
Abstract
Endometriosis is characterized by the presence of functional endometrial tissue in other pelvic organs. This gynecologic problem occurs in 35–50% of women with pain and infertility. Endometriotic cells share some characteristics such as proliferation, migration, and invasion with tumor cells. Pyrvinium pamoate, an anthelmintic drug approved by the Food and Drug Administration, could inhibit the Wnt/β-catenin signaling pathway and its anticancer effects were examined by several researchers. In this study, 12 ectopic and eutopic endometrial biopsies from females with ovarian endometrioma and 12 endometrial biopsies from nonendometriotic females were obtained. Ectopic (EESCs), eutopic (EuESCs), and control (CESCs) endometrial stromal cells were isolated. Then, the effect of pyrvinium pamoate on the proliferation and invasiveness of in vitro cultured cells was evaluated. The proliferation of CESCs, EuESCs, and EESCs was significantly decreased after treatment with pyrvinium pamoate. In addition, treatment with pyrvinium pamoate significantly inhibited the invasiveness of CESCs, EuESCs, and EESCs compared to nontreated groups. The results of the present research showed that pyrvinium pamoate inhibits the proliferation and invasion of human endometriotic stromal cells in vitro, further investigations on the therapeutic potential of this compound in endometriosis are required.
Introduction
Endometriosis is defined as the growth of functional endometrial tissue outside of the uterus. Approximately 10–20% of reproductive-aged women are affected by this disease worldwide. 1 Endometriosis commonly engages pelvic organs, such as the ovaries, fallopian tubes, and peritoneum. Pelvic pain, dysmenorrhea, dyspareunia, and infertility are the most frequent symptoms. 2 According to Sampson’s hypothesis, which is generally accepted, endometriotic lesions are detectable on irregular target locations in the abdominal area due to the distribution of endometrial fragments during menstrual periods in the fallopian tubes. 3,4 Endometriotic cells exhibit invasive, proliferative, and adhesive features that contribute to the pathogenesis of endometriosis. 5 Cell differentiation, proliferation, migration, invasion, survival, and apoptosis are known to be regulated by the Wnt signaling pathway. 6,7 According to the World Health Organization histological classification of ovarian tumors, endometriosis has been newly classified as tumor-like lesions. 8 Current treatment options for endometriosis are hormonal therapies, such as contraceptives, gonadotropin-releasing hormone agonists, GnRH antagonists, progestins, and danazol, or laparoscopic surgery. 9 Hormonal therapy used by patients with endometriosis may interfere with the reproductive cycle and may result in infertility. 10
Deregulation of Wnt signaling contributes to the progression of various types of cancers and endometriosis. 11 –13 Recently, novel antagonists that interfere with the Wnt signaling pathway have been introduced as a promising means of medical management of endometriosis. 14 Pyrvinium pamoate, an oral FDA-approved antihelminthic drug, effectively inhibits WNT signaling and thereby increases β-catenin degradation by activation of casein kinase 1 alpha (CK1α). 15 An in vitro inhibitory effect of this compound on several types of cancers has been previously demonstrated. 16,17 Wiegering et al. found that intraportal injection of pyrvinium pamoate in nude mice inhibits the in vivo growth of tumors and in vitro migration of HCT116 colon cancer cells. 18 A previous study has demonstrated that PKF 115-584 and CGP049090 (fungal-derived products) attenuate proliferation, migration, and invasion of in vitro cultured endometriotic epithelial and stromal cells by targeting the Wnt/β-catenin pathway. 19 Zhang et al. reported an antiproliferative effect of pyrvinium pamoate on lung cancer stem cells. 20
Based on the proven inhibitory effect of pyrvinium pamoate on the Wnt signaling pathway, we evaluated the effects of pyrvinium pamoate for the first time on proliferation and invasion of in vitro cultured endometrial stromal cells (ESCs) from ectopic endometrioma (EESCs), eutopic endometrium from endometriotic patients (EuESCs), and endometrium of nonendometriotic controls (CESCs).
Materials and methods
Sample collection
Eutopic and ectopic endometrial biopsies were obtained from 12 women with ovarian endometrioma (aged 29–43 years) undergoing laparoscopy, and endometrial tissues from 12 control women with benign gynecological disorders lacking any type of endometriosis, as evaluated by laparoscopic surgery, were also collected. Eutopic and control endometrial biopsies of the functional layer were obtained from several spots of the fundal part of the uterine cavity using a biopsy catheter (Pipelle® Endometrial Suction Curette, Cooper Surgical, Connecticut, USA).
All patients had regular menstrual cycles and received no hormonal medication for at least 3 months before undergoing surgery with no previous malignancy or autoimmunity. All collected biopsy samples were in the early secretory phase of the menstrual cycle (days 16–19) based on the timing of the last menses, which was confirmed by histopathologic evaluation.
Stage III was determined in patients with endometrioma according to the revised American Fertility Society system. The present study was approved by the Ethics Committee of Shahid Beheshti University of Medical Sciences (IR.SBMU.MSP.REC.1395.589), and all enrolled participants provided written informed consent. Collected biopsies were immediately placed in Dulbecco’s Modified Eagle Medium: Nutrient Mixture F-12 containing 1% penicillin-streptomycin and transferred on ice to the laboratory.
Cell isolation and culture
Isolation and culture of ESCs were performed according to a previously published protocol. 21 In brief, after cutting the endometrial biopsies from women with ovarian endometrioma and control individuals into small pieces, digestion was performed using 0.1% collagenase I (Gibco, New York, New York, USA) for 1 h at 37°C. Removal of epithelial cells was performed by passing the cell suspension through a 70-µm cell strainer followed by a 40-µm cell strainer. Isolated single cells were then cultured in T25 flasks with DMEM/F12 medium supplemented with 10% fetal bovine serum and 1% penicillin-streptomycin (Gibco, Grand Island, New York, USA), and cells were incubated at 37°C in 5% CO2 for 12 h.
Purification and characterization of ESCs from all sources were assessed by immunofluorescent staining and flow cytometry using antibodies against vimentin and cluster of differentiation10 (positive markers) as well as cytokeratin and CD34 (negative markers).
Measurement of pyrvinium pamoate cytotoxicity
To determine the effective and safe concentration, ESCs were treated with 10, 1, 0.1, and 0.01 µM of pyrvinium pamoate (Sigma, New York, New York, USA) dissolved in 0.25% dimethyl sulfoxide for 24, 48 and 72 h. The sham group received only DMSO at a concentration of 0.25%, which is nontoxic to cultured cells. The experiment was performed in triplicate and repeated at least three times. The half-maximal inhibitory concentration (IC50) values using a nonlinear regression curve were calculated by the GraphPad Prism® 6.0 (GraphPad Software, San Diego, California, USA) software.
Cell proliferation assay
The 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide proliferation assay was used to assess the proliferative behavior of ESCs. For this purpose, 5 × 104 cells per well in triplicate were seeded in 24-well culture plates (SPL, Seoul, Korea) in 500 µL of DMEM/F12 medium (Gibco, Grand Island, New York, USA) containing 10% FBS. The culture plates were incubated at 37°C in 5% CO2 for 24 h to allow cell attachment. On the next day, experimental group treated for 72 h with 1.1 µM pyrvinium pamoate, sham and control groups received DMSO (at a concentration of 0.25%) and no treatment, respectively. After 72 h, culture medium of all culture wells was removed and replaced with 450 µL of fresh DMEM/F12 medium containing 10% FBS, then 50 µL of MTT solution (final concentration of 0.5 mg/mL) was added to all culture wells and cells were incubated for an additional 4 h. Subsequently, the content of each well was removed, and 200 µL of DMSO was added to solubilize the formazan crystals. Culture plates were incubated for 5 min at 37°C, and 100 µL of supernatant was then transferred to a 96-well enzyme-linked immunosorbent assay plate, which was read with Synergy™ HTX Multi-Mode Microplate Reader (BioTek Instruments, Winooski, Vermont, USA) at 470 nm.
Flow cytometric annexin V and PI staining assay
In brief, cells were cultured in six-well culture plates (1 × 105 cells per well) before treatment with pyrvinium pamoate. Cells were subsequently treated with the IC50 value of pyrvinium pamoate (1.1 µM) for 72 h. Cells were trypsinized and washed twice in cold PBS. After staining with fluorescein isothiocyanate-labeled annexin V and propidium iodide (PI), stained cells were analyzed (in duplicates) by the BD FACSCanto II (Becton Dickinson, Waltham, Massachusetts, USA).
Cell invasion assay
Invasiveness of EESCs, EuESCs, and CESCs was assessed using the Cultrex® 96 Well Cell Invasion Assay kit (R&D System, Minneapolis, Minnesota, USA). Cultured cells were starved in serum-free medium for 24 h to allow expression of free receptors. Briefly, FBS-starved nontreated cells or cells pretreated for 72 h with 1.1 µM pyrvinium pamoate were seeded in duplicate at a density of 5 × 104 in 50 µL of serum-free medium in basement membrane extract-coated insert wells (8 µm pore size), and 150 µL of 10% FBS medium was added to the bottom chamber as a chemoattractant. Cells were incubated at 37°C in a CO2 incubator for 24 h. After incubation, the top and bottom chambers were aspirated and washed with Wash Buffer. The top chambers were then transferred to the bottom chambers of the assay plate containing Cell Dissociation Solution/Calcein-AM, and they were incubated at 37°C in a CO2 incubator for 1 h. The top chambers were then removed, and the plate was read using a Synergy HTX Multi-Mode Microplate Reader at 485 nm excitation and 520 nm emission. Finally, the number of invaded cells (through the BME) was quantified using a standard curve.
RNA extraction and real-time reverse transcriptase-polymerase chain reaction
After a 72 h treatment of cultured ESCs with 1.1 µM pyrvinium pamoate and DMSO, total RNA was extracted using RiboEx (GeneAll, Seoul, Korea) according to the manufacturer’s instructions. The concentration and purification of extracted RNA were measured by a Synergy HTX Multi-Mode Microplate Reader. Reverse transcription was performed using 500 ng of total RNA and a complementary DNA Synthesis kit (Qiagen, Hilden, Germany). Real-time polymerase chain reaction (PCR) was performed in triplicate with SYBR® Green Master Mix (Ampliqon, Odense, Denmark) using a StepOnePlus (Applied Biosystems, Foster City, California, USA) thermal cycler. The glyceraldehyde-3-phosphate dehydrogenase housekeeping gene was used as an internal control for the normalization of gene expression. The primer sequences and amplicon sizes are shown in Table 1. The PCR cycling protocol was as follows: 95°C for 10 min; and 40 cycles of 95°C for 15 s and 60°C for 40 s. Analysis of real-time PCR data was done using the ΔΔCT method and REST-RG software (version 3). The negative control included a reaction mixture of water, SYBR Green, and primers.
List of primer sequences used in real-time PCR.
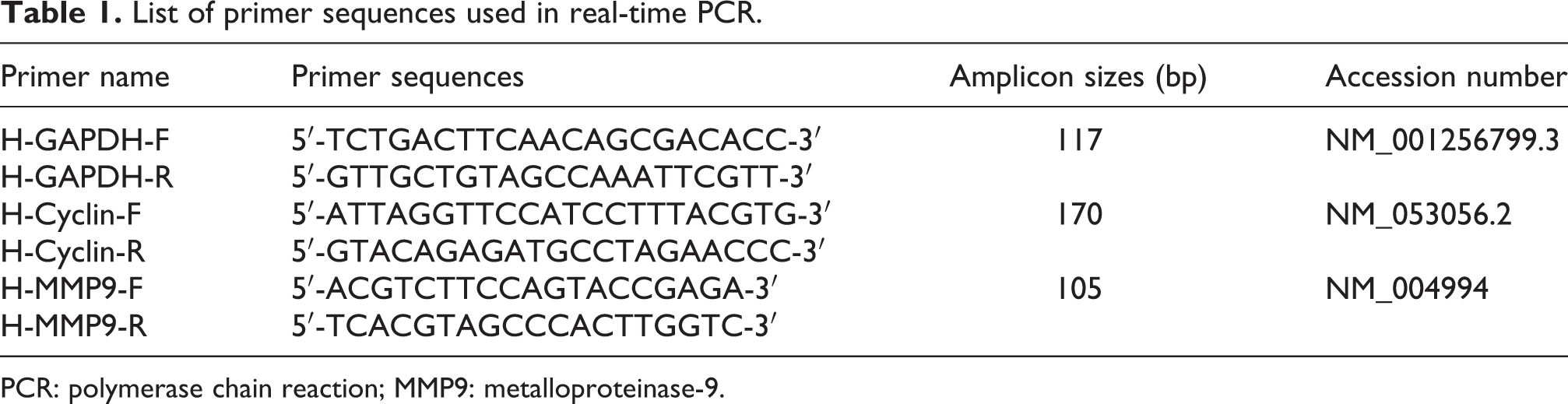
PCR: polymerase chain reaction; MMP9: metalloproteinase-9.
Statistical analyses
Results were expressed as the mean ± standard deviation. One-way analysis of variance (ANOVA) was used for comparing proliferation, invasion and messenger RNA (mRNA) expression between different groups, and a two-way ANOVA was used for comparisons of different concentrations of pyrvinium pamoate. Data were statistically analyzed using SPSS Program version 19 (Chicago, Illinois, USA). p Value < 0.05 was considered statistically significant.
Results
Characterization of isolated ESCs
The immunofluorescent and flow cytometry analyses of ESCs from all origins in the third passage demonstrated the expression of vimentin (stromal cell cytoskeletal marker; Figure 1(a)) but no expression of cytokeratin (epithelial marker; Figure 1(b)). Flow cytometric evaluation of ESCs showed that more than 94% of cells were positive for the CD10 marker (Figure 1(c)), indicating the purity of isolated stromal cells without any contamination with epithelial and endothelial cells.
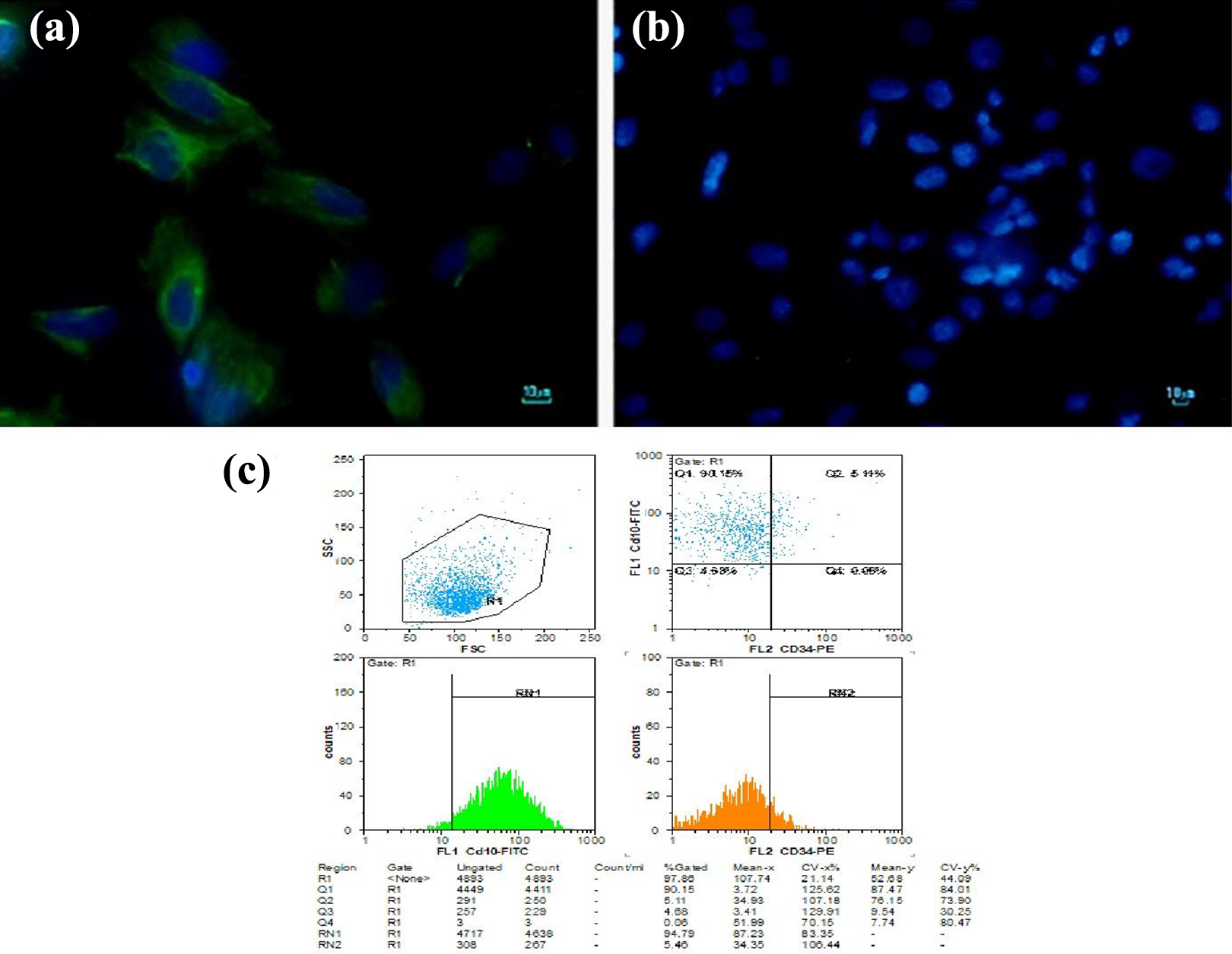
Immunofluorescent staining and flow cytometry evaluation were performed to characterize CESCs, EuESCs, and EESCs isolated from endometriotic and nonendometriotic subjects. (a) Vimentin-positive cells. (b) Cells lacking expression of cytokeratin. (c) Flow cytometric immunophenotyping of ESCs from all origins by expression analysis of CD10 (positive) and CD34 (negative) markers. ESC: endometrial stromal cell.
Cell viability assay and IC50 calculation
We assessed the viability of in vitro cultured human ESCs at 10, 1, 0.1, and 0.01 µM concentrations of pyrvinium pamoate in three time points (24, 48, and 72 h). The IC50 values of pyrvinium pamoate at 24, 48, and 72 h were 3.5 µM (confidence interval (CI): 1.589–8.008), 2.6 µM (CI: 1.193–5.937), and 1.1 µM (CI: 0.4542–2.998), respectively. Statistical comparisons between these three IC50 values did not show any significant difference (p = 0.1646; Figure 2). We used the 72 h period of incubation and its IC50 (1.1 µM) for further study.

The IC50 values of different concentrations of pyrvinium pamoate (10, 1, 0.1, and 0.01 μM) were determined on in vitro cultured human ESCs after incubation for three periods of time (24, 48, and 72 h) using GraphPad Prism® 6.0. Each point was in triplicate and represented as mean ± SD. IC50: half-maximal inhibitory concentration; ESC: endometrial stromal cell; SD: standard deviation.
Effect of pyrvinium pamoate on ESC proliferation
The comparison of the proliferation rates of CESCs, EuESCs, and EESCs before treatment indicated that there was a significant difference between the EESC group and CESC and EuESC groups (p < 0.05; Figure 3(a)).To determine the antiproliferative capacity of pyrvinium pamoate at a dosage of 1.1 µM for 72 h on the proliferation of CESCs, EuESCs, and EESCs, the MTT proliferation assay was performed. Pyrvinium pamoate significantly reduced the proliferation of CESCs (p < 0.001; Figure 3(b)), EuESCs (p < 0.001; Figure 3(c)), and EESCs (p < 0.05; Figure 3(d)) compared to the nontreated groups.
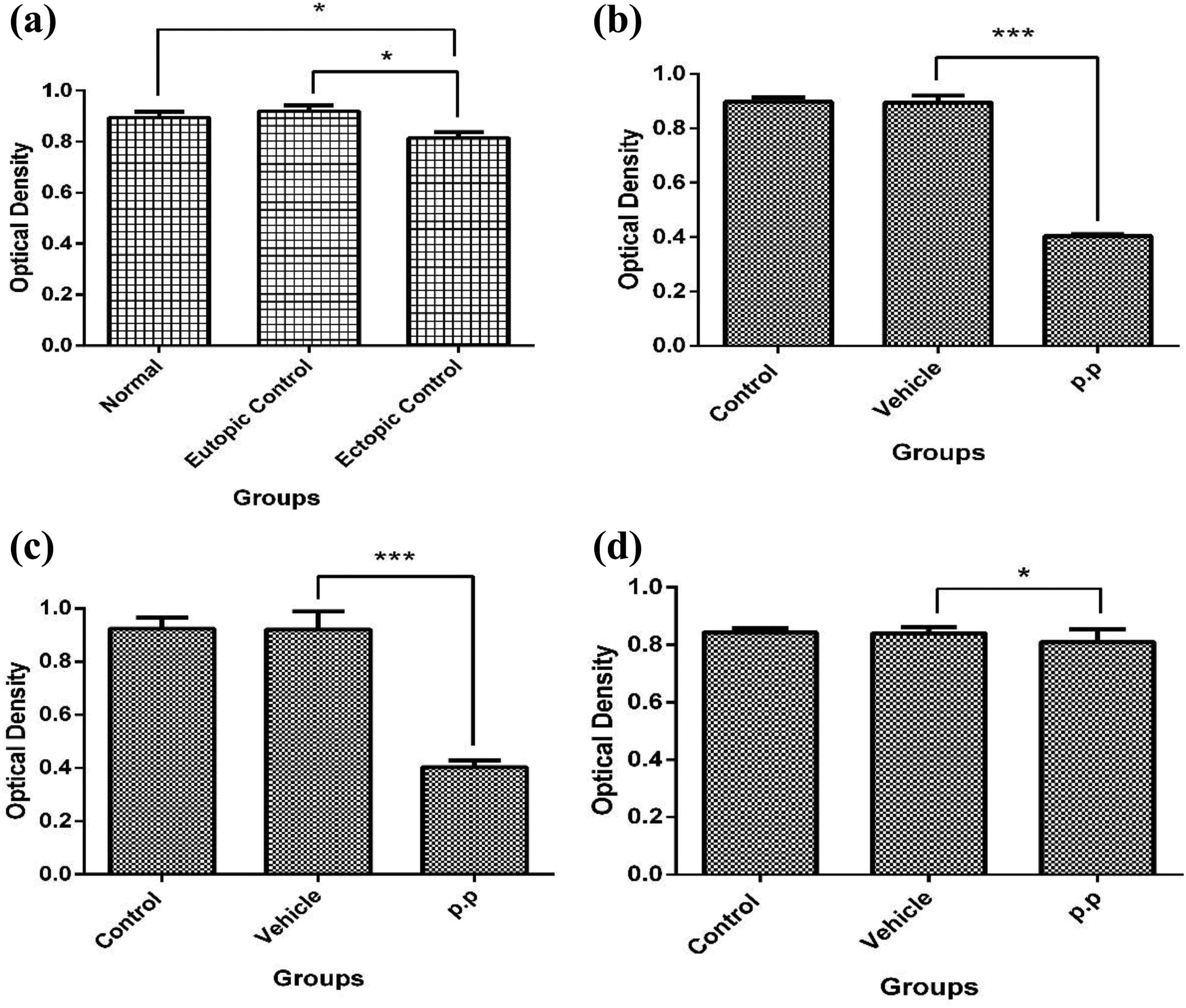
Effects of 1.1 μM PP treatment for 72 h on proliferative behavior of cultured CESCs, EuESCs, and EESCs. MTT assay was performed before (a) and after drug treatment on isolated normal (b), eutopic (c), and ectopic (d) stromal cells from endometriotic and nonendometriotic subjects OD. *p < 0.05 and ***p < 0.001. PP: pyrvinium pamoate; OD: optical density.
Effect of pyrvinium pamoate on apoptosis and necrosis
ESCs were analyzed for annexin V (apoptosis marker) and PI (necrotic cell marker). Approximately 96% of cells were negative for annexin V and PI, which confirmed the growth retardation of cultured cells induced by pyrvinium pamoate and confirmed its toxic effect (Figure 4).

Effects of 1.1 μM PP treatment for 72 h on apoptosis and necrosis of cultured CESCs, EuESCs, and EESCs. PP: pyrvinium pamoate.
Effect of pyrvinium pamoate on ESC invasion
The invasiveness of human ESCs isolated from all three origins was compared, and we found a significant difference between the EESC and EuESC groups (Figure 5(a); p < 0.05). The effect of pyrvinium pamoate on the invasive behavior of EESCs, EuESCs, and CESCs was assessed by their capacity to digest and invade through the basement membrane (8 µm pores). In the pyrvinium pamoate-treated groups, the invasiveness of CESCs (Figure 5(b)), EuESCs (Figure 5(c)), and EESCs (Figure 5(d)) was significantly decreased compared to nontreated groups (p < 0.05 and p < 0.01). Interestingly, a greater reduction of invasion features was observed after treatment with 1.1 µM pyrvinium pamoate for 72 h in the EESC group (Figure 5(d); p < 0.01).

Effects of 1.1 μM PP treatment for 72 h on invasive behavior of cultured CESCs, EuESCs, and EESCs. Cell invasion assay was performed before (a) and after drug treatment of purified normal (b), eutopic (c), and ectopic (d) stromal cells from endometriotic and nonendometriotic individuals. The results are represented as the mean ± SD. *p < 0.05 and **p < 0.01. PP: pyrvinium pamoate; SD: standard deviation.
Effect of pyrvinium pamoate on ESC Cyclin D1 and MMP9 expression
The mRNA expression of Cyclin D1 and metalloproteinase-9 (MMP9) was compared among nontreated groups. Furthermore, the effect of pyrvinium pamoate on Cyclin D1 and MMP9 expression (proliferation and invasion markers) was evaluated using real-time PCR. In the nontreated groups, the expression of Cyclin D1 was significantly increased in the EuESC group compared to the EESC group (p < 0.01; Figure 6(a)). However, no statistically significant difference was found when comparing Cyclin D1 expression in the EESC and EuESC groups to the CESC group. Pyrvinium pamoate had no significant effect on Cyclin D1 mRNA expression in the EuESC and CESC groups (Figure 6(b) and (c)), but Cyclin D1 mRNA levels were significantly decreased in the EESC group (p < 0.05; Figure 6(d)). Although the mRNA expression of MMP9 was higher in the EuESC group compared to both the EESC and CESC groups, this difference was not statistically significant (Figure 7(a)). The expression of MMP9 was significantly reduced in the CESC (p < 0.05; Figure 7(b)), EuESC (p < 0.01; Figure 7(c)), and EESC groups (p < 0.05; Figure 7(d)) after treatment with pyrvinium pamoate.
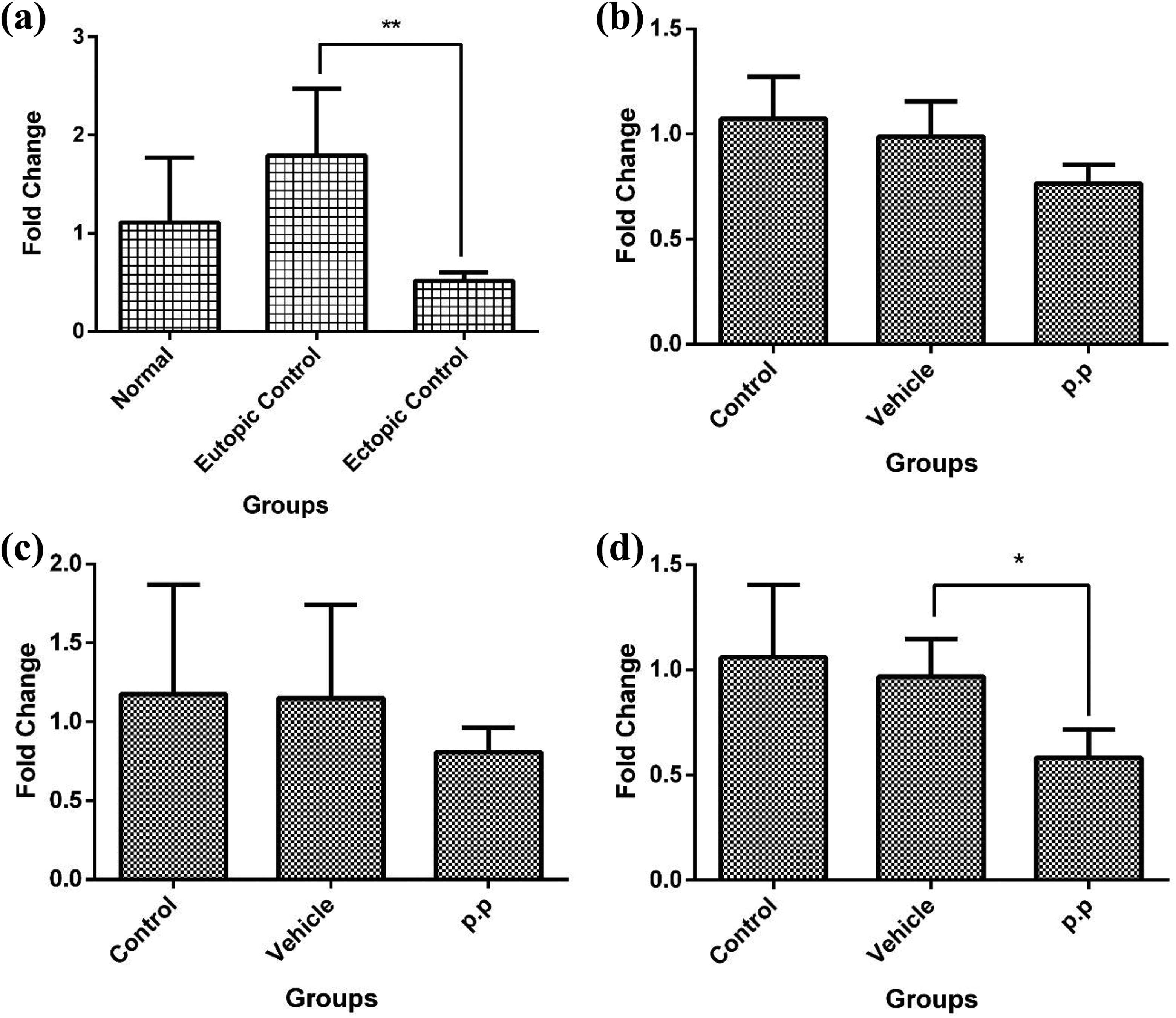
Effects of 1.1 μM PP treatment for 72 h on Cyclin D1 mRNA expression in cultured CESCs, EuESCs, and EESCs. Relative mRNA expression of Cyclin D1 was measured before (a) and after drug treatment of purified normal (b), eutopic (c), and ectopic (d) stromal cells from endometriotic and nonendometriotic individuals. The results are represented as the mean ± SD. *p < 0.05 and **p < 0.01. PP: pyrvinium pamoate; mRNA: messenger RNA; SD: standard deviation.

Effects of 1.1 μM PP treatment for 72 h on MMP-9 mRNA expression in cultured CESCs, EuESCs, and EESCs. Relative mRNA expression of MMP-9 was measured before (a) and after drug treatment of purified normal (b), eutopic (c), and ectopic (d) stromal cells from endometriotic and nonendometriotic individuals. The results are represented as the mean ± SD. *p < 0.05 and **p < 0.01. PP: pyrvinium pamoate; MMP-9; metalloproteinase-9; mRNA: messenger RNA; SD: standard deviation.
Discussion
Endometriosis is defined as the presence of endometrial tissue outside of the uterus that shows tumor-like behavior, such as proliferation and invasiveness. 1,22 Currently, available medications are mainly based on pain relief and reduction of estrogen levels through suppression of the pituitary-ovarian axis. Therefore, all of these scenarios impair the reproductive cycle. 2,23,24 Recently, many attempts have been made to identify novel compounds that do not interfere with the fertility potential of women with endometriosis and only target endometriotic tissue. Because abnormal activation of the Wnt/β-catenin signaling pathway has been confirmed in endometriosis 25 and some downstream Wnt target genes are involved in proliferation and invasive behavior, 15,26,27 inhibitors of this major signaling pathway may efficiently control the progression of endometriosis. 28,29 Pyrvinium pamoate blocks the Wnt/β-catenin signaling pathway through the direct activation of CK1α.
For the first time, the present study evaluated the effectiveness of pyrvinium pamoate at a dosage of 1.1 µM after 72 h of treatment on proliferation and invasion features of EESCs and EuESCs purified from endometriotic subjects. Pyrvinium pamoate is a Wnt signaling pathway inhibitor that may significantly alleviate the proliferation and invasiveness of isolated stromal cells. Aberrant activation of the Wnt/β-catenin signaling pathway in human endometriosis and cancer has recently been demonstrated in several studies. In accordance with this evidence, Matsuzaki and Darcha examined the effect of PKF 115-584 as a Wnt/β-catenin signaling pathway inhibitor on the behavior of endometriotic stromal and epithelial cells. 19 These authors showed that PKF 115-584 suppresses the invasion of endometriotic stromal cells. Moreover, the percent of inhibition induced by this compound in eutopic stromal cells purified from the endometrium of patients with ovarian endometrioma is higher than their ectopic counterparts. Elevated expression of matrix MMP-9 has been reported in the eutopic endometrium of endometriotic patients, thereby facilitating the invasion into target organs. 30 In the present study, EuESCs showed the highest invasiveness compared to the other two groups. Furthermore, pyrvinium pamoate significantly reduced the invasion of ESCs from all sources and exerted a greater inhibitory effect on the ectopic group.
Delbandi et al. reported that EuESCs show more invasiveness than control and ectopic groups, which agreed with the outcome of the present study, and they reported that cell proliferation is increased in the control group on either untreated or fibronectin-coated plates compared to EESCs and EuESCs. The proliferation of ectopic stromal cells is significantly lower than that of eutopic and control cells. 5 Similar results for the lowest proliferation level in ectopic stromal cells were found in our investigation. In contrast, another study has found that ESCs derived from eutopic endometrium display higher proliferative capacity than the control group. 31 This finding is compatible with our data regarding the increased proliferation of eutopic stromal cells in comparison to the ectopic and control groups. An explanation for the lower proliferative capacity of EESCs is that when shedding endometrial cells during the menstrual cycle migrate to form endometriotic tissue on a target organ, they exhibit slow growth and almost inactive behavior in terms of invasion. 5 The antiproliferative impact of pyrvinium pamoate on in vitro cultured lung cancer stem cells and multiple myeloma cell lines by targeting the Wnt signaling pathway has previously been elucidated. 17,20 Pyrvinium pamoate represses the growth and migration of colon cancer cells as well as the metastatic capacity of breast cancer stem cells in vitro by interfering with the WNT signaling pathway. 18,32 Similarly, in the present investigation, significantly reduced proliferation and invasion of in vitro cultured EESCs and EuESCs were observed following treatment with pyrvinium pamoate. In addition, the inhibitory effect of pyrvinium pamoate on the proliferation of ectopic cells was lower than that of eutopic and control cells, but no clear reason was found for this disparity.
Our results showed that pyrvinium pamoate effectively lowered Cyclin D1 and MMP9 mRNA expression in treated cells. According to previous studies, the Cyclin D1 and MMP9 genes play a critical role in cell proliferation and invasion and are regulated by the Wnt/β-catenin signaling pathway. 33 –35 Cyclin D1 in ESCs is reduced after treatment with the Wnt/β-catenin signaling pathway inhibitor, PKF 115-584. 19 Resveratrol, another Wnt/β-catenin signaling pathway suppressor, decreases mRNA levels of Cyclin D1 and MMP-9 in the human osteosarcoma U2-OS cell line. Additionally, the proliferation and invasion of U2-OS cells are reduced following treatment with a resveratrol compound. 36 After reverse transcriptase-PCR (RT-PCR) analysis, Goumenou et al. reported elevated expression of MMP-9 in endometriotic tissues compared to normal endometrium. 37 High expression of MMP-9 mRNA has also been reported in the 22-B human endometriotic stromal cell line. 38 MMP9 mRNA expression has been found in ESCs isolated from subjects with and without endometriosis. 39 Increased mRNA expression of MMP9 has been reported in ectopic endometrium from patients with ovarian endometriosis. 40 These results were not consistent with the present study, which found higher MMP9 mRNA expression in EuESCs of women with endometrioma compared to other groups. These discrepancies may partly be explained by different sources of endometrial tissues and cells as well as the stage of menstrual cycle and techniques for evaluation of Cyclin D1 and MMP9 expression. Collette et al. demonstrated no statistically significant difference in MMP-9 mRNA expression between normal and endometriosis patients following RT-PCR analysis. 30 This finding is consistent with our data, in which no significant difference in MMP-9 mRNA expression was detected between nontreated groups although the eutopic endometrium had higher MMP-9 mRNA expression levels.
The results of our research demonstrated that pyrvinium pamoate, an inhibitor of the Wnt signaling pathway, suppresses the proliferation, invasion behavior, and related mRNA expression in ESCs in vitro. Additional in vitro and animal research is needed to investigate the efficacy of this FDA-approved drug in the prevention and treatment of endometriosis.
Footnotes
Authors’ note
This article has been extracted from the thesis written by Mr Amin Karamian at the School of Medicine, Shahid Beheshti University of Medical Sciences, Tehran, Iran (registration no: 12). A-H Zarnani is also affiliated with Department of Immunology, School of Public Health, Tehran University of Medical Sciences, Tehran, Iran.
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The present study was financially supported by a grant from the Biology and Anatomical Sciences Department of the School of Medicine Shahid Beheshti University of Medical Sciences (Project No: 11104).