
Abstract
This study aimed to evaluate maternal toxicity, teratogenic, and placental oxidative effects resulting from the exposure of rats to crack cocaine smoke during pregnancy. Pregnant rats were exposed either to the smoke of crack and ashes (Crack) or to the smoke of ashes alone, nonexposed or pair-fed with the Crack group. Crack group was exposed to the smoke resulting from the burning of 250 mg of crack for 10 min, twice a day, from 7 days prior to mating until cesarian on gestational day 20. Placental oxidative stress and classical parameters of maternal and fetal evaluation were studied, in addition to the morphometric analysis of the fetal metamers. Even in the absence of changes in body weight gain and feed intake, crack altered the reproductive performance of dams. Exposure to the drug promoted late closure of the fetal fontanel. Furthermore, the morphometric study of the brain mass (BM)/skull cap ratio revealed a decrease in the BM of the fetuses exposed to the drug. Exposure to crack has an oxidative potential in fetal development, since exposure to the drug promoted placental lipid peroxidation. Our study showed that daily exposure to crack, even in lower frequency than that performed by users, has a teratogenic potential.
Introduction
The use of illicit drugs is a major concern for public health, particularly during pregnancy, due to its effects on the mother and on the fetus, along with postnatal consequences. The use of crack cocaine throughout the prenatal period is associated with several maternal and fetal complications, such as maternal hypertension with high rates of eclampsia, preterm birth, low birth weight, smaller cranial circumference, neurological and behavior changes, and, ultimately, fetal death and abortion. 1 –3
Crack cocaine is the free base form of cocaine hydrochloride, a potent central nervous system stimulant, and it is considered one of the most dangerous drugs due to its addictive potential. 4 This drug is freely obtained due to its low price and high availability, and it is easy to consume by smoking. Additionally, this way of exposure promotes a heightened effect that leads to abuse and addiction faster than other forms of cocaine. 5
Due to the potent dependence caused by crack cocaine, pregnant users often keep their compulsive use of the drug throughout pregnancy. Cocaine crosses the placenta through simple diffusion and, in addition to exposing the developing fetus, it causes a vasoconstrictive effect on the placenta that increases the chance of fetal hypoxia and the development of congenital abnormalities. 6 Besides, cocaine can stimulate uterine contraction and thereby cause miscarriage or premature birth. 7,8
However, little is known about the true toxic effects of crack in embryofetal development in pregnant women, since most users are inserted in a polydrug scenario and exposed to several interfering factors, such as malnutrition, social stress, behavioral factors, and the anorexic effect caused by cocaine. 1,9
Thus, the aim of the present study was to evaluate the effects of crack smoke on maternal reproductive parameters and on the embryofetal development of pregnant rats exposed to the drug during the gestational period. For the exposure, we used a device developed and validated by our research group that generates serum cocaine levels in rats similar to those observed in humans exposed to the same form of drug. 10
Materials and methods
Crack cocaine
Crack cocaine was obtained from legally seized drug trafficking products and was courtesy of The State Narcotic Division of São Paulo, pursuant to the court order of the Hon. Judge Lucas Pereira Garcia from the Forum of Itapecerica da Serra, São Paulo (proc. Number: 007417-59.2012.8.26.0268). The crack rocks were homogenized and tested for composition, presenting 63.7% cocaine.
Animals
Male and female Wistar rats, approximately 90 days old, were obtained from the Department of Pathology of the School of Veterinary Medicine and Animal Science—University of São Paulo (FMVZ-USP) and used according to the ethical principles in animal research of the Bioethics Committee of FMVZ-USP (process n°. 2351140916). The cages, with internal dimensions of 48.7 × 33.2 × 21.1 cm3 (length × depth × height), were placed in a room under controlled conditions of temperature (22°C to 24°C), relative humidity (50–65%), and light/dark cycle of 12/12 h.
Mating procedure
A total of 48 female and 24 male rats were used, with the objective of forming 4 experimental groups with 10 pregnant rats each. The time interval determined between the first and the last female to be pregnant did not exceed 10 days (2 estrus cycles). One male rat was placed with two females during the night period (6:00 p.m. to 7:00 a.m.). The day when sperm was found in the vaginal smear was designated as gestational day 0 (GD0). Before mating and after pregnancy diagnosis, female rats were kept in a ratio of two rats per cage, physically separated by a grid that allowed physical and visual contact between them and the evaluation of the feed intake of each animal.
The apparatus and crack exposure
The exposure apparatus, developed by Hueza et al., 10 consists of two chambers interconnected and separated by a hatch. Briefly, the animals are placed in an entrance chamber before the exposure and then transferred to the exposure chamber, where the smoke of 250 mg of burned crack cocaine is sucked in. The exposure chamber has the following internal dimensions: 34 × 21.5 × 22 cm3 (length × depth × height) and is made with crystal acrylic walls (polymethyl methacrylate—12 mm thickness). The burner is a metal pipe coupled to a glass connector that allows all the smoke produced to be sucked into the exposure chamber under −70 mmHg pressure. Since crack melts under the flame, a steel screen with cigarette ashes is placed inside the pipe, preventing the loss of the drug into the pipe, in the same manner employed by drug users. After the crack is burned and the equilibrium of the atmospheric pressure among chambers is reached, the hatch is opened, and the animals are introduced into the exposure chamber, which was designed to comfortably accommodate five adult rats per period of exposition.
Animal exposure
Four experimental groups with pregnant rats were set up as follows: a group exposed to the smoke of 250 mg of crack burned on a bed of cigarette ashes (Crack; n = 7 pregnant out of 12 placed to mate); a group submitted to the burning of cigarette ashes alone (Ash; n = 10 pregnant out of 12 placed to mate); a group inserted into the exposure box without any burning, aiming to mimic a possible stress of confinement (Stress; n = 9 pregnant out of 12 placed to mate); and a pair-fed group (Pf; n = 9 pregnant out of 12 placed to mate), composed of nontreated pregnant rats that received only the average amount of food consumed by the rats of the Crack group, which had food ad libitum, simulating any anorectic effect of cocaine. For that purpose, the amount of food consumed by the Crack group was recorded and used to determine the amount of diet given the following day to the Pf group, so that equal amounts were consumed by both groups. Rat exposure to the drug was initiated at least 7 days before GD0, and the last day of exposure was GD20.
The exposure was performed daily, twice a day with an interval of 12 h, for 10 min each. The drug here employed was the same used by Hueza et al., 10 thus, a single exposure of 250 mg of crack smoke for 10 min promotes serum levels of cocaine around 170 ng/mL. During the experimental period, the pregnant rats were observed daily in relation to clinical signs, food waste, morbidity, and mortality. Food consumption and body weight gain were measured every day from GD0, avoiding the period when the male was present.
Cesarean section, maternal, and fetal evaluation
On GD21, the pregnant rats were anesthetized (xylazine and ketamine, 5 and 50 mg/kg, respectively, intraperitoneally) and underwent cesarean. In the maternal evaluation, gravid and nongravid utero weight were measured; the number of corpora lutea, implantations, resorptions, live and dead fetuses, and embryo losses in the preimplantation and postimplantation period were recorded. The fetuses were weighed, measured, sexed, and examined for macroscopic external malformations. The placentas were collected, weighed, and immediately processed for oxidative stress measurements. After this procedure, half of each litter was separated for skeletal analysis and the other half for the visceral analysis. Morphometric analysis of the metameres obtained from visceral analysis was also performed.
Visceral and skeletal analysis
The incidence of external anomalies and visceral and skeletal changes in all fetuses was evaluated with a high contrast digital imaging stereo microscope for superficial and deep diaphanized tissues (Nikon® - model SMZ1000). After external analysis, half the fetuses were fixed in Harrison’s solution containing 95% ethanol, glycerol, 40% formalin, and 9% saline mixed in the proportions of 8:1:2.5:7.5 (v/v), respectively. Harrison’s solution was exchanged for 80% alcohol for 2 days and then for 90% alcohol. After 4 days, serial sections (transversal and coronal metameric segments) (Figure 1) were freehand performed with a razor blade as described by Wilson 11 for visceral examination. After visceral examination, skull cap (SC) metamers were placed into vials with formaldehyde previously identified until the morphometric analysis. The remaining fetuses were prepared for examination of the bones by the staining procedure of Staples and Schnell. 12 Briefly, the other half of pups were placed into acetone (99.5%) for 24 h. On the following day, gross malformations and the presence of cleft palate were evaluated and recorded. After that, evisceration was performed, and fetuses were washed with distilled water and placed into a 0.8% potassium hydroxide solution with 6 mg of alizarin that was changed every day for four consecutive days. After staining, fetuses were placed into a bleaching solution (ethanol, benzyl alcohol, and glycerin in the proportions 2:1:2, (v/v), respectively). Bleaching was considered perfect if the inner bones were white colored. Fetuses whose bones did not acquire this characteristic were discarded.

Fetus showing regions from which transverse and coronal sections were obtained: was 13 sections were performed, resulting in 14 regional metamers.
Morphometric analysis
The morphometric analysis of the transversal metamers was performed to complement the visceral analysis employing the Nikon Nis-Elements (Br)® software geared for acquisition and device control for standart research applications, requiring four-dimensional imaging. The metameric segment number 3 was placed on a petri dish, with its posterior side facing the lens of the stereoscopic zoom microscope, and was examined to evaluate the percentage of the brain mass (BM) by the internal SC area as follows: BM is calculated as the whole encephalon area (E) minus the ventricular area (V), when visible, and SC as the internal area of the SC (Figure 2). The cut line of the metamer was standardized as the inferior delimitation of the SC. This percentage was determined and calculated by the formula:


Metameric segment number 3 was used for morphometric evaluation. SC: skull cap; E: encephalon; V: ventricle (when visible) (magnification ×16).
The metameric segment number 14 is examined by its front side facing the lens of the stereoscopic zoom microscope for the determination of the proportions between height and width of the kidneys and adrenals.
Placental oxidative stress parameters
Around 0.1 g of placental tissue from each female was immediately homogenized in a solution containing 5% metaphosphoric acid and 0.6% sulfosalicylic acid (1:50 w/v). Samples were centrifuged and an aliquot of 20 µL of supernatant was added to 180 µL of 0.1 M potassium phosphate buffer with 5 mM ethylenediaminetetraacetic acid disodium salt, pH 7.5 for total glutathione (GSH) content evaluation. For oxidized glutathione (GSSG) determination, 20 µL of 2-vinylpyridine were added and incubated for 1 h to derivatize reduced GSH. 13 In both cases, an aliquot of 20 µL was added to 180 µL of a solution containing 5,5-dithio-bis-2-nitrobenzoic acid (DTNB), glutathione reductase (GR), and nicotinamide adenine dinucleotide phosphate (NADPH). DTNB reacts with reduced GSH produced by the action of GR and NADPH, forming 5-thio-2-nitrobenzoate, a chromophore that causes absorbance variation at 412 nm. 14 Results are calculated using a standard curve of GSSG.
The remaining placentas were homogenized with a buffered solution containing 140 mM potassium chloride and 10 mM monopotassium phosphate pH 7.0 (1:4 w/v) and centrifuged (3000 r/min, 20 min, 4°C). An aliquot of the supernatant was used in the lipid peroxidation assay. The remaining supernatant was further centrifuged (9000 r/min, 20 min, 4°C), and this second supernatant was used to determine the activities of the antioxidant enzymes: superoxide dismutase (SOD), 15 catalase, 16 and glutathione peroxidase (GPx), 17 all by spectrophotometric methods. Results are expressed in units/mg of protein. Protein content was assayed according to Lowry et al. 18
Lipid peroxidation was evaluated by malondialdehyde (MDA) reaction with thiobarbituric acid (TBA), producing the fluorescent adduct MDA-TBA2, which was assessed as described by Tanizawa et al. 19 Briefly, 100 μL of placenta homogenate were vigorously shaken with 500 μL 3% SDS, then added to acetate buffer (2 M, pH 3.6) and 0.8% TBA solution and incubated in water bath at 95°C for 1 h. After cooling, 1 mL of hydrochloric acid 0.2 N and 5 mL of n-buthanol:pyridine (15:1 v/v) were added to each tube, which was vigorously shaken for 2 min. The top layer was collected with a glass pipette and centrifuged (3000 r/min, 15 min). Fluorescence was read in a microplate reader (λ excitation 515 nm and λ emission 553 nm). Calculations were made using a standard curve for MDA and corrected by protein content.
Statistical analysis
Comparisons of the mean values between the experimental groups were determined by analysis of variance, followed by Tukey’s multiple comparison test. For reproductive performance, maternal measurements, and fetal evaluation, which may have inflated the type I error rate, a false discovery rate (FDR)-controlling procedure was employed. 20 Differences in proportions were calculated by the Fisher’s exact test. Placental oxidative stress parameters were analyzed with nonparametric Kruskal–Wallis test, followed by Dunn’s multiple comparison test. Data are expressed as mean ± standard deviation. Statistical differences were considered significant at p < 0.05.
Results
It is interesting to note that the pregnancy rate of females exposed to crack was 58.3%, versus 75% of the Stress and Pf groups, and 83.3% of the Ash group. Clinical signs of acute or subchronic toxicity in pregnant rats were not observed in any group throughout the experimental period.
From GD9 on, dams exposed to the burning of ashes (Ash group) presented a statistically significant increase in the food consumption compared to dams of the other groups (Figure 3). This reflected on the total feed intake, as shown in Figure 4(a), with a statistically significant increase in the food intake of Ash females when compared to the females of the Stress (p < 0.001) and Crack (p < 0.01) groups. It is important to remember that the Pf group consumed the same amount of food as animals from the group exposed to the drug (Crack), so no differences can be found between them.

Food consumption of pregnant rats exposed to or not to the smoke of 250 mg of crack cocaine during gestation period registered every 3 days. The Stress group mimicked the exposure by just entering the apparatus; the Ash group was exposed to the burning of ashes alone; and the Crack group was exposed to the crack and ashes smoke. Data are expressed as mean ± SD. *p < 0.05; Tukey’s multiple comparison test. SD: standard deviation.

(a) Total food consumption and (b) total body weight gain of female rats exposed or not to the smoke of 250 mg of crack cocaine, twice a day, from 7 days before mating until the 21st gestational day. The Stress group mimicked the exposure by just entering the apparatus; the Ash group was exposed to the burning of ashes alone; the Crack group was exposed to the crack and ashes smoke; and the Pf group had their food restricted to match crack food consumption. Data are expressed as mean ± SD. **p < 0.01 and ***p < 0.001; Tukey’s multiple comparison test. SD: standard deviation.
Females exposed to ashes showed a daily weight gain curve similar to that of their food intake (data not shown); however, the total body weight gain of these females was equivalent to the total weight gain of the females of the other groups (Figure 4(b)).
Exposure to crack smoke promoted a decrease in the weight of the nongravid utero when compared to those from the females of the Ash and Pf groups (p < 0.05); however, after calculating the critical value of multiple analyses of inflated type I error rate, the statistical significance no longer exists. No dead fetuses were observed among the groups. Moreover, when fetus’s evaluation and placenta weight were assessed, no statistical differences among the groups were observed (Table 1).
Reproductive performance, maternal measurements, and fetal evaluation of female rats exposed or not to the smoke of 250 mg of crack cocaine, twice a day, from 7 days before mating until the 21st gestational day.a
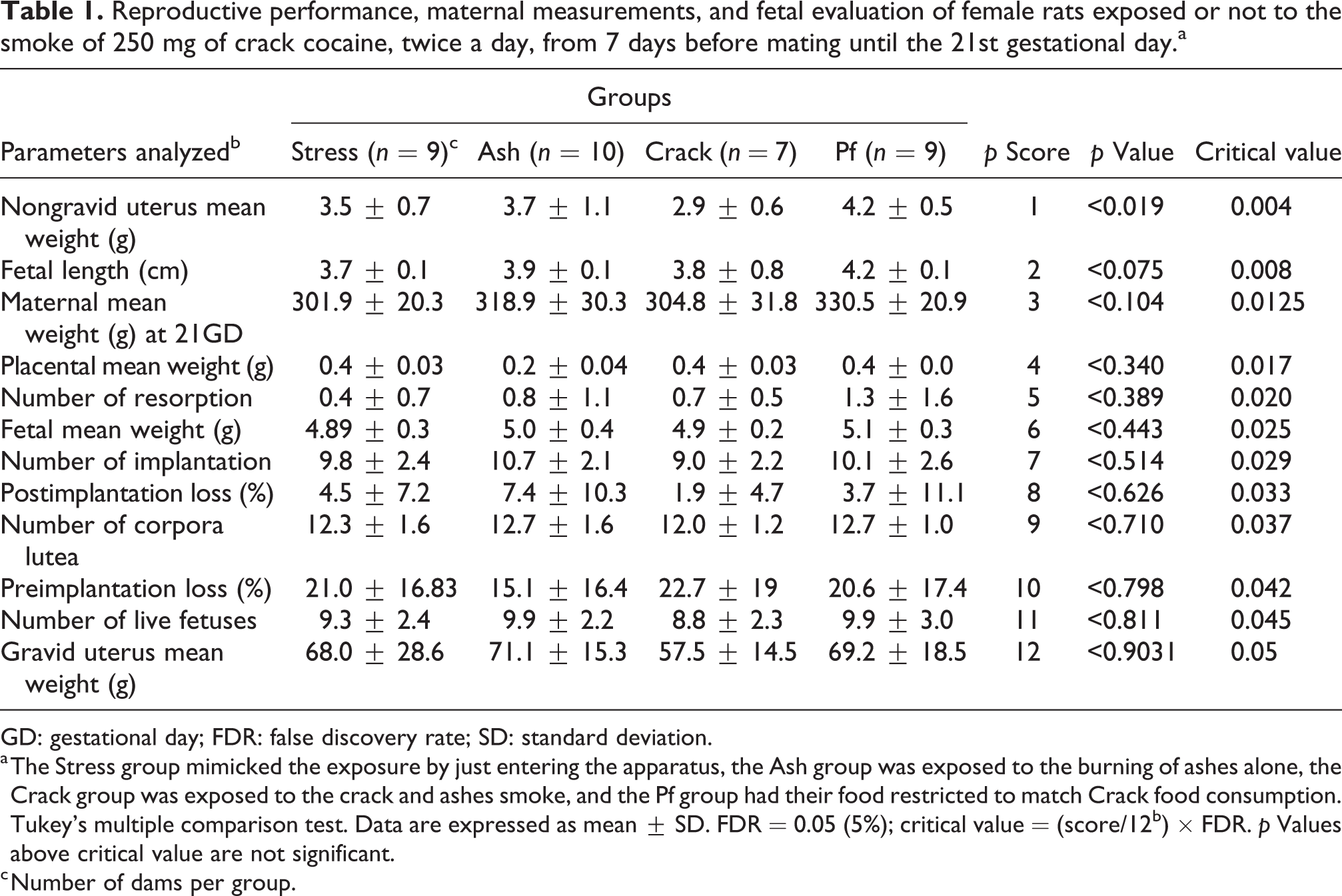
GD: gestational day; FDR: false discovery rate; SD: standard deviation.
a The Stress group mimicked the exposure by just entering the apparatus, the Ash group was exposed to the burning of ashes alone, the Crack group was exposed to the crack and ashes smoke, and the Pf group had their food restricted to match Crack food consumption. Tukey’s multiple comparison test. Data are expressed as mean ± SD. FDR = 0.05 (5%); critical value = (score/12b) × FDR. p Values above critical value are not significant.
c Number of dams per group.
Although most morphological or numerical fetal bone parameters were not altered (Table 2), there was a delayed ossification of the skull in 16% of fetuses exposed in utero to crack smoke (Figure 5).

(a) Photomicrography of fetal skull presenting normal closure of the fontanelle (arrow) and (b) photomicrography of fetus skull from dam exposed to crack smoke during gestation. The arrowhead shows the nonclosure of the fontanelle (magnification ×8).
Incidence of skeletal alterations in litters of female rats exposed or not to the smoke of 250 mg of crack cocaine, twice a day, from 7 days before mating until the 21st gestational day.a

a The table shows the total number of fetuses and litters that were evaluated and the ratio of fetuses and litters affected.
b Only fetuses whose inner bones acquired white color were evaluated. The Stress group mimicked the exposure by just entering the apparatus, the Ash group was exposed to the burning of ashes alone, the Crack group was exposed to the crack and ashes smoke, and the Pf group had their food restricted to match crack food consumption. Different letters denote statistical significance (i.e. any groups with the same footnoted “c” or “d” are not statistically different) (p < 0.05); Fischer’s test.
The comparative analyses of the fetal metameric regions, as proposed by Wilson, 11 did not reveal any limb defects or changes in head and neck, as cleft palate, and hydrocephalus. No changes in the thorax, as cardiac anomalies, were observed. Into the abdominal cavity, no hydronephrosis, renal hypoplasia, or any other alteration in reproductive organs were noticed. On the other hand, the morphometric analysis of the skull revealed a statistically significant decrease in the proportion of the BM in relation to the SC of fetuses exposed to crack in utero when compared to the fetuses of the other groups. Also, a statistically significant decrease of this parameter was observed in fetuses of dams under food restriction (Pf group) when compared to fetuses of females that only entered the box (Stress group) (Figure 6).

BM/SC ratio (%) of fetuses from female rats exposed or not to the smoke of 250 mg of crack cocaine, twice a day, from 7 days before mating until the 21st gestational day. The Stress group mimicked the exposure by just entering the apparatus; the Ash group was exposed to the burning of ashes alone; the Crack group was exposed to the crack and ashes smoke; and the Pf group had their food restricted to match crack food consumption. Data are expressed as mean ± SD. **p < 0.01 and ****p < 0.0001; Tukey’s multiple comparison test. BM: brain mass; SC: skull cap; SD: standard deviation.
Lipid peroxidation, assessed as TBA reactant substances (TBARS), was enhanced in placentas from the Crack and Pf groups but only when compared to the Ash group. Nevertheless, this could be indicative of crack cocaine-induced placental oxidative stress, since the activities of antioxidant enzymes catalase and GPx were increased in placentas from the Crack group compared to the Stress group. Crack exposure resulted in unaltered placental GSH levels, total or oxidized. However, SOD activity was increased solely in the Pf group, accompanied by a decrease in GSSG levels, when compared to the Stress group. The burning of ashes alone also elicited an increase in placental catalase activity (Table 3).
Placental oxidative stress parameters.a
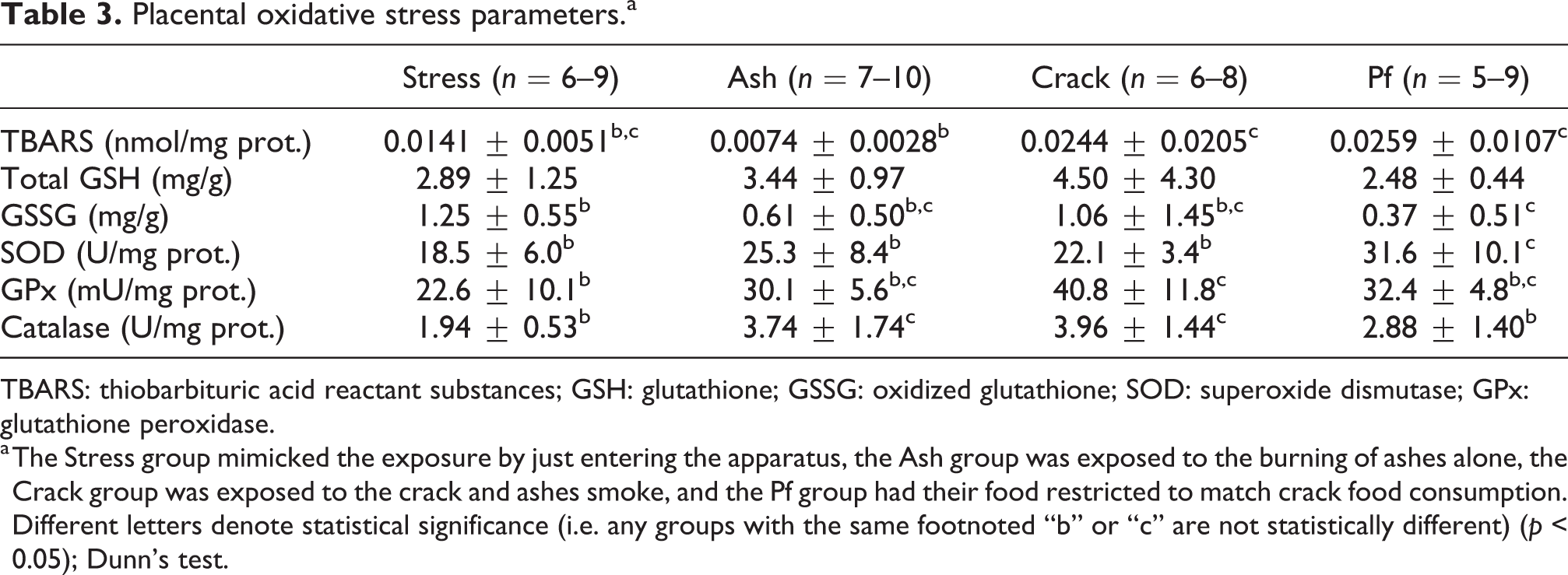
TBARS: thiobarbituric acid reactant substances; GSH: glutathione; GSSG: oxidized glutathione; SOD: superoxide dismutase; GPx: glutathione peroxidase.
a The Stress group mimicked the exposure by just entering the apparatus, the Ash group was exposed to the burning of ashes alone, the Crack group was exposed to the crack and ashes smoke, and the Pf group had their food restricted to match crack food consumption. Different letters denote statistical significance (i.e. any groups with the same footnoted “b” or “c” are not statistically different) (p < 0.05); Dunn’s test.
Discussion
In the present study, we initiated the exposure of female rats to the drug around 7 days before they were pregnant for two main reasons: first, to partially mimic human behavior, since women do not begin to consume crack at conception day, and second, to observe any interferences of a previous exposure in the fertilization or in the embryo implantation, which could result in a smaller number of pups per litter. Indeed, a satisfactory number of pregnant rats was more difficult to achieve in the Crack group (pregnancy rate of 58.3% × 75–83.3% from not exposed groups). A reproductive study performed by Raap et al., 21 with female rats exposed to cocaine hydrochloride, showed changes in the estrous cycle, reducing the transition from the proestrus to the estrus period. In our study, we observed higher frequency of estrus in females exposed to crack than in other groups (data not shown); in addition, even during this favorable period, spermatozoa were not frequently observed in the vaginal smear. This may be related to the sexual dysfunctions promoted by the chronic use of cocaine, such as hyperprolactinemia, which leads to lower sexual interest in females. 22 –24 Thus, it is possible to conclude that crack exposure interferes indirectly with the fertilization rate of rats without affecting embryo implantation, since no differences were observed in the number of pups per litter.
Among the main effects promoted by cocaine, whether in the form of hydrochloride or in crack form, are eating disorders and anorexic effect. 25,26 For this reason, a group of females that received the same amount of food ingested by dams from the Crack group, known as pair-fed (Pf group), was included among the experimental groups. This should clarify whether effects on maternal–fetal evaluations are due to exposure to crack or to lower food intake, since it is well known that malnutrition or reduced nutrient intake during pregnancy can lead to maternal and fetal development alterations, with consequent teratogenic action. 27
Since the evaluation of feed intake was not performed in the period before gestation due to the presence of the male in the cage for mating, it is not possible to assume that there were changes in the food consumption in that period. It is suggested that cocaine leads to an increase in brain dopamine levels in the synaptic clefts of the dorsal striatal region, resulting in undereating. 28,29 Conversely, repeated exposure to psychostimulants may cause neuronal adaptation with the occurrence of tolerance to the anorexic effects. 30 In fact, in the initial period of gestation, the consumption of food by all groups of animals was similar. Later on the gestational period, rats exposed to the smoke of burning ashes had a higher feed intake than rats from the other groups; however, this increase in feed consumption was not accompanied by greater total weight gain when compared to the other groups of animals. As the presence of gnawed ration was not observed in the cages, nor a smaller number of pups per female resulted from this effect, more studies must be conducted to better understand this result.
Previous studies in humans relate the use of cocaine during pregnancy to several complications for both, mother and fetus, such as preterm birth, placental abruption, preeclampsia, spontaneous abortion, lower intrauterine growth, low birth weight, and fetal neurological changes with consequent cognitive and neurodevelopmental outcomes. 2,31 –33 However, similarly to other studies performed with pregnant rats and mice exposed to cocaine hydrochloride, 34 –36 we did not detect abortion or signs of fetal loss. This difference related to reproduction between human and rodents may not be directly related to the physiologic susceptibility of each species, but to the fact that women who use cocaine usually associate it with other drugs, such as alcohol and tobacco, which are well known to produce heavy toxic effects in pregnancy. 37 –39
Even though the use of FDR-controlling procedure discarded a possible statistical significance of the decrease in nongravid utero weight in female rats exposed to crack, little is known about the toxic effect of cocaine on the utero. Some studies have shown that it may inhibit or decrease the expression of beta-adrenergic receptors in the utero, promoting greater contractility during pregnancy, contributing to preterm birth and abortion. 40,41 However, as previously mentioned, no occurrence of preterm birth or miscarriage was observed in the rats. Moreover, this fact alone did not compromise the reproductive outcome of these rats, since no resorptions or alterations in the number and weight of fetuses were observed.
Previous studies in rats and mice have shown that in utero exposure to high doses of cocaine hydrochloride may cause skeletal anomalies and delay the fetal ossification process. 42 –44 In the present study, only the incomplete closure of the fontanel was observed, evidencing that cocaine in the form of crack promotes similar results to the hydrochloride form. This effect may be directly related to the action of cocaine in the process of bone development and mineral deposition and not to maternal undernutrition, since there was no decrease in food intake of these animals. This phenomenon may be related to two mechanisms that are interrelated: decreased levels of osteocalcin, a hormone that regulates bone remodeling, as observed in pregnant users by Rico et al., 45 and lower deposition of calcium in the bones of rodents exposed to cocaine, as observed by Church et al. 46
In rodents, only high doses of cocaine hydrochloride, often close to LD50, or a combination with other drugs, such as ketamine, diazepam, and others, have promoted fetal organogenesis and development abnormalities, such as cranial and abdominal hemorrhagic alterations, palatine cleft, and occurrence of hydronephrosis and cryptorchidism. 36,42 The visceral analysis here performed as proposed by Wilson 11 did not reveal the occurrence of such effects, which may be related to the frequency (twice a day) of exposure to crack smoke, since the dose employed in this study promoted blood levels of cocaine similar to those observed in users.
Although the cranial metamers of the fetuses of rats exposed to crack did not present morphological alterations or presence of edema, hemorrhage, or hydrocephalus, the brains appeared to be smaller than those of fetuses not exposed to the drug. Indeed, later morphometric evaluation of formaldehyde preserved metamer sections, using Nis-Elements (Br)® software detected significant changes in the brain size of these fetuses. This result in brain size corroborates studies with children exposed in utero to the drug when reduced brain volume was analyzed by magnetic resonance imaging (MRI). 47 Changes in cranial parietal distance have been seen in mice 42 ; also a decrease in cranial circumference 48 –50 and occurrence of cloverleaf skull 51 have been shown in neonates of women who used cocaine during pregnancy. To our knowledge, this is the first report about this type of alteration on prenatal exposure of rodents to crack cocaine.
Placental cells can form reactive oxygen species (ROS) by several mechanisms, such as mitochondrial adenosine triphosphate synthesis, xanthine oxidase (XO), cytochrome P450, and NADPH oxidase activities. 52 Cocaine exerts an important vasoconstrictor effect in the placenta, diminishing blood flow and leading to a hypoxic microenvironment. Circulating catecholamines and low O2 pressure may alter placental mitochondrial function, hindering energy production. 53 In human preeclampsia, another condition where placental blood flow is diminished, mitochondrial number and activity increase to compensate the energy requirements of the placenta, contributing to ROS formation and consequent oxidative stress. 54 Moreover, in response to lower O2 pressure, xanthine dehydrogenase/oxidase may shift to increase ROS production, as XO increases in rat placenta between GD16 and GD22. 55 Cytochrome P450 (CYP) enzymes that participate in oxidative inactivation and detoxification of several compounds, including cocaine, are also found in placental mitochondria and microsomes of trophoblastic cells and can yield O2·−. The pattern of CYP isoforms present in the placenta changes during pregnancy, but drug abuse during human pregnancy enhances the presence of specific isoforms later in pregnancy. 56,57 Although placental oxidative stress was detected in GD21 in the present work, further studies must be conducted to evaluate oxidative parameters throughout the whole period of fetal development.
Enhanced TBARS formation in placentas from the Crack group at the end of the gestational period, even in the presence of increased GPx and catalase activities, could be indicative of crack-induced placental oxidative stress. Unexpectedly, the Pf group showed TBARS enhancement in the same manner as seen for the Crack group, although the antioxidant enzyme activation pattern was different as well as the observed decreased GSSG content. Food restriction in this group did not manage to impact animal weight gain and total food consumption was not significantly different from that of the control group, which means that the Pf group was not submitted to any significant caloric restriction. However, the animals may have been submitted to a certain level of stress from not having food available at certain times of the day, which did not happen in the other groups. This anxiety stress may render a different central nervous response than that provoked by crack cocaine intoxication, and it may have promoted different consequences in the oxidative balance in the placenta of these animals. Behavioral and further oxidative stress studies would be necessary to better understand these phenomena.
Conclusions
In conclusion, crack cocaine exposition can promote interferences in the reproductive parameters manifested by decrease in pregnancy rate. Repeated exposure to crack cocaine, twice a day, during pregnancy did not promote anorexic effects to the dams that could promote the delayed fetal ossification process here observed, showing a direct effect of this drug on fetuses. Similarly, it is possible to suggest that crack cocaine has an oxidative potential on the placenta, which cannot be related to changes in the food intake of the animals. Finally, although classical methods of fetal visceral analysis proposed by Wilson 11 did not show alterations in the development of fetuses exposed to crack, the morphometric analysis here employed proved to be valuable to show effects observed by other more sophisticated methods, such as MRI, 47 clearly demonstrating the deleterious effect of cocaine on the development of rat BM. Our study is, to our knowledge, the first in the literature that shows that daily exposure to crack, even in lower frequency than that performed by users (women consumption can reach more than 20 rocks per day), 58 has a teratogenic potential.
Footnotes
Declaration of conflicting interests
The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was supported by FAPESP (Fundação de Amparo à Pesquisa do Estado de São Paulo), process number 2016/17050-9, Brazil.