
Abstract
Bisphenol A (BPA) is used to produce polycarbonate plastic and epoxy resins which are used in many consumer products. Most people encounter BPA in their daily routines. However, it has been heavily reported that BPA has a neurotoxic effect. The present study aimed to investigate the effect of lycopene on cognitive deficits induced by a high dose of BPA focusing on mitogen-activated protein kinase (MAPK)/extracellular signal-regulated kinase (ERK) pathway, oxidative stress, apoptosis, and memory retrieval in adult male rats. Therefore, 72 rats were divided into four groups: control group, BPA group (50 mg/kg body weight (bw)) 3 days a week for 42 days, lycopene group (10 mg/kg bw) daily for 42 days, and lycopene + BPA group. Concurrent treatment of lycopene with BPA improved the learning and cognition memory in Morris water maze and novel object recognition tests along with an increase in acetylcholine esterase activity as well as inhibition of oxidative stress by restoring reduced glutathione and suppressing malondialdehyde hippocampal level to their normal levels. Mechanistically, lycopene upregulated the protein expression of tyrosine receptor kinase B, which resulted in an upsurge in its downstream cascades MAPK/ERK1/2/cAMP response element-binding protein (CREB)/brain-derived neurotrophic factor (BDNF) signaling pathway in the hippocampus of BPA-intoxicated rats. Furthermore, concurrent treatment of lycopene with BPA prevented apoptosis by marked decrease in Bcl-2 associated X protein (Bax) gene expression and caspase 3 activity while restoring B-cell leukemia/lymphoma-2 (Bcl-2) gene expression. In conclusion, the present study provided evidence that lycopene exerted a neuroprotective effect against BPA intoxication in hippocampi of rats via its antioxidant properties, activation of MAPK/ERK pathway, and inhibiting a neuronal apoptosis which reflected on improving the learning and cognition memory.
Introduction
Bisphenol A (BPA) (4,4′-(propane-2,2-diyl) diphenol) is an environmental toxicant that is classified as a synthetic xenoestrogen endocrine disruptor. It is used as an additive in the manufacturing of polycarbonate plastics and epoxy resins. It is also used in the production of domestic containers, water bottles, thermal papers, dental sealants, and prostheses and as a constituent of the inner coatings of beverages and food containers. 1
BPA was detected in human urine samples from populations of different countries. 2 Interestingly, leaching of BPA from polycarbonate plastic bottles into water was observed at a rate of 0.2–0.8 ng/h. However, the elevating water temperature caused a dramatic 55-fold increase in the rate of leaching of BPA from containers. 3
Unfortunately, BPA has shown serious hazardous effects. Previous studies showed that exposure to BPA may induce multiple organs toxicity including cardiotoxicity, hepatotoxicity, renal toxicity, and testicular toxicity. 4 –7 However, BPA-induced neurotoxicity is of special interest since BPA can be absorbed following ingestion and may pass the blood–brain barrier. In addition, BPA may accumulate in the brain due to its lipophilic properties. 8
Neurobehavioral deficits such as anxiety, reduced exploratory activity, and impaired spatial memory were reported in male offspring mice following a perinatal BPA exposure. 9 The reported underlying mechanisms comprised oxidative stress and lipid peroxidative capacity, decreased acetylcholinesterase activity, increased apoptosis, decreased glutamate receptors expression, and suppressed cAMP response element-binding protein (CREB) and brain-derived neurotrophic factor (BDNF) phosphorylation. 10 –13 Experimental studies have shown protective effects of natural antioxidants such as curcumin and lipoic acid against BPA-induced cognitive and other neurobehavioral disturbances. 13,14
Lycopene, on the other hand, is an aliphatic hydrocarbon carotenoid that can be found in different fruits and vegetables, for example, watermelon and tomatoes. Carotenoids are lipid-soluble pigments. Interestingly, lycopene has shown powerful antioxidant activities in comparison to other carotenoids and natural compounds. 15
Recently, lycopene has demonstrated remarkable neuroprotective effects in several models of neuroinflammation, dementia, and cognitive deficits. 16,17 These beneficial effects were attributed to the inhibition of nuclear factor kappa B, the preservation of mitochondrial integrity, and the suppression of neuronal apoptosis. 18,19
However, the possible protective effects of lycopene against BPA-induced neurotoxicity in rats have not been investigated yet. Therefore, the present study was carried out to examine the underlying mechanistic pathway by which lycopene may attenuate BPA-induced memory impairment in rats.
Materials and methods
Drugs and chemicals
Lycopene was obtained from Holland & Barrett (UK). BPA and all other chemicals were obtained from Sigma-Aldrich Chemical Co. (St Louis, Missouri, USA).
Animals’ preparation
Seventy-two adult male albino rats aged 8 weeks (190–210 g) were obtained from the breeding unit of the Egyptian Organization of Biological Products and Vaccines (Helwan, Egypt). The animals were caged (five rats per cage) with free access to water (using glass water bottles) and standard chow diet pellets. They were kept under controlled environmental conditions: temperature (24 ± 2 C), humidity (50–70%), and 12-h light/12-h light/dark cycle during the experimentation period. The animals were kept for adaptation for 1 week prior to the experiment. The present investigation was approved by the Ethics Committee, Faculty of Pharmacy, Helwan University, Cairo, Egypt (Protocol Number: 012A2018), which is in agreement with the Guide for the Care and Use of Laboratory Animals published by the US National Institutes of Health (NIH publication No. 85-23, revised 2011).
Experimental design
Rats were randomly selected and categorized by a technical assistant not involved in the analysis into four groups (n = 18) as follows: Control group (vehicle group): Rats were given orally 10% Tween 80 solution daily by gavage, followed by corn oil 1 h later 3 days a week for 42 days. BPA group: Rats were administered 50 mg/kg body weight (bw) of BPA which was dissolved in corn oil. The administration was done orally by gavage 3 days a week for 42 days.
20
Lycopene group: Rats were administered 10 mg/kg bw of lycopene, suspended in 10% Tween 80 solution. This was done on a daily basis orally by gavage for 42 days.
21
Lycopene + BPA group: Rats received orally by gavage 10 mg/kg bw of lycopene, suspended in 10% Tween 80 solution daily. An hour later, this was followed by 50 mg/kg bw of BPA dissolved in corn oil. The BPA administration was done orally by gavage 3 days a week for 42 days.
The administration volumes of vehicles, BPA, and lycopene were 5 mL/kg. Five days before the termination of the experiment, two behavioral tests were carried out on the rats in a special room designed for this kind of tests. The tests were arranged according to their stressfulness where the less stressful test was conducted first, followed by novel object recognition (NOR) from days 40 to 42, and finally the more stressful test was conducted. Morris water maze (MWM) test was conducted from days 38 to 42.
After conducting all the tests, the rats were euthanized by cervical dislocation. The brains of the rats were detached immediately; the hippocampi were detached on ice plates and then washed out by cold normal saline. Finally, they were divided into three sets (n = 6 rats each) and stored for analysis at −80°C.
The first set of hippocampi were homogenized in phosphate-buffered saline for biochemical analysis of reduced glutathione (GSH), malondialdehyde (MDA), acetylcholine esterase (AChE), BDNF, and caspase 3 (Casp-3). The second set was, however, used for the determination of tyrosine receptor kinase B (TrKb), mitogen-activated protein kinase (MAPK), extracellular signal-regulated kinases 1/2 (ERK1/2), and CREB using Western blot analysis. Finally, the third set was used for the determination of Bcl-2 associated X protein (Bax) and B-cell leukemia/lymphoma-2 (Bcl-2) expressions using quantitative real-time polymerase chain reaction (RT-PCR).
Behavioral tests
Morris water maze
MWM test was carried out to estimate the spatial learning and memory function. 22 Water maze experiments were executed in a circular stainless tank of 150 cm in diameter × 50 cm in height, half filled with water mixed with dry milk to make it opaque. The pool was divided into four quadrants. An invisible platform (10 cm in diameter) which was 2 cm below the surface of the water was placed in the same target quadrant of the training session.
Each rat was given four daily training trials to find and climb on the platform for 4 consecutive days. Each trial began with placing the rat gently in the pool at different starting positions equally spaced around the perimeter of the pool and then the escape latency was calculated, which is the time taken by the rat to find the platform. The rat was then given a rest period on the platform for 15 s. If rats failed within 60 s to find the platform, they were placed on it for 15 s. The escape latency for each rat (four different positions) was averaged and then the average of each group was calculated per day.
On the fifth day, the platform was removed to perform the probe test, and a single 60-s trial was carried out for each rat. The time spent in the target quadrant was recorded, which indicates the extent of memory function. A digital camera was used to record the rats’ motion during the maze.
Novel object recognition
NOR test was performed to assess object recognition memory. It consists of three phases: habituation, familiarization, and test phase. 23 A no-lid box made of wood with dimensions 70 × 70 × 30 cm3 was used to perform the experiment.
On the first day, each rat was subjected to the session-habituation phase for 3 min when rats were allowed to move freely without any obstacle in the box. The box was then continuously cleaned after each rat’s session with alcohol (70%).
On the second day, each rat entered the familiarization phase in which two identical objects in color and size were placed in the box. The rat was allowed to get familiar with these two objects for 3 min.
On the third day, the test phase, only one object was replaced by another object which was different in color, and each rat was allowed to get familiar with this new setup for 3 min. The time spent by the rat to discover the old object and the new object was named (T
f) and (T
n), respectively. A discrimination index was calculated by the following equation:
Biochemical parameters
Colorimetric assay
MDA and GSH levels and AChE activity were evaluated using the method of Mihara and Uchiyama, 24 Ellman, 25 and Ellman et al., 26 respectively. The method of Lowry 27 was used to estimate the protein content.
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay kit was used to determine the BDNF level and the Casp-3 activity (My BioSource, San Diego, California, USA; Cat. No. MBS824814 and MBS7244630, respectively) according to the provided procedure.
Western blot analysis
The proteins from the hippocampus tissues were homogenized in the radioimmunoprecipitation assay (RIPA) lysis buffer. The Lowry method was used to evaluate the total proteins. Protein samples were loaded into an individual lane in sodium dodecyl sulfate polyacrylamide gel electrophoresis and then transferred to polyvinylidene difluoride membranes. After blocking the membranes with the blocking solution (Tris-buffered saline with Tween 20 (TBST) buffer and 3% bovine serum albumin) at room temperature for 1 h, they were incubated overnight at 4°C with primary antibodies diluted in TBST against TrKb, MAPK, ERK1/2, and CREB. After washing, the membranes were incubated with horseradish peroxidase goat anti-rabbit–conjugated secondary antibodies (Novus Biologicals, Littleton, Colorado, USA) at room temperature for 1 h. The protein bands were shown up with enhanced chemiluminescence system (BIORAD, Hercules, California, USA). Finally, the band intensity of the target proteins was read using an image analysis software against the control sample by normalization to β actin on the ChemiDoc MP imager (BIORAD, Hercules, California, USA).
Gene analysis by quantitative RT-PCR
Using the SV Total RNA Isolation System (Promega, Madison, Wisconsin, USA), total RNA was extracted from the hippocampus tissue homogenates. The obtained RNA was verified for pureness spectrophotometrically at 260 nm. The extracted RNA was reversely transcribed to complementary DNA using RT-PCR kit (Applied Biosystem, Waltham, Massachusetts, USA). Quantitative RT-PCR of Bax and Bcl-2 was performed using SYBR Green PCR Master Mix (Qiagen, Germany) as described by the manufacturer and β actin was used as a housekeeping gene. The primer sequences are described in Table 1. The PCR amplifications were performed as follows: 10 min at 95°C for activation of AmpliTaq DNA polymerase, 40 cycles of denaturation at 95°C for 15 s, and annealing/extension at 72°C for 10 min. The relative expression of target genes was obtained using the 2−ΔΔCt formula. 28
The sequence of primers used for real-time PCR analysis.

Bcl-2: B-cell leukemia/lymphoma-2; Bax: Bcl-2 associated X protein; PCR: polymerase chain reaction; β actin: beta-action.
Statistical analysis
The GraphPad Prism software (version 5) was used to perform the statistical analysis. Normality and homogeneity tests were performed for all the data using D’Agostino–Pearson and Barlett’s tests, respectively. These data were then analyzed using one-way analysis of variance, followed by Tukey’s multiple comparison test. Statistical significance was accepted at a level of p < 0.05, and the results were expressed as mean ± standard error of the mean.
Results
Effect of lycopene on the cognitive performance in MWM and NOR
In MWM, the efficiency of the learning ability in BPA-intoxicated rats was decreased, there was a significant (p < 0.001) increase in escape latency from the second to the fourth day of training by 35%, 45%, and76%, respectively, and in the probe test, BPA intoxication decreased (p < 0.001) the time spent in the target quadrant by 66% as compared to the control group. Yet the coadministration of lycopene with BPA reverted the escape latency in the third and fourth day of training as well as the time spent in the target quadrant in the probe test to their normal values (Figure 1(a) and (b)).

Effect of lycopene on cognitive functions in the hippocampus of rats subjected to BPA intoxication in (a and b) MWM and (c) NOR. Data are the mean ± SEM (n = 18). # p < 0.05: significantly different from control, $ p < 0.05: significantly different from BPA, and *p < 0.05: significantly different from lycopene, using one-way ANOVA, followed by Tukey’s multiple comparison test. BPA, 50 mg/kg orally 3 days a week for 42 days and lycopene, 10 mg/kg orally daily for 42 days. BPA: bisphenol A; MWM: Morris water maze; NOR: novel object recognition; ANOVA: analysis of variance; SEM: standard error of the mean.
In NOR, there was a significant (p < 0.001) decline observed in the discrimination index in the BPA treated group as compared to the control group by 81%. However, in the lycopene + BPA group, there was a significant (p < 0.001) increase in the discrimination index by fourfold as compared to the BPA group (Figure 1(c)). Treatment with lycopene alone did not show any significant change in the training trials, nor the probe test in MWM or the discrimination index in NOR as compared to the control group.
Effect of lycopene on the oxidative stress markers
BPA-intoxicated rats provoked a significant (p < 0.001) reduction in the hippocampal GSH level by 53%, along with a fivefold rise (p < 0.001) in the hippocampal MDA level as compared to the control group. On the other hand, co-treatment of lycopene with BPA reverted their levels to the normal values. There was however no significant change in the oxidative stress markers of the group treated with lycopene alone compared to the control group (Table 2).
Effect of lycopene on oxidative stress markers in the hippocampus of rats subjected to BPA intoxication.a

MDA: malondialdehyde; GSH: reduced glutathione; BPA: bisphenol A; ANOVA: analysis of variance.
a Data are the mean ± SEM (n = 6). BPA, 50 mg/kg orally 3 days a week for 42 days and lycopene, 10 mg/kg orally daily for 42 days.
b Significantly different from control group at p < 0.05 using one-way ANOVA, followed by Tukey’s multiple comparison test.
c Significantly different from BPA group at p < 0.05 using one-way ANOVA, followed by Tukey’s multiple comparison test.
d Significantly different from lycopene group at p < 0.05 using one-way ANOVA, followed by Tukey’s multiple comparison test.
Effect of lycopene on TrKB, MAPK, ERK1/2, CREB, and BDNF
Herein, the BPA-intoxicated group showed a significant (p < 0.001) decrease in the hippocampal TrKB, MAPK, ERK1/2, and CREB protein expression by 75%, 83%, 82%, and 70%, respectively (Figure 2(a) to (d)), along with a significant (p < 0.001) decline in the hippocampal BDNF level by 46% (Figure 3(a)) as compared to the control group. On the other hand, concurrent treatment of intoxicated rats with lycopene caused a marked (p < 0.01) threefold increase in the protein expression of TrKB as compared to the BPA-intoxicated group and restored the remaining aforementioned parameters to their normal levels.

Effect of lycopene on protein expression of (a) TrKb, (b) MAPK, (c) ERK1/2, and (d) CREB in the hippocampus of rats subjected to BPA intoxication. Data are the mean ± SEM (n = 6). # p < 0.05: significantly different from control, $ p < 0.05: significantly different from BPA, and *p < 0.05: significantly different from lycopene, using one-way ANOVA, followed by Tukey’s multiple comparison test. BPA, 50 mg/kg orally 3 days a week for 42 days and lycopene, 10 mg/kg orally daily for 42 days. BPA: bisphenol A; TrKb: tyrosine receptor kinase B; MAPK: mitogen-activated protein kinase; ERK1/2: extracellular signal-regulated kinases 1/2; CREB: cAMP response element-binding protein; ANOVA: analysis of variance; SEM: standard error of the mean.

Effect of lycopene on (a) BDNF level and (b) AChE activity in the hippocampus of rats subjected to BPA intoxication. Data are the mean ± SEM (n = 6). # p < 0.05: significantly different from control, $ p < 0.05: significantly different from BPA, and *p < 0.05: significantly different from lycopene, using one-way ANOVA, followed by Tukey’s multiple comparison test. BPA, 50 mg/kg orally 3 days a week for 42 days and lycopene, 10 mg/kg orally daily for 42 days. BPA: bisphenol A; BDNF: brain-derived neurotrophic factor; AChE: acetylcholinesterase; ANOVA: analysis of variance; SEM: standard error of the mean.
Effect of lycopene on AChE activity
BPA administration significantly (p < 0.001) reduced the AChE activity in the hippocampus by 50% as compared to the control group, while concurrent treatment of intoxicated rats with lycopene significantly (p < 0.001) increased its activity by 57% as compared to the BPA group. Treatment of rats with lycopene alone did not affect the normal hippocampal AChE activity (Figure 3(b)).
Effect of lycopene on apoptotic/antiapoptotic markers
It has been observed that BPA intoxication caused a massive upregulation (p < 0.001) in the hippocampal proapoptotic gene Bax (5-fold), in addition to rush (p < 0.001) hippocampal Casp-3 activity (4-fold), accompanied by a significant decrease (p < 0.001) in the hippocampal antiapoptotic gene Bcl-2 (4.5-fold), as compared to the control group. However, the concurrent treatment of intoxicated rats with lycopene blunted (p < 0.001) Bax messenger RNA (mRNA) expressions as well as Casp-3 activity by 61% and 59%, respectively, as compared to the BPA-intoxicated group, while it upregulated Bcl-2 mRNA level to the level of the control group (Figure 4(a) to (c)).
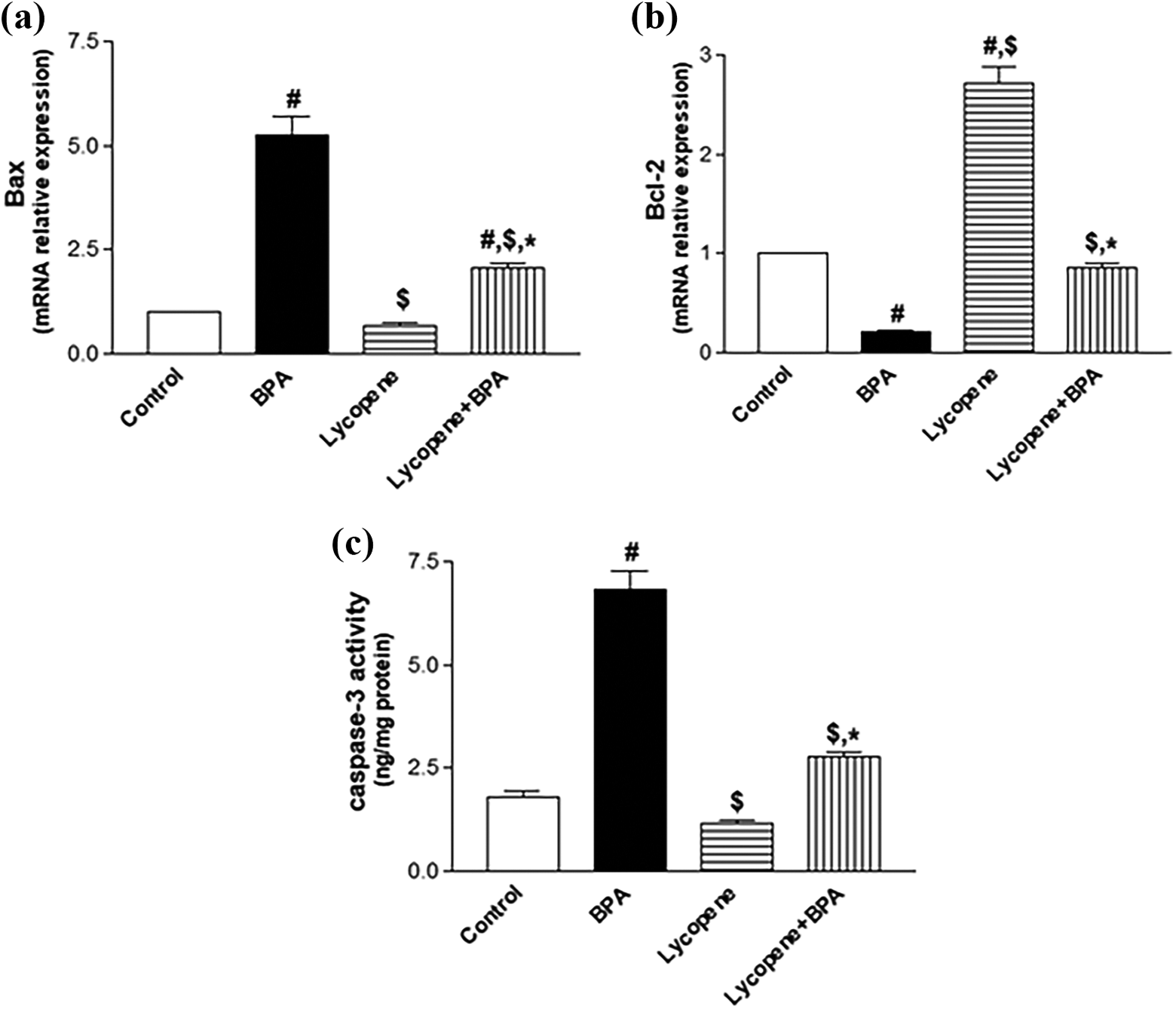
Effect of lycopene on apoptotic/antiapoptotic markers in the hippocampus of rats subjected to BPA intoxication. mRNA relative expression of Bax (a) and Bcl-2 (b) as well as (c) Casp-3 activity. Data are the mean ± SEM (n = 6). # p < 0.05: significantly different from control, $ p < 0.05: significantly different from BPA, and *p < 0.05: significantly different from lycopene, using one-way ANOVA, followed by Tukey’s multiple comparison test. BPA, 50 mg/kg orally 3 days a week for 42 days and lycopene, 10 mg/kg orally daily for 42 days. BPA: bisphenol A; Bax: Bcl-2 associated X protein; Bcl-2: B-cell leukemia/lymphoma-2; ANOVA: analysis of variance; SEM: standard error of the mean; mRNA: messenger RNA; Casp-3: caspase 3.
Discussion
To our knowledge, the present study is the first that shows the neuroprotective effect of lycopene against BPA intoxication in hippocampi of rats. Concurrent treatment of lycopene with BPA curbed oxidative stress by reverted MDA and GSH to their normal level. Activation of TrKB/MAPK/ERK1/2/CREB signaling pathway transcribed BDNF and its receptor TrKB. Moreover, CREB increased the antiapoptotic Bcl-2 that prevents apoptosis which was confirmed by the decrease of BAX and Casp-3 levels. The valuable protection effects of lycopene were reflected on the behavioral tests showing improvement in the impairment of learning and cognition memory induced by BPA in MWM and NOR tests accompanied by increasing in the AchE activity (Figure 5).
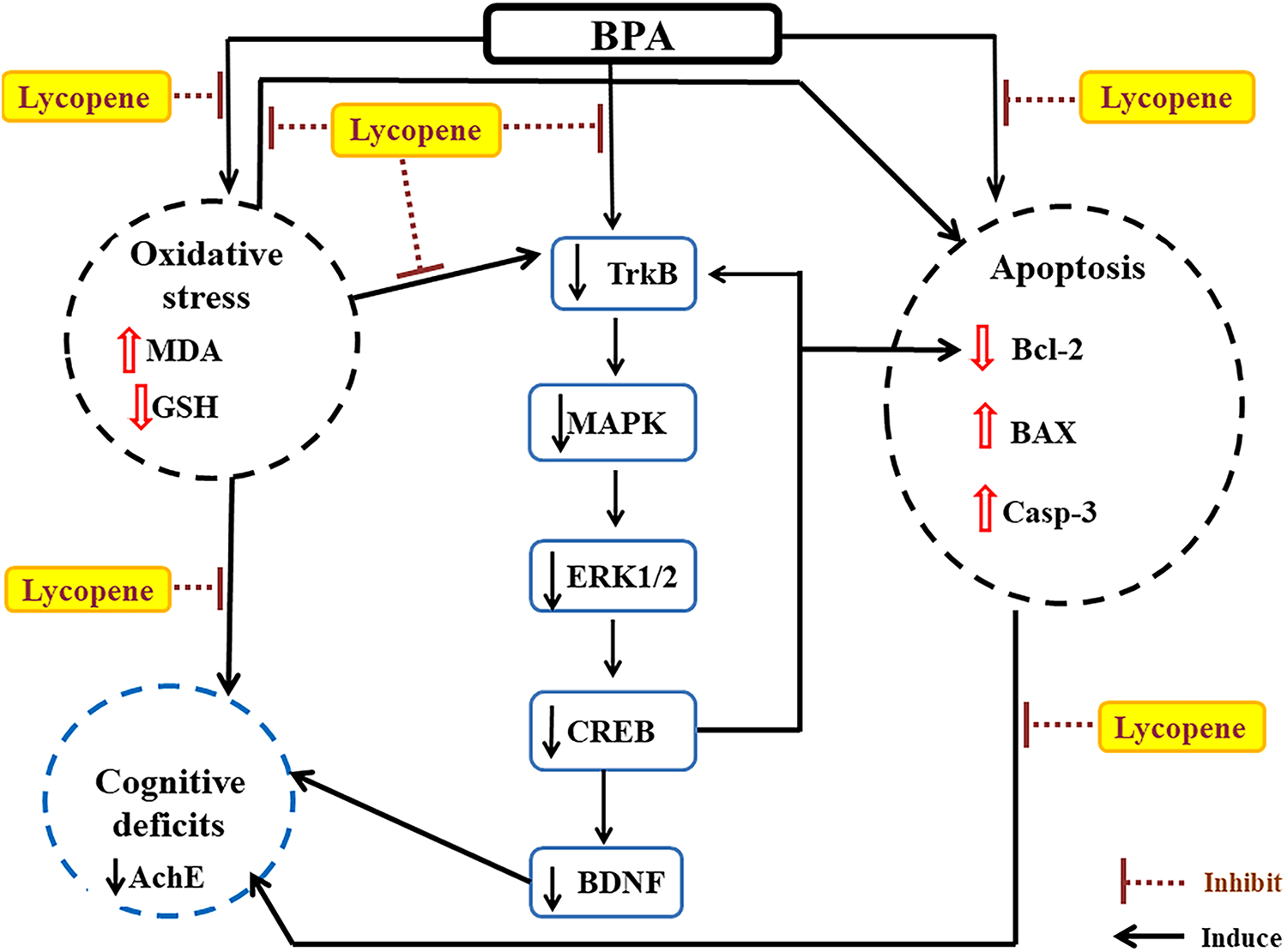
A sum-up of lycopene protection mechanisms in the hippocampus of rats subjected to BPA intoxication. BPA: bisphenol A; MDA: malondialdehyde; GSH: reduced glutathione; TrKb: tyrosine receptor kinase B; MAPK: mitogen-activated protein kinase; ERK1/2: extracellular signal-regulated kinases 1/2; CREB: cAMP response element-binding protein; BDNF: brain-derived neurotrophic factor; AChE: acetylcholinesterase; Bcl-2: B-cell leukemia/lymphoma-2; Bax: Bcl-2 associated X protein; Casp-3: caspase 3.
BPA intoxication disturbed the brain redox state by augmenting lipid peroxidation and lowering GSH in the rat hippocampus. These results are in harmony with previous studies in different brain regions of rats. 20,29 BPA has a lipophilic chemical structure, which can easily penetrate the blood–brain barrier and induce its toxicity through oxidative stress. 30 Moreover, the quinone and hydroquinone forms of BPA disturb the redox state in the rat striatum by the increased hydroxyl radical formation. 31 The brain is the most susceptible organ to oxidative injury, due to its high concentration of lipids, high rate of oxidative metabolism, and low level of intracellular antioxidant system. 32 In the present study, reactive oxygen species (ROS) induced by BPA stimulated polyunsaturated fatty acid oxidation and produced MDA as an essential product of lipid peroxidation which construct adducts with proteins and DNA bases leading to cellular damage. The decline in GSH level in the hippocampus of the BPA-intoxicated rats at the current study is due to it being consumed in scavenging hydroxyl radicals induced by BPA
Co-treatment of lycopene markedly counteracted the depletion in GSH level and the increase in MDA level induced by BPA and restored their levels to normal. These results were in agreement with previous studies that proved the antioxidant effect of lycopene in various oxidative stress conditions. 33,34 It has been reported that the antioxidant capacity of the carotenoids molecule increased by increasing the number of conjugated double bonds. Lycopene among all the natural carotenoids is the most efficient and easiest to be attacked by electrophilic entities with its 11 conjugated and 2 nonconjugated double bond showing high activity against ROS. 35 Lycopene also possess blood–brain barrier permeability. 36 This explains its ability in trapping ROS, guarding against the oxidative stress, and reducing the cellular components damage in the present study.
BPA intoxication showed a significant decline in the hippocampal protein expression of TrKB and its downstream molecules MAPK, ERK1/2, and CREB, as well as a marked decrease in the hippocampal BDNF level compared to the control group, indicating an inhibition of BDNF/TrKB system. These results are in agreement with Wang and his colleagues. 11 However, co-treatment of lycopene offered significant increases in TrKB protein expression while upregulating and restoring the other aforementioned parameters to their normal levels. Previous reports revealed that lycopene could increase BDNF. 34,37 BDNF is a member of the neurotrophins group, which is very important for brain neuronal health, markedly implicated in neuron development and differentiation, plasticity, and repair mechanisms. 38 The activation of TrkB by BDNF triggers the intracellular-signaling cascades including phosphatidyl inositol-3 kinase, phospholipase C-γ, and MAPK/ERK pathways, which are involved in neural survival, growth, and synaptic plasticity that helps in the process of learning and memory. 39,40 CREB is a transcription factor that has an important effect in neuronal survival, which additionally plays an important role in regulating learning and memory. 41 Phosphorylation of CREB is mediated by BDNF. 42 Moreover, phospho-CREB helps the transcription of BDNF and its receptor TrKB. 43,44 Many studies reported that oxidative stress can disturb synaptic plasticity and cognition in the brain by interacting with the BDNF system in several neurological disorder. 45 –47 Promoting and activating the aforementioned pathway in the present study by lycopene could probably be the result of its potent antioxidant properties that scavenge ROS and improve synaptic plasticity.
Next, we evaluated the protection of lycopene against apoptosis, by assessing its effect over the apoptotic and the antiapoptotic markers. In the present study, the coadministration of lycopene with BPA showed a marked decrease in the proapoptotic Bax gene expression, hampering Casp-3 to its normal level while upregulating and restoring the antiapoptotic Bcl-2 gene expression to its normal level in the hippocampus of rats. A previous study indicated that BPA induces apoptosis via Bax upregulation, Bcl-2 downregulation, and Casp-3 activation. 48 BPA accumulated into the mitochondrial membrane due to its lipophilic character, inducing mitochondrial dysfunction, inhibiting both nicotinamide adenine dinucleotide (NAD)- and flavin adenine dinucleotide (FAD)-linked respiration, and uncoupling oxidative phosphorylation leading to a depleting mitochondrial intracellular ATP. 49 This mitochondrial dysfunction induced by BPA led to an overexpression of free radicals and an enhancement of downstream molecules in the mitochondrial apoptotic signaling pathway. This was revealed by the increase in Bax gene expression and the inhibition of Bcl-2 gene expression, ending with rising Casp-3 activation and resulting in hippocampal neural death. Recent studies proved the ability of lycopene to rebalance the proapoptotic and the antiapoptotic markers and restoring brain homeostasis. 33,50 Moreover, previous studies proved that the inhibition of Casp-3 activation could contribute to the antiapoptotic effect of lycopene in primary cultured rat hippocampal and cerebral cortical neurons. 51,52 The observed antiapoptotic effect of lycopene may be attributed to its powerful antioxidant properties in scavenging free radicals that induced neuronal apoptosis. Moreover, the lycopene antiapoptotic effect may be referred to its ability to increase the protein expression of CREB, as many previous studies indicated that CREB is upstream of Bcl-2 and that CREB phosphorylation could positively regulate the antiapoptotic Bcl-2 which stabilizes the integrity of the mitochondrial outer membrane and could inhibit apoptosis. 53 Furthermore, Bcl-2-overexpressing cells exhibit elevated levels of cellular GSH. 54
It is well-known that neuronal networks in the hippocampus play an important role in learning and memory and any deterioration in these networks will affect the cognitive performance. 55 In the present study, administration of BPA caused a marked prolongation in the escape latency in the daily training trials and a decrease in the time spent in the target quadrant of MWM test, as well as a decline in the discrimination index of NOR test, which indicated impairment in acquisition and retention of spatial memory and cognition deficits. These results were in accordance with El Tabaa et al. 56 In addition, a previous study reported that BPA impaired object recognition in adult male rats by blocking synaptic plasticity processes in the hippocampus. 57 It has been reported that BDNF plays a critical role in memory consolidation. 58,59 Wang and colleagues 11 also proved that inhibiting ERK1/2/CREB/BDNF pathway in the hippocampus is related to the impairment of object recognition memory. The impairment in object recognition memory and spatial learning observed in the present study may be due to two reasons: firstly, the state of oxidative stress induced by BPA intoxication. This finding is supported by Massaad and Klann 60 who have proclaimed that the structures and functions of many enzymes and proteins that were involved in cell signaling could be inactivated by oxidative stress, leading to synaptic plasticity deterioration, neural cell death, and, consequently, memory loss. Secondly, it may be due to the decline in hippocampal BDNF and CREB levels, which are involved in the learning and memory function.
This reduction in cognitive performance induced by BPA is accompanied by a decline in hippocampal AChE activity. This result has been consistent with a former study conducted by Fan and colleagues. 61 Miyagawa and colleagues found that BPA caused memory impairment associated with a marked decrease in cholinergic fibers in the hippocampus of prenatal and neonatal mice. 62 This reduction may be attributed to the case of oxidative stress caused by BPA, as Wyse et al 63 reported that oxidative stress could inhibit AChE activity. Moreover, BPA inhibited synaptogenesis in hippocampus. 64 Therefore, the reduction in AChE in the present study may be a consequence of the reduction in acetylcholine production due to reduction in cholinergic fibers induced by BPA in the hippocampus of rats. A previous study reported that the functional state of the cholinergic neurons could be monitored by measuring the AChE activity, and the acetylcholine system definitely plays a critical role in regulating the major functions of hippocampus. 61
Lycopene co-treatment with BPA showed an improvement in cognitive functions revealed by normalizing the escape latency as well as increasing the time spent in the target quadrant of MWM and the discrimination index in NOR test. This was in accordance with previous studies which proved that lycopene could ameliorate the impairment in learning and memory induced by different models. 37,65 Lycopene co-treatment also showed a marked increase in the AChE activity. This can be explained by the ability of lycopene to scavenge ROS and increase BDNF/TrKB system, which is an important modulator for neurons plasticity and synaptogenesis. This led to an increase in cholinergic fibers, an enhanced ACh releasing, and, consequently, an increased AChE activity. A previous study supported this reasoning and indicated that BDNF could increase Ach releasing in the hippocampus of rats. 66
Conclusion
The present study revealed that lycopene was able to protect against neurotoxicity induced by BPA in the hippocampus, which was mediated by the inhibition of oxidative stress, improving TrKB/MAPK/ERK1/2/CREB/BDNF signaling pathway, the suppression of neural apoptosis, and, consequently, boosting synaptic plasticity along with reversing the cognitive impairment in rats. Further studies will be needed to elucidate the full molecular mechanisms beyond the neuroprotection of lycopene in BPA intoxication.
Footnotes
Acknowledgment
The authors would like to thank Dr Laila A Rashed, Professor of Medical Biochemistry Department, Faculty of Medicine, Cairo University, Egypt, for supplying us with the needed equipment to perform the biochemical analysis.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Ethical approval
All applicable international, national, and/or institutional guidelines for the care and use of animals were followed.
Funding
The author(s) received no financial support for the research, authorship, and/or publication of this article.