is reported that circular RNAs (circRNAs) feature prominently in the occurrence and metastasis of NSCLC.
Purpose
This study aims to decipher the biological functions of circ_0006220 in NSCLC and the underlying mechanism.
Methods
The microarray data (GSE101586) were downloaded from the Gene Expression Omnibus database, and differentially expressed circRNAs in NSCLC tissues were screened using the GEO2R tool. Quantitative real-time polymerase chain reaction was used for detecting the expression of circ_0006220, miR-203-3p, and regulator of G-protein signaling 17 (RGS17) mRNA in NSCLC tissues and cells. The connection between circ_0006220 expression and clinicopathological indicators was analyzed through the chi-square test. EdU and cell counting kit-8 assays were carried out to detect cell growth. Cell migration and invasion were detected by transwell assays. Bioinformatics was used to predict, and RNA immunoprecipitation assay and dual-luciferase reporter gene assay were conducted for verifying, the targeted relationship among circ_0006220, miR-203-3p, and RGS17.
Results
The expression of circ_0006220 was elevated in NSCLC cells and tissues, and high circ_0006220 expression was significantly associated with unfavorable clinicopathological indicators. In addition, it was revealed that circ_0006220 overexpression facilitated NSCLC cell growth, migration, and invasion, whereas knocking down circ_0006220 had contrary effects. Furthermore, miR-203-3p was identified as a downstream target of circ_0006220, and circ_0006220 could sponge miR-203-3p; RGS17 was identified as a downstream target of miR-203-3p and was positively modulated by circ_0006220.
Conclusions
Circ_0006220 up-regulates RGS17 expression by adsorbing miR-203-3p to promote NSCLC development.
Introduction
At present, known as the malignancy with the highest morbidity and mortality globally, lung cancer has become one of the leading causes of death; non-small cell lung carcinoma (NSCLC) patients make up over 85% of all lung cancer cases. 1 Although the treatment strategies for NSCLC have developed in recent years, the 5-year survival rate of NSCLC patients is still quite low. 2 In this context, further study on the mechanism of NSCLC progression is urgently needed.
Circular RNAs (circRNAs) are considered to be non-coding RNAs naturally produced in the eukaryotic transcriptome, with stable structure and high tissue specificity; circRNAs have been proven to have various biological regulatory functions, for instance, acting as miRNA sponges, forming RNA-protein complex, and regulating the transcription and alternative splicing of target genes.3,4 For example, circ_ARHGAP10 promotes GLUT1 expression and inhibits NSCLC development via acting as the competing endogenous RNA (ceRNA) of miR-150-5p. 5 Another study reports that circ_FGFR3 promotes the expression of galectin-1 (Gal-1), p-ERK1/2 and p-AKT via competitively binding to miR-22-3p, and facilitates NSCLC progression. 6 Additionally, circRNA_101237 is highly expressed in NSCLC cells and tissues, and it promotes NSCLC development by regulating the miR-490-3p/MAPK1 axis. 7 In this study, our bioinformatics analysis indicates that circ_0006220 is highly expressed in NSCLC. Nonetheless, the roles and regulatory mechanism of circ_0006220 in NSCLC warranted further investigation.
MicroRNAs (miRNAs, miRs) are involved in regulating almost all aspects of cellular processes; miRNAs provide a new insight into understanding tumorigenesis and cancer progression.8,9 For example, miR-621 targets SIX4 to inhibit NSCLC progression. 10 It has been previously reported that miR-203 expression is reduced in NSCLC, and miR-203 targets and regulates RGS17 expression to play a tumor-suppressing role in NSCLC. 11 Interestingly, our bioinformatics analysis suggests that miR-203-3p is a potential downstream target of circ_0006220. We supposed that circ_0006220 could probably exert oncogenic functions via regulating miR-203-3p. This study was performed to validate this hypothesis. We report that circ_0006220 is highly expressed in NSCLC tissues and cell lines, and circ_0006220 plays a cancer-promoting role in NSCLC development via modulating the miR-203-3p/RGS17 axis.
Materials and methods
Ethics statement and tissue samples
For this study, the para-cancerous and cancerous tissues of 45 NSCLC patients diagnosed in Shaanxi Provincial People’s Hospital were collected from February 2018 to March 2020. These samples were obtained during surgery and quickly frozen in liquid nitrogen. All patients were diagnosed with NSCLC through clinical, imaging, and histological examination, and all patients did not suffer from other malignancies. This study was endorsed by the Ethics Committee of Shaanxi Provincial People’s Hospital. The subjects all signed an informed consent. None of the subjects received chemotherapy, radiotherapy, or targeted therapy before the surgery.
Cell culture and transfection
Normal human bronchial epithelial cells (HBE cell line), human embryonic kidney cells (HEK293T cell line), and NSCLC cell lines (H1299, H1975, A549, and H2228) were purchased from ATCC (Rockville, MD, USA). These cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Thermo Fisher Scientific, Waltham, MA, USA) with 100 U/mL penicillin, and 0.1 mg/mL streptomycin (Biyuntian Biotechnology Co., Ltd, Shanghai, China), and 10% fetal bovine serum (FBS; Biyuntian Biotechnology Co., Ltd, Shanghai, China) at 37°C in 5% CO2. The medium was replaced every 2 or 3 days. When cell confluence reached 70–80%, 0.25% trypsin (Roche, Basel, Switzerland) was used for passage. Circ_0006220 overexpression plasmid (pcDNA-circ_0006220), empty vector cDNA (pcDNA-NC), small interfering RNA (siRNA) oligonucleotides targeting human circ_0006220 (si-circ_0006220-1, si-circ_0006220-2), siRNA negative control (si-NC), miR-203-3p inhibitor and its control (Inh NC), and miR-203-3p mimic and its control (miR-NC) were constructed by GenePharma (Shanghai, China). Lipofectamine® 3000 (Invitrogen, Carlsbad, CA, USA) was used for transfecting the above vectors/oligonucleotides into the cells. After 24 h, quantitative real-time polymerase chain reaction (qRT-PCR) was performed for detecting cell transfection efficiency.
qRT-PCR
TRIzol reagent (Invitrogen, Carlsbad, CA, USA) was employed for extracting total RNA from cells and tissues. After detection of the RNA concentration and purity, the reverse transcription of total RNA into cDNA was carried out with a first-strand cDNA synthesis kit (Thermo Fisher Scientific, Waltham, MA, USA). With the cDNA as the template, qRT-PCR was conducted using a SYBR® Premix-Ex-Taq™ kit (Takara, Tokyo, Japan) in an ABI7300 system (Thermo Fisher Scientific, Waltham, MA, USA). U6 and GAPDH were utilized as the endogenous controls. To determine the subcellular localization of circ_0006220 in NSCLC cells, a PARIS™ kit (Thermo Fisher Scientific, Waltham, MA, USA) was adopted for performing subcellular fractionation. Then, circ_0006220 expression in cytoplasm and nucleus was detected by qRT-PCR. U6 and GAPDH functioned as control for nucleus and cytoplasm, respectively. Below are the specific primer sequences:
Circ_0006220 Forward: 5′-TGTTTGCATCTACCCTGCTG-3′
Circ_0006220 Reverse: 5′-ACACTCCTCCTTGGTCTTGG-3′
miR-203-3p Forward: 5′-GCGCGTGAAATGTTTAGGAC-3′
miR-203-3p Reverse: 5′-GTGCAGGGTCCGAGGT-3′
RGS17 Forward: 5′-CTCGAGGTAACAA ACCA ATTC TTACAA CTATATAC-3′
RGS17 Reverse: 5′-GCGGCCGCTGCCACAGTTTTGGCAGTGAAC-3′
U6 Forward: 5′-CTCGCTTCGGCAGCACA-3′, U6 Reverse: 5′-AACGCTTCACGAATTTGCGT-3′; GAPDH Forward: 5′-GCACCGTCAAGGCTGAGAAC-3′, GAPDH Reverse: 5′-TGGTGAAGACGCCAGTGGA-3′.
Cell counting kit-8 (CCK-8) assay
H1299 or A549 cells during logarithmic growth were harvested, and the cell density was adjusted to 2 × 104 cells/ml after trypsinization. Afterward, the cells were transferred to 96-well plates with three replicate wells in each group and 100 μL of cell suspension per well. Subsequently, the 96-well plates were put in the incubator to continue the culture. After 24, 48, 72, or 96 h, each well was added with CCK-8 solution (10 μL per well; MedChemExpress, Monmouth Junction, NJ, USA), and the cells were incubated in the incubator for 1 h. After the culture was finished, the 96-well plates were put in a microplate reader for measuring the absorbance at 450 nm of each well.
EdU assay
A EdU kit (Beyotime Biotechnology, Shanghai, China) was utilized to perform the EdU assay following the manufacturer’s instructions. Briefly, when the confluence of H1299 or A549 cells reached about 50%, the medium was replaced with fresh medium containing 50 μmol/L EdU, and the cells were cultured for another 24 h. Next, the medium was discarded, and the cells were fixed for 15 min in 4% polyformaldehyde, followed by incubation for 10 min with 0.2% glycine, and the cells were subsequently washed twice with PBS. The incubator was placed in the dark for 10 min. Then, the cells were incubated with 0.5% Triton X-100 for 10 min, and then stained with Apollo in the dark at room temperature for 30 min. Next, the cells and DAPI staining solution were incubated in the dark for 20 min at room temperature. After PBS washing, the cells were photographed and counted under a fluorescence microscope, and the proportion of EdU-positive cells was calculated.
Transwell assay
The lower compartment of the transwell chamber (Costar, Cambridge, MA, USA) was added with 500 μL of medium, and 5 × 104 transfected cells in serum-free medium were transferred into the upper compartment. The transwell plate was put in the incubator, and 24 h later, a cotton swab was employed for wiping away the non-migrated cells in the upper compartment. Then, crystal violet staining solution was added, and the migrated cells were stained for 15 min at room temperature. After the cells were washed and air-dried, the cells were counted and photographed in five random fields under the inverted microscope. A transwell chamber without Matrigel (1:10; BD Biosciences, Franklin Lakes, NJ, USA) was used for the migration assay, and Matrigel was used for the invasion assay.
RNA immunoprecipitation (RIP) assay
A Magna RIP RNA-binding protein immunoprecipitation kit (Millipore, Billerica, MA, USA) was utilized to perform RIP assay. RIP lysis buffer was employed to lyse H1299 and A549 cells, and whole cell extract (100 μL) was incubated with magnetic beads coupled with human anti-argonaute2 (Ago2) antibody (Millipore, Billerica, MA, USA), and they were incubated at 4°C for 8 h. The negative control was anti-IgG antibody (Millipore, Billerica, MA, USA). Next, washing buffer was used for washing the immunoprecipitate, which was then incubated with proteinase K for 30 min at 55°C to separate the RNA-protein complex from the magnetic beads and remove the protein. Ultimately, qRT-PCR was performed to analyze the enrichment of circ_0006220 and miR-203-3p in the immunoprecipitated RNA.
Dual-luciferase reporter assay
Bioinformatics analysis was utilized for predicting the binding sequences between circ_0006220 and miR-203-3p or between miR-203-3p and RGS17 mRNA 3′UTR. The circ_0006220 or RGS17 3′UTR fragments with the predicted binding sequence or mutated sequence was amplified and cloned into pmirGLO Dual-Luciferase miRNA Target In Expression Vector (Promega, Madison, WI, USA) to construct wild-type (circ_0006220 WT, RGS17 WT) or mutant (circ_0006220 MUT, RGS17 MUT) luciferase reporter gene vectors. The above-mentioned reporter vectors and miR-203-3p mimics or its negative control were co-transfected into 293T cells, respectively. The dual-luciferase assay system (Promega, Madison, WI, USA) was adopted to measure the luciferase activity after 48 h of transfection.
Western blot assay
NSCLC cells were lysed with RIPA lysis buffer (Beyotime Biotechnology, Shanghai, China), and the supernatants of the lysates were collected after centrifugation to extract total cellular protein. The total protein concentration was determined by the BCA method. The supernatant was heated for 10 min in boiling water to denature the proteins. Next, the proteins were separated via SDS-PAGE and then transferred onto the polyvinylidene fluoride (PVDF) membrane (Millipore, Bedford, MA, USA). The PVDF membranes were blocked with 5% skim milk, and then they were incubated overnight with primary antibodies: anti-RGS17 antibody (ab150814, 1:500, Abcam, Cambridge, UK) and anti-GAPDH (ab9485, 1: 1000, Abcam, Cambridge, UK) at 4°C. After rinsing the membranes with TBST solution, the membranes were incubated with goat anti-rabbit IgG H&L (ab205718, 1:2000, Santa Cruz Biotechnology, Santa Cruz, CA, USA) at room temperature for 1 h. Eventually, the membranes were rinsed 4 times with TBST. An ECL chemiluminescence kit (Amersham Pharmacia Biotech, Little Chalfont, UK) was used for protein band developing. GAPDH acted as the internal reference.
Statistical analysis techniques
All of the experiments were performed at least in triplicate. SPSS 24.0 (SPSS Inc., Chicago, IL, USA) was used as the statistical analysis tool. Measurement data were expressed as “mean ± standard deviation.” Student’s t-test was conducted to perform the comparison between two groups. Comparison among multiple groups was performed with one-way analysis of variance. Enumeration data were presented in a contingency table. The differences between two groups were analyzed by the χ2 test. A difference was statistically significant when P < 0.05.
Results
Circ_0006220 was highly expressed in NSCLC cells and tissues
The GSE dataset GSE101586 was downloaded from GEO database, and the differentially expressed circRNAs in NSCLC tissues compared with adjacent tissues were analyzed. Following the criteria (log2 |fold change| > 1, P < 0.05), it was found that circ_0006220 expression was markedly elevated in NSCLC tissues (Figure 1(a) and (b)). Consistently, qRT-PCR showed that circ_0006220 was significantly highly expressed in NSCLC tissues (Figure 1(c)). In contrast to HBE cells, circ_0006220 expression was remarkably up-regulated in NSCLC cell lines (H1299, H1975, A549, and H2228 cells) (Figure 1(d)). Furthermore, it was revealed that high circ_0006220 expression was associated with increased tumor size and the advanced TNM stage (Table 1). These data suggested that circ_0006220 could probably participate in regulating NSCLC progression. Moreover, it was revealed that circ_0006220 was distributed mainly in the cytoplasm of NSCLC cells, which implied that it might exert its biological function as a molecular sponge (Figure 1(e)). Circ_0006220 expression was significantly up-regulated in NSCLC tissues and cells. (A–B) Differentially expressed circRNAs in NSCLC tissues compared with normal tissues were analyzed with dataset GSE101586. (C) qRT-PCR was performed to detect circ_0006220 expression in NSCLC tissues and para-cancerous tissues. (D) qRT-PCR was used to detect circ_0006220 expression in NSCLC cell lines. (E) The nucleocytoplasmic separation experiment was used to detect the subcellular location of circ_0006220. ** P < 0.01 and *** P < 0.001. Correlation between clinicopathological features and circ_0006220 expression in NSCLC. P < 0.05.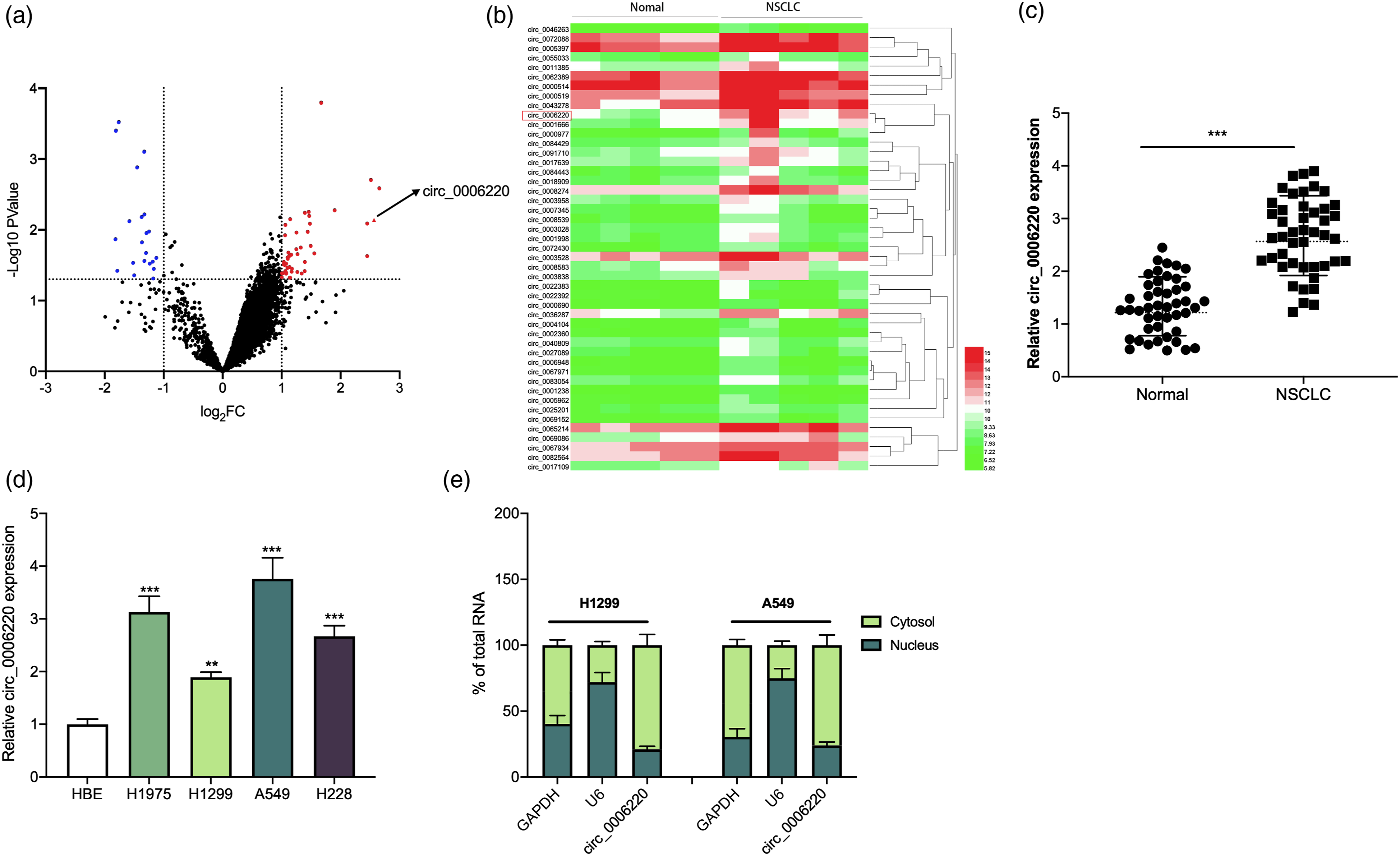

Circ_0006220 facilitated NSCLC cell growth, migration, and invasion
In NSCLC cell lines, circ_0006220 expression was the lowest in H1299 cells, and its expression was the highest in A549 cells (Figure 1(d)). Therefore, H1299 cells were transfected with circ_0006220 overexpression plasmids and A549 cells were transfected with si-circ_0006220-1 and si-circ_0006220-2 (Figure 2(a) and (b)). Since the knockdown effect of si-circ_0006220-1 was more significant, si-circ_0006220-1 was used for subsequent experiments. Results of CCK-8, EdU, and transwell assays suggested that circ_0006220 overexpression in H1299 cells significantly promoted cell growth, migration, and invasion, while circ_0006220 knockdown in A549 cells inhibited cell growth, migration, and invasion (Figure 2(c) to (e)). Circ_0006220 promoted NSCLC cell proliferation, migration, and invasion. (A–B) The transfection efficiency of pcDNA-circ_0006220, si-circ_0006220-1, and circ_0006220-2 was detected by qRT-PCR. (C–D) CCK-8 and EdU assays were used to detect the effects of circ_0006220 overexpression or knockdown on H1299 and A549 cell proliferation. Scale bars, 150 μm. (E) Transwell assay was used to determine the effects of circ_0006220 overexpression or knockdown on H1299 and A549 cell migration and invasion. Scale bars, 50 μm. * P < 0.05, ** P < 0.01, and *** P < 0.001.
Circ_0006220 directly targeted miR-203-3p
Next, the Circinteractome database (https://circinteractome.nia.nih.gov) was searched, and it predicted that there were three potential target miRNAs of circ_0006220: miR-203-3p, miR-1299, and miR-520f. Among them, miR-203-3p showed the highest Context + score percentile (Figure 3(a)). Furthermore, the targeted relationship between miR-203-3p and circ_0006220 was verified by the luciferase reporter gene assay: miR-203-3p overexpression notably repressed the luciferase activity of circ_0006220 WT reporter; miR-203-3p inhibition markedly promoted the luciferase activity of circ_0006220 WT reporter, whereas neither up-regulation nor down-regulation of miR-203-3p expression had a significant effect on the luciferase activity of circ_0006220 MUT reporter (Figure 3(b)). RIP assay showed that compared with the IgG group, circ_0006220 and miR-203-3p were markedly enriched in the Ago2 group (Figure 3(c)). Next, qRT-PCR revealed that circ_0006220 overexpression in H1299 cells significantly inhibited miR-203-3p expression, while circ_0006220 knockdown in A549 cells induced up-regulation of miR-203-3p expression (Figure 3(d)). Moreover, it was revealed that miR-203-3p expression in NSCLC tissues was under expressed in adjacent lung tissues (Figure 3(e)); in comparison to HBE cells, miR-203-3p expression in NSCLC cell lines was also markedly reduced (Figure 3(f)). In addition, the results of Pearson correlation analysis suggested that in NSCLC tissues, circ_0006220 expression and miR-203-3p expression were negatively correlated (Figure 3(g)). The above-mentioned findings indicated that miR-203-3p was the downstream target of circ_0006220. Circ_0006220 adsorbed miR-203-3p. (A) The potential binding site between circ_0006220 and miR-203-3p was predicted by the Circinteractome database. (B) The relative luciferase activity of the cells in each group was detected by dual-luciferase gene reporter assay. (C) RIP assay was used for detecting the relative enrichment of circ_0006220 and miR-203-3p. (D) After overexpression or knockdown of circ_0006220 in NSCLC cells, miR-203-3p expression was detected by qRT-PCR. (E) qRT-PCR was performed to detect miR-203-3p expression in NSCLC tissues. (F) qRT-PCR was performed to detect miR-203-3p expression in NSCLC cell lines. (G) Pearson correlation analysis of the correlation between circ_0006220 and miR-203-3p expressions. ** P < 0.01 and *** P < 0.001.
Circ_0006220 played a cancer-promoting role in NSCLC by targeting miR-203-3p
To understand whether circ_0006220 participated in regulating NSCLC progression by sponging miR-203-3p, pcDNA-NC, pcDNA-circ_0006220, and pcDNA-circ_0006220+miR-203-3p mimics were transfected into H1299 cells, respectively; si-NC, si-circ_0006220, and si-circ_0006220 + miR-203-3p inhibitors were transfected into A549 cells, respectively; and qRT-PCR suggested that the transfection was successful (Figure 4(a)). It was revealed that up-regulating circ_0006220 expression promoted H1299 cell growth, migration, and invasion, yet transfection of miR-203-3p mimics attenuated this effect (Figure 4(b) to (d)); knocking down circ_0006220 inhibited A549 cell proliferation, migration, and invasion, yet this inhibitory effect was reversed by transfection of miR-203-3p inhibitors (Figure 4(b) to (d)). Circ_0006220 played a cancer-promoting role in NSCLC by targeting miR-203-3p. (A) H1299 cells were transfected with pcDNA-NC, pcDNA-circ_0006220, or pcDNA-circ_0006220 + miR-203-3p mimics, respectively, and A549 cells were transfected with si-NC, si-circ_0006220 + miR-203-3p inhibitors, respectively, and qRT-PCR was used to detect miR-203-3p expression. (B–C) CCK-8 and EdU assays were performed to detect the proliferation of H1299 or A549 cells. (D) Transwell assays were conducted to detect the migration and invasion of H1299 or A549 cells. * P < 0.05, ** P < 0.01, and *** P < 0.001.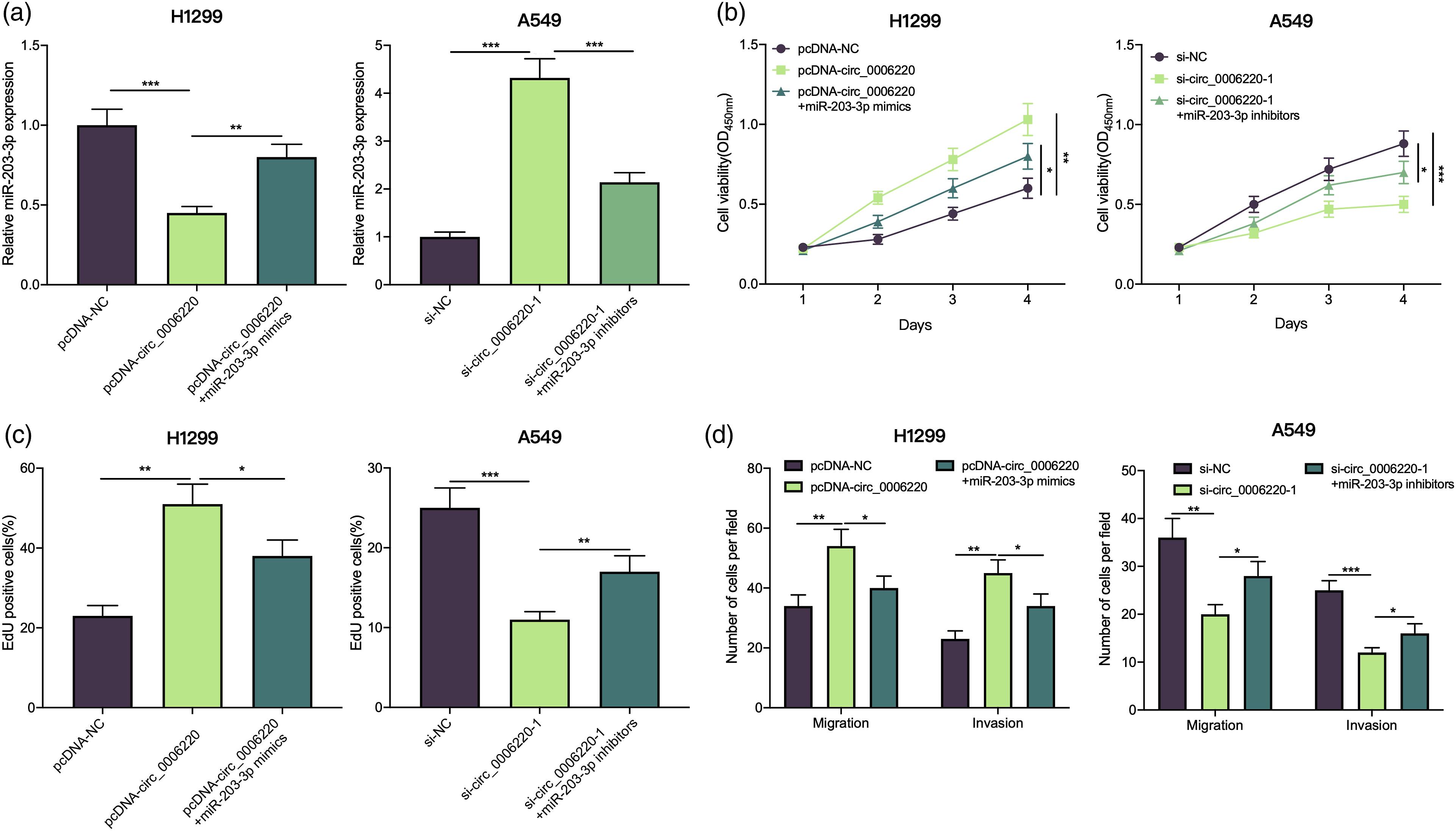
Circ_0006220 participated in NSCLC development via modulating the miR-203-3p/RGS17 axis
To further clarify the underlying mechanism of the circ_0006220/miR-203-3p axis functioning in NSCLC, the TargetScan database (http://www.targetscan.org/) was used for predicting the downstream genes of miR-203-3p, and it was revealed that there existed a binding sequence between miR-203-3p and RGS17 mRNA 3′UTR (Figure 5(a)). Next, dual-luciferase reporter gene assay confirmed that up-regulating miR-203-3p expression remarkably inhibited the luciferase activity of RGS17 WT reporter; down-regulating miR-203-3p expression promoted the luciferase activity of RGS17 WT reporter, yet up-regulation or down-regulation of miR-203-3p expression did not significantly change the luciferase activity of RGS17 MUT reporter (Figure 5(b)). Then, we studied the effect of the circ_0006220/miR-203-3p axis on RGS17 expression, and it was found that up-regulating circ_0006220 increased RGS17 mRNA and protein expression in H1299 cells, but this effect would be weakened by up-regulating miR-203-3p; knocking down circ_0006220 inhibited RGS17 mRNA and protein expression in A549 cells, yet the inhibitory effect was partially counteracted by transfection with miR-203-3p inhibitors (Figure 5(c) and (d)). In addition, compared with adjacent tissues, RGS17 mRNA expression in NSCLC tissues was markedly up-regulated (Figure 5(e)). Correlation analysis revealed that in NSCLC tissues, RGS17 mRNA and miR-203-3p expressions were negatively correlated, and RGS17 mRNA and circ_0006220 expressions were positively correlated (Figure 5(f) and (g)). Overall, these results indicated that RGS17 was the downstream target of miR-203-3p in NSCLC cells, and its expression was negatively regulated by miR-203-3p and positively regulated by circ_00062220. Circ_0006220 played a cancer-promoting role by regulating the miR-203-3p/RGS17 axis. (A) The potential binding sites between RGS17 mRNA 3′UTR and miR-203-3p was predicted by TargetScan database. (B) The relative luciferase activity was detected by dual-luciferase reporter gene assay. (C–D) Western blotting and qRT-PCR were used to measure the expression level of RGS17 expression in protein and mRNA level. (E) qRT-PCR was performed to detect RGS17 mRNA expression in NSCLC tissues and adjacent tissues. (F–G) Pearson correlation analysis of the correlation between RGS17 mRNA and miR-203-3p expressions, as well as between RGS17 mRNA and circ_0006220 expressions. **P < 0.01 and *** P < 0.001.
Discussion
This study demonstrates that circ_0006220 facilitates NSCLC cell growth, migration, and invasion via modulating the miR-203-3p/RGS17 axis, which provides a novel mechanism to explain NSCLC progression.
Known as a new kind of RNA with a covalent closed-loop structure, circRNAs were first discovered in viruses as early as the 1970s, and they have become a new research hotspot in recent years. 12 CircRNAs have the characteristics of stability and evolutionary conservation. 13 Some circRNAs regulate gene expression at the transcriptional, post-transcriptional, and translational levels, and they take part in regulating multiple pathological processes by sponging miRNAs, modulating alternative splicing, binding with proteins, and even encoding polypeptides. 14 Some circRNAs are reported to be associated with NSCLC turmorigenesis and progression. For example, circ_FGFR3 is over-expressed in NSCLC, and it promotes the expression of galectin-1 (Gal-1), p-ERK1/2, and p-AKT through competitively binding with miR-22-3p, thus facilitating NSCLC cell proliferation and invasion. 15 Another study reports that circ_101237 expression is up-regulated in NSCLC cells and is linked to the poor prognosis of lung cancer patients, and circ_101237 plays a cancer-promoting role by regulating the miRNA-490-3p/MAPK1 axis. 16 Circ_0006220 expression is down-regulated in lung fibroblasts of idiopathic pulmonary fibrosis (IPF), and circ_0006220 inhibits lung fibroblast proliferation by regulating the miR-526b/Cav1 axis, thereby inhibiting the excessive deposition of extracellular matrix. 17 For the first time, this study report that circ_0006220 was remarkably highly expressed in NSCLC, and its high expression implies poor prognosis of the patients; circ_0006220 overexpression promotes NSCLC cell growth, migration, and invasion. The aforementioned findings suggested that circ_0006220 played a cancer-promoting role in NSCLC.
MiRNAs are short single RNA transcripts composed of approximately 21–24 nucleotides, and they can regulate the translation of mRNA via binding with 3′UTR. 18 MiRNAs can serve as important regulators in NSCLC progression. 19 For instance, miR-621 expression is decreased in NSCLC tissues, and is related to the survival time of NSCLC patients; miR-621 inhibits NSCLC progression by targeting SIX4. 20 MiR-203 features prominently in regulating NSCLC progression, and miR-203 increases the sensitivity of NSCLC cells to cisplatin by inhibiting Dickkopf-1 expression. 21 Another study reports that miR-203-3p expression is markedly reduced in NSCLC tissues and cells, and its down-regulation is associated with lymph node metastasis, higher TNM stage, and poor prognosis of the patients, and miR-203-3p exerts a tumor-suppressing effect by inhibiting AVL9 expression. 22 Consistently, in the present study, we demonstrate that miR-203-3p expression is reduced in NSCLC cells and tissues. Additionally, it is revealed that circ_0006220, which is mainly located in the cytoplasm of NSCLC cells, serves as the molecular sponge of miR-203-3p in NSCLC and negatively regulates miR-203-3p expression, which partly explains the mechanism of miR-203-3p dysregulation in NSCLC.
RGS17 belongs to the RZ(A) subfamily of regulators of G-protein signaling (RGS protein).23,24 The main functions of RGS17 are to accelerate the hydrolysis of guanosine triphosphate (GTP) and induce the inactivation of G protein. 25 RGS17 has also been reported to regulate the sustaining proliferation of malignant cells. 26 RGS17 expression is up-regulated in rectal cancer, and RGS17 overexpression notably facilitates colorectal cell growth, migration, and invasion. 27 In addition, some studies report that RGS17 expression is increased in lung cancer and prostate cancer tissues; besides, RGS17 promotes the expression of cyclic adenosine monophosphate (cAMP) and cAMP response-element binding protein (CREB) to promote tumor cell proliferation.28,29 Our study confirms that miR-203-3p targets RGS17 and negatively regulates its expression. In addition, circ_0006220 can positively regulate RGS17 expression via repressing miR-203-3p. These findings indicated that the circ_0006220/miR-203-3p/RGS17 axis could participate in NSCLC progression.
To sum up, circ_0006220 is expected to be new diagnostic biomarker and therapeutic target for NSCLC, and we report that the ceRNA network consisting of circ_0006220, miR-203-3p, and RGS17 is a novel mechanism involved in NSCLC progression. Nevertheless, this work still has some shortcomings. Firstly, the sample number of present study is relatively small, and in the following work, more samples should be collected to evaluate the prognostic value of circ_0006220 for NSCLC. Additionally, in vivo models are needed to further consolidate our conclusion in the present study. Last but not least, whether circ_0006220 regulates the progression of NSCLC via other downstream targets or mechanisms remains to be explored in the following studies.
Footnotes
Authors Contribution
Ruilin Chen designed the methodology, the goal of the study, and the logical flow. Shaochun Wang conducted the experiments and analyzed the data. Shaochun Wang and Chengcheng Zhang wrote the initial draft. Shaochun Wang and Ruilin Chen reviewed and revised the draft. All authors read and approved the final manuscript.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study is funded by The mechanism of OPN/FOXM1 signaling pathway in airway remodeling in chronic obstructive pulmonary disease and its clinical value (2020ZDLSF01-06).
Research ethics
Our study was approved by the Ethics Review Committee of Shaanxi Provincial People’s Hospital.
Data availability
The data used to support the findings of this study are available from the corresponding author upon request.