
Abstract
The present study has been designed to investigate the ameliorative effect of grape seed proanthocyanidins (GSP) on cadmium (Cd)-induced oxidative damage in rat erythrocytes. Twenty four male Wistar rats were divided into four groups: control, GSP-treated group (100 mg kg−1 body weight (BW)), Cd-treated group (cadmium chloride, 5 mg kg−1 BW), and GSP + Cd-treated group in which GSP was orally pre-administered 90 min before Cd intoxication for 4 weeks. At the end of the experimental period, blood samples were collected by cardiac puncture and were processed for various biochemical estimations. The extent of oxidative damage in isolated rat erythrocyte membrane was assessed by measuring lipid peroxidation, enzymatic and non-enzymatic content, calcium ion (Ca2+)/magnesium ion (Mg2+)-ATPase and sodium ion (Na+)/potassium ion (K+)-ATPase activities, free iron, calcium, hydrogen peroxide (H2O2) concentration, and osmotic fragility. Our results unveiled that Cd intoxication significantly increased the erythrocyte lipid peroxidation markers and decreased the activity of enzymatic and non-enzymatic markers in erythrocytes. Conversely, GSP pretreatment significantly prevented the decrease in the activities of antioxidant enzymes and membrane-bound ATPases. GSP also restored the levels of iron, calcium, and H2O2 in Cd-treated rats. Conformational changes in erythrocytes of various groups were also determined using morphological and ultrastructural electron microscopic analysis. The findings of our study clearly revealed that GSP affords superior protection against Cd-induced reactive oxygen species generation, lipid peroxidation, and free radical generation in Cd-treated rats, which presumably reflects the ability of this flavonoid to protect erythrocytes and lymphocytes of rats from the toxic effects of Cd.
Introduction
Cadmium (Cd) is one of the most toxic non-essential heavy metals that causes concern as an environmental and occupational toxicant, endangering human and animal health. Exposure to Cd causes toxic effects on a variety of tissues, but the first to be affected is blood. Following absorption, a greater proportion of the Cd is transported in the blood, where it binds with the erythrocyte membranes and plasma albumin. Cd accumulates in blood and causes a longer half-life with a fast component of 3–4 months and a slow component of approximately 10 years. Therefore, the blood Cd level is considered as the most valid marker and a good reflector of the Cd body burden. In the blood and tissues, Cd stimulates the formation of metallothioneins and reactive oxygen species (ROS), thus causing oxidative stress in red blood cells (RBCs), lymphocytes and in various tissues.
Erythrocytes, the unique carriers of oxygen, are susceptible to the free radical attack induced by Cd due to the presence of both oxygen transport with redox active haemoglobin molecules. Due to the high contents of polyunsaturated fatty acids (phospholipids, glycolipids and sterols) in their membranes, which are potent promoters of ROS, makes the erythrocytes a good substrate for oxidative damages. 1 Cd-bound erythrocyte is released into plasma when the erythrocyte lifetime has expired or following haemolysis, which can be achieved by the activation of neutrophils. Exposure of erythrocytes to ROS leads to lipid peroxidation, changes in cellular morphology, protein cross linking, antioxidant enzymes depletion and conformation changes in membrane proteins. 2 Due to Cd-induced oxidative stress, cell losses the plasticity with impaired deformability, associated with changes in the cytoskeleton network and also the suicidal death of erythrocytes, which leads to the elimination of erythrocytes and finally to anaemia. 3
The presence of antioxidants such as polyphenols, carotenoids and anthocyanins in fruits and vegetables helps to prevent various free radical-mediated diseases. A broad diversity of physical and chemical properties with the presence of hydroxyl-containing phenolic groups in these phytochemicals renders them as excellent antioxidants due to their quenching action of free radicals by electron or proton (hydrogen) donation and can protect body tissues against oxidative stress. 4 Proanthocyanidins are a class of phenolic compound, extracted from grape seeds, which take the form of oligomers or polymers of polyhydroxy flavan-3-ol units, such as (+) – catechin and (−) – epicatechin, and are condensed through the link models of C4–C8 or C4–C6 bonds. 5 The electronic configuration of grape seed proanthocyanidins (GSP) allows the easy release of electrons to free radical species (R.) such as superoxide anion, hydroxyl, peroxy and nitric oxide (NO) radicals. 5
During hydrogen donation, hydroxyl group attached to the aromatic ring structure of GSP enables them to undergo redox reactions scavenging free radicals more easily. Also the aromatic and heterocyclic ring as well as multiple unsaturated bonds helps to delocalize and regulate the free radicals. 5 GSP has a potent inhibitory effect towards H2O2, xanthine oxidase, organic hydroperoxide, dialuric acid, and 2,2′ azobis(2-amidinopropane)dihydrochloride-induced haemolysis. 6 One of the most advantageous features of proanthocyanidins’ free radical scavenging activity is that it is incorporated within cell membranes. The presence of hydrophilic residues within the flavan-3-ol molecule allows GSP to interact with phospholipid head groups and to be adsorbed onto the surface of membranes. 7 Nevertheless, the protective effect of GSP on Cd-induced oxidative stress in erythrocyte and lymphocyte remains to be explored. In view of the above prospective benefits, the present study has been designed to appraise the potential ameliorative action of GSP against Cd-induced toxicity on the erythrocytes and lymphocytes of rats.
Materials and methods
Chemicals
GSP (batch no. 005004) was obtained from Inter Health Nutraceuticals, Inc. (Benicia, California, USA). GSP is a natural extract containing approximately 54% dimeric, 13% trimeric and 7% tetrameric proanthocyanidins. Cadmium chloride and other fine chemicals were obtained from Pfizer (west Mumbai, Maharashtra, India). OSMMR2000 glucose-6-phosphate dehydrogenase (G6PD) kit was obtained from R&D Diagnostics Ltd (Holargos, Greece). The other chemicals used for the present study were procured from Sigma Chemical Co. and Sisco Research Laboratories (Mumbai, Maharashtra, India). The chemical structure of GSP is shown in Figure 1.

Chemical structure of grape seed proanthocyanidins formed by (7) - epicatechin units linked by 4 → 8 bonds (e.g. B-type dimer, C-type trimer).
Animals and experimental design
The experiment was carried out on healthy male albino rats of Wistar, which were reared and acquired from Central Animal House, Department of Experimental Medicine, Rajah Muthiah Medical College, Annamalai University (India). Animals with body weight (BW) ranging from 180 g to 200 g were used in the study and housed in polypropylene cages lined with husk. The rats were kept in a normal day and in night schedule (12-h light/12-h dark cycle). Each rat was given pellet foodstuff from the Amrut Laboratory Animal Feed (Pune, Maharashtra, India) and water ad libitum. The animal treatment and protocol employed were approved by the Institutional Animal Ethics Committee, Annamalai University (vide no. 1020, 2013). After an acclimatization period of 1 week, 24 rats were divided arbitrarily into four groups (6 rats/group) treated orally with intragastric tubes.
Group I: normal rats received vehicles only for 28 days and served as a normal group. Group II: treated with Cd (5 mg kg−1 BW) in normal saline for 28 days.
8
Group III: treated with GSP (100 mg kg−1 BW) in normal saline for 28 days.
7
Group IV: treated with Cd (5 mg kg−1 BW) + GSP (100 mg kg−1 BW) administered 90 min before the administration of Cd.
At the final stage of the experimental period, the rats in each group were allowed to fast overnight and anatomized under anaesthesia by diethyl ether and then killed by decapitation. Blood samples were collected in two heparinized tubes and heparinized vials by cardiac puncture and were processed for biochemical estimation. The blood obtained was centrifuged to separate the plasma and cells. Packed red cells were washed three times with ice-cold phosphate-buffered saline (PBS buffer 0.01 M, pH 7.4, containing 0.15 M sodium chloride (NaCl)) and used for the estimation of thiobarbituric acid reactive substances (TBARS).
Hematological parameters
Level of mean cell volume (MCV), haematocrit (HCT), haemoglobin (HGB), mean cell haemoglobin (MCH), mean cell haemoglobin concentration (MCHC), platelets (PLT), RBC count and white blood cell (WBC) count were measured using a Sysmex hematology analyzer (model K4500; Japan).
Plasma analysis
In the plasma, activities of aspartate transaminase (AST) and alanine transaminase (ALT) were measured using the method of Reitman and Frankel, 9 and acid phosphatase (ACP; U/L), alkaline phosphatase (ALP; U/L) and lactate dehydrogenase (IU/L) were measured by an auto analyser (BM/HIT-ACH) using the kit from Randox Laboratories Ltd (Crumlin, Ireland). Plasma γ-glutamyl transferase (γGT) was estimated by the method of Rosalki and Tarlow, 10 vitamin C by the method of Roe 11 and vitamin E by the method of Desai. 12 Lipid peroxidation in plasma was estimated spectrophotometrically by measuring TBARS and lipid hydroperoxides (LHs) by the method of Niehius and Samuelsson 13 and Jiang et al., 14 respectively. Reduced glutathione (GSH) was determined by the method of Ellman. 15
Preparation of erythrocyte membranes
The erythrocyte membranes were prepared according to the method of Dodge et al. 16 All procedures were carried out at 4°C. Erythrocytes were separated from plasma and buffy coat by centrifugation at 2000g for 10 min, and washed three times with isotonic 10 mM PBS (150 mM NaCl, 10 mM disodium phosphate (Na2HPO4)–monosodium phosphate (NaH2PO4), pH 7.4). The erythrocytes were then haemolyzed with 40 volumes of hypotonic 5P8 buffer (5 mM Na2HPO4–NaH2PO4, 0.1 mM phenylmethanesulphonyl fluoride, pH 8.0) followed by centrifugation at 15,000g for 20 min to collect the membrane fractions. The membrane pellets were washed with 5P8 buffer and centrifuged until the supernatants were colourless (white membranes). For determination of ATPase activity, the membranes were prepared in tris(hydroxymethyl)aminomethane (Tris)–hydrochloric acid (HCl) buffer in place of PBS. Membrane protein concentration was determined by the method of Lowry et al. 17 using bovine serum albumin as a standard.
Estimation of erythrocyte membrane lipid peroxidation
Lipid peroxidation in the erythrocyte membrane was assayed as malondialdehyde (MDA) according to Ohkawa et al. 18 and expressed as nanomoles per milligram protein. Increased levels of ROS during oxidative stress attack the polyunsaturated fatty acids of membranes to release cytotoxic aldehydes like MDA, which serves as a good indicator of the oxidative stress status. Packed red cells (0.2 mL) were used for the estimation of MDA as TBARS. 19 The absorbance of supernatant was measured at 532 nm and corrected for unspecific turbidity by subtracting the value of absorbance at 600 nm. Haemoglobin concentration was measured by the cyanmethemoglobin method of Tentori and Salvat. 20 Protein carbonyl (PC) levels were measured according to the method described by Reznick and Packer 21 based on spectrophotometric (Beckman DU 640 B, Switzerland) detection of the reaction of 2,4-dinitrophenylhydrazine (DNPH) with PC to form protein hydrazones. Briefly, after precipitation of protein with an equal volume of 1% tricarboxylic acid, the pellet was resuspended in 10 mM DNPH in 2 N HCl or with 2 N HCl as control blank. After the washing procedure with 1:1 ethanol/ethyl acetate, the final pellet was dissolved in 6 M guanidine hydrochloride. The carbonyl group was determined from the absorbance at 370 nm. The result was expressed as nanomoles of carbonyl groups per milligram of haemoglobin with molar coefficient of 22,000 M−1cm−1. The absorbance ratio A 233/A 215, defined as the oxidation index, was used as a relative measurement of Cds according to Konings. 22 The role of NO synthase (NOS) was indirectly assessed by estimating the amount of production of NO in both plasma and erythrocytes. NO decomposes rapidly in aerated solution to form stable nitrite/nitrate products. In our study, nitrite accumulation was estimated by Griess reaction 23 and was used as an index of NO production. The amount of nitrite in the sample (micromolar unit) was calculated from a sodium nitrite curve. 24
Antioxidant enzymes
Erythrocytes from the second tube were lysed by fourfold dilution with H2O followed by repeated freezing–thawing cycles. The haemolysates obtained were used for the determination of total GSH contents and antioxidant enzyme activities. Haemoglobin in erythrocytes was determined by the method of Samuel. 25 GSH content was estimated according to the method of Beutler et al. 26 and expressed as micromoles per gram haemoglobin (Hb). Superoxide dismutase (SOD) activity was estimated according to the method described by Misra and Fridovich. 27 Catalase (CAT) activity was determined using the method described by Aebi 28 by measuring H2O2 decomposition at 240 nm. Glutathione peroxidase (GPx) activity was assayed using the method described by Flohe and Gunzler 29 by the subsequent oxidation of reduced nicotinamide adenine dinucleotide phosphate (NADPH) at 240 nm with t-butyl-hydroperoxide as substrate. The values of SOD, CAT and GPx are expressed in units per gram of haemoglobin (U g−1 Hb). Glutathione reductase (GR) activity in erythrocytes was assayed by the methods of Goldberg and Spooner. 30 The GR activity in erythrocytes has been expressed as nanomoles NADPH oxidized to nicotinamide adenine dinucleotide phosphate (NADP) per gram of Hb per minute. Glutathione-S-transferase (GST; E.C.2.5.1.18.) was also assayed by haemolysates. 31 The G6PD kit (Span Diagnostics Ltd; Chattisgarh, Kolkata, India) was used for a rapid quantitative measurement of G6PD activity coupled with a simultaneous evaluation of the haemoglobin content in the same sample, and the results are expressed in units per gram of haemoglobin.
Assay of membrane-bound enzymes
Na+/K+-ATPase was assessed in erythrocyte membrane preparation according to the method of Quigley and Gotterer. 32 Sodium ion (Na+)/potassium ion (K+)-ATPase activity was measured under two conditions: in the presence of magnesium ion (Mg2+), Na+/K+ (total ATPase) and in the presence of Mg2+, Na+/K+ and ouabain. The Na+/K+-ATPase activity was measured as the difference between total ATPase activities and ouabain-insensitive ATPase activities. The inorganic phosphate released by the action of ATPases was estimated by the method of Fiske and Subbarrow. 33 The calcium ion (Ca2+)-ATPase activity was measured according to the method of Desaiah et al. 34 The Mg2+-ATPase activity was determined in the presence of 1 mM ethylene glycol tetraacetic acid (which specifically chelates Ca2+ ion), and this was subtracted from the total activity in order to obtain the net Ca2+-ATPase activity. The erythrocyte α-tocopherol level 35 and ascorbic acid level 36 were also analysed.
Free iron, H2O2 and calcium measurement
Erythrocytes free iron was determined according to Leardi et al. 37 using a commercially available kit from Biomaghreb (Tunisia). At acidic pH 4.8, all ferric ions (Fe3+) released from transferrine were reduced by ascorbic acid into ferrous ions (Fe2+), which constitutes with ferrozine a purple colourful complex measured at 560 nm. Briefly, 50 μL of erythrocytes extract was added to 250 μL of reaction mixture containing ascorbic acid (5 g L−1) and ferrozin (40 mM), and incubation was performed at 37°C for 10 min. The erythrocyte H2O2 level was determined enzymatically according to Kakinuma et al. 38 using a commercially available kit from Biomaghreb. Erythrocyte ionisable calcium was determined according to Stern and Lewis 39 using a commercially available kit from Biomaghreb. At basic pH, calcium constitutes with cresolphatalein a purple, colourful complex measurable at 570 nm. Briefly, 50 μL of erythrocyte extract was added to 650 μL of reaction mixture containing 2-amino 2-methyl 1-propanol buffer (500 mmol L−1), cresolphatalein (0.62 mmol L−1) and hydroxy-8 quinoleine (69 mmol L−1). Incubation was carried out at room temperature for 5 min assuming the complex was stable for 1 h.
Erythrocyte osmotic fragility test
Erythrocyte osmotic fragility was estimated according to the methods of Azeez et al. 40 and Mineo et al. 41 Briefly, 0.02 mL of blood of each rat was transferred into microtubes containing 1 mL of increasing concentration of phosphate-buffered NaCl solution (0.0, 0.1, 0.5 and 0.9%) at pH 7.4. The microtubes were then gently mixed, incubated at room temperature for 30 min and then centrifuged at 2000 r min−1 for 15 min. The supernatant (200 μL) of each microtube was decanted into 96-well micro-plate, and its optical density was determined spectrophotometrically at 540 nm (micro-plate reader, Bio-Rad Laboratories, Hercules, California, USA). The percentage of RBC haemolysis in each NaCl concentration was determined using haemolysis in distilled water (0.0% NaCl) as the maximum percentage. The higher haemolysis occurs, the greater is osmotic fragility of RBC.
Lymphocyte separation
Five milliliters of the whole blood samples obtained by cardiac puncture were diluted 1:1 with PBS and carefully layered on the top of a lymphocyte separation medium having a density of 1.077 g mL−1 (aqueous solution of Ficoll, 57 g L−1) in a centrifugation tube and in a ratio of 1:1. After centrifugation for 20 min at 1000g, gradient-separated lymphocytes were recovered, diluted 1:1 with PBS and centrifuged for second time at 1000g for 5 min. The cell pellets were resuspended in 0.5 mL of PBS, and the cell concentration was adjusted to 5000 cells μl−1 in preparation for the comet assay.
Comet assay
The comet assay was performed under alkaline conditions according to the method described by Singh et al.
42
with slight modifications. A 10 μL aliquot of a cell suspension was mixed with 90 μL of preheated 0.5% (w/v) low-melting point agarose (LMP) in PBS at 37°C, and the mixtures were then added to frosted glass microscopic slides precoated with a 1.5% normal melting point agarose. After solidification of the second layer, a third layer of 100 μL of LMP agarose (0.5% w/v) was applied. Immediately after the addition of the cells and their inclusion into the agarose and the solidification of the gel, the slides were then washed with PBS, immersed in a cold lysing solution (2.5 M NaCl, 100 mM Na2 EDTA, 10 mM Tris, 1% Triton X-100 and 10% dimethyl sulphoxide, pH 10.0) and incubated overnight at 4°C. At the end of the incubation period, the slides were washed with PBS and placed in an electrophoresis tank. DNA was allowed to unwind for 30 min in freshly prepared alkaline electrophoresis buffer (1 mM Na 2 EDTA and 0.3 N sodium hydroxide, pH 13.0). Electrophoresis was conducted at 300 mA at 4°C for 30 min. All procedural steps were performed under yellow light conditions to minimize additional DNA damage. The slides were then neutralized by washing three times with Tris–HCl buffer (0.4 M, pH 7.5) and stained with ethidium bromide (20 μg mL−1). The slides were studied using a fluorescent microscope (Nikon 100, Tokyo, Japan) attached to a charge-coupled device camera connected to a personal computer. Fifty individual cells were selected for calculations for each analysis, and four separate experiments (four slides for each experimental point) were conducted for each series. Single cells were analysed with Casp 1.2.2 software. The DNA damage was expressed as tail length, olive tail moment and % tail DNA:

A higher % tail DNA indicated a higher level of DNA damage.
Morphological evaluation
Blood samples were collected using heparinized tubes. Immediately after mixing with blood by inverting the tubes, blood smears were made by spreading one drop on a slide and then staining with the Giemsa solution. RBC morphology was determined using a Nikon Optiphot-2 microscope with an oil immersion 100/1.25 objective.
Electronic structural morphology
Fresh blood samples were drawn from treated and control rats, and a drop of blood was immediately immersion fixed in 2.5% glutaraldehyde made in 0.1 M phosphate buffer (pH 7.4). After 1 h of fixation, cells were centrifuged at 1000–1500 r min−1 and the pellets were resuspended in triple distilled water. After two to three washings, the final pellet was suspended in triple distilled water. A drop of the sample was smeared on the metallic scanning electron microscopic stubs, which were loaded with a conductive silver tape on the top. The stubs were then coated with gold to a thickness of 100 Å using a sputter-ion coater, with a gold source, for 4–5 min and the specimens were finally observed under scanning electron microscope, JSM-6100 (Redwood City, California, USA).
Statistical analysis
All the data were expressed as mean ± SD of a number of experiments (n = 6). The statistical significance was evaluated by one-way analysis of variance (ANOVA) using Statistical Package for the Social Sciences version 15.0 (SPSS, Cary, North Carolina, USA) and the individual comparisons were obtained by Duncan’s multiple range test. Statistical significance was determined by one way ANOVA followed by post hoc test. Values were considered as statistically significant when p < 0.05.
Results
Effect of GSP on morphological changes
Table 1 depicts the effect of Cd and GSP on BW gain, water intake and food intake in normal and experimental rats. In Cd-treated rats, a decreased BW gain with reduced water and pellet consumption was noted (p < 0.05). Interestingly, a significant improvement for all these changes was found on pre-administration of GSP in a dose-related manner. GSP alone did not show any significant difference when compared with the control rats.
Body weight, absolute and relative kidney weight, food and water intake in control and experimental rats.a

DMRT: Duncan’s multiple range test.
aValues are given as mean ± SD from six rats in each group. Values with different superscript letters (b to d) in the same column differ significantly at p < 0.05 (DMRT).
Haematological parameters
Table 2 summarizes the levels of WBC, RBC, PLT, MCV and HCT and the concentration of HGB, MCH and MCHC in blood of normal and experimental rats. In our study, a significantly (p < 0.05) decreasing levels of MCV, HCT, WBC, RBC and PLT and decreasing concentration of HGB, MCH and MCHC were found in Cd-exposed rats when compared with the normal rats. Pretreatment of GSP effectively attenuated the Cd-induced alterations in haematological variables. Except for a significant increase in RBC count in the administration of GSP alone treated rats and did not show any alterations in other clinical haematological variables and didn’t differs significantly from that of the normal rats.
Effect of GSP on haematological parameter studies in the blood of control and experimental rats.a
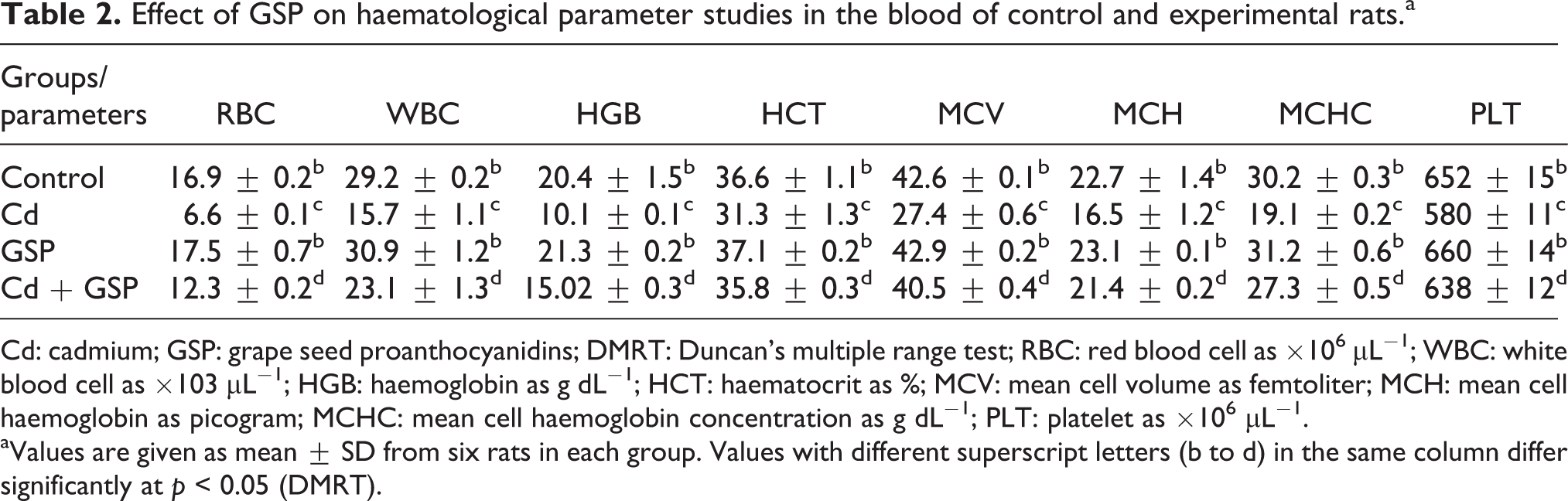
Cd: cadmium; GSP: grape seed proanthocyanidins; DMRT: Duncan’s multiple range test; RBC: red blood cell as ×106 μL−1; WBC: white blood cell as ×103 μL−1; HGB: haemoglobin as g dL−1; HCT: haematocrit as %; MCV: mean cell volume as femtoliter; MCH: mean cell haemoglobin as picogram; MCHC: mean cell haemoglobin concentration as g dL−1; PLT: platelet as ×106 μL−1.
aValues are given as mean ± SD from six rats in each group. Values with different superscript letters (b to d) in the same column differ significantly at p < 0.05 (DMRT).
Effects of GSP on plasma biomarker enzymes
A significant (p < 0.05) increase in the activities of the plasma biomarker enzymes, viz, AST, ALT, ACP, LDH and γGT with decreased activity of ALP in the Cd-treated group when compared with the normal group is depicted in Figure 2. Pre-administration of GSP in Cd -intoxicated rats for 4 weeks significantly (p < 0.05) reduced the levels of serum AST, ALT, ACP, LDH and γGT activities and stabilized ALP activity. There is no significant difference between GSP alone treated rats and control rats.

Effects of GSP against Cd on plasma biomarker enzymes in control and experimental rats. Values are expressed as mean ± SD for six rats in each group. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance; Cd: cadmium.
The data presented in Figure 3 show the changes in plasma lipid peroxidation indices TBARS (A), LH (B), NO production (C) and non-enzymatic antioxidant levels in control and experimental rats. In rats treated with Cd, the levels of plasma TBARS, LH and NO production were significantly (p < 0.05) increased when compared with the control rats, whereas the levels of non-enzymatic antioxidants, namely, vitamins C (D), vitamin E (E) and GSH (F), were significantly (p < 0.05) decreased upon Cd intoxication. Administration of GSP in Cd-intoxicated rats significantly increased the depleted levels of non-enzymatic antioxidants with a significant decrease in Cd-induced lipid peroxidation as compared to the Cd alone treated rats. Rats administered GSP alone did not show significant difference when compared with the control rats.

Effects of GSP against Cd on plasma oxidative stress markers and nonenzymatic antioxidants in control and experimental rats. Values are expressed as mean ± SD for six rats in each group. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance; Cd: cadmium.
Effect of GSP on erythrocyte membrane oxidative stress markers and non-enzymatic antioxidant status
Data presented in Table 3 show a significant (p < 0.05) increase in the level of MDA concentration in erythrocyte membrane of Cd treated group when compared with the control group. The level of PC and NO production was also significantly (p < 0.05) elevated in rat erythrocyte membrane treated with Cd when compared with the control group. GSP along with Cd effectively ameliorated the toxic effect of Cd significantly by reducing the levels of MDA, PC content and NO production in the erythrocyte membrane as compared to the Cd alone treated group. Figure 4 shows the levels of non-enzymatic antioxidants, namely, vitamins C and E and GSH, in control and experimental rats. The levels of vitamins C and E (A) and GSH (B) were significantly (p < 0.05) decreased in the rat erythrocytes treated with Cd when compared with the control group. The depleted levels of vitamins C and E (A) and GSH (B) were significantly (p < 0.05) restored with GSP pre-administration in Cd-intoxicated rats. There is no significant difference between rats treated with GSP alone and control rats.
Effect of GSP on erythrocyte membrane oxidative stress markers in Cd-treated rats.a

Cd: cadmium; PC: protein carbonyl; GSP: grape seed proanthocyanidins; DMRT: Duncan’s multiple range test; NO: nitric oxide; MDA: malondialdehyde; Hb: haemoglobin.
aValues are given as mean ± SD from six rats in each group. Values with different superscript letters (b to d) in the same column differ significantly at p < 0.05 (DMRT).

Effects of GSP on Cd-induced changes in non-enzymatic antioxidant activity of erythrocytes. Values are expressed as mean ± SD for six rats in each group. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance; Cd: cadmium.
Effect of GSP on Cd-induced changes in antioxidant enzyme activity of erythrocytes
Table 4 shows that treatment with Cd caused a significant (p < 0.05) decrease in the activities of SOD, CAT, GPx, GR, GST and G6PD when compared with the control. A significant (p < 0.05) recovery relating to the activities of SOD, CAT, GPx, GR, GST and G6PD was observed in response to the pre-administration of GSP with Cd.
Effect of GSP on Cd-induced changes in antioxidant enzyme activity of erythrocytes.a
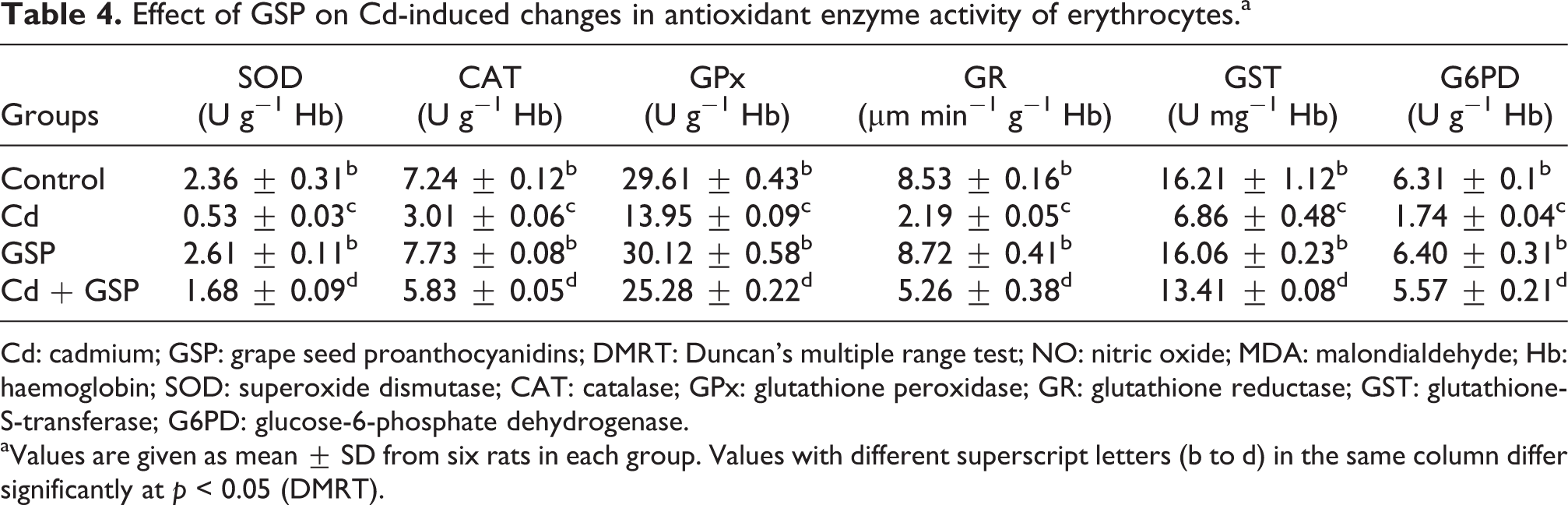
Cd: cadmium; GSP: grape seed proanthocyanidins; DMRT: Duncan’s multiple range test; NO: nitric oxide; MDA: malondialdehyde; Hb: haemoglobin; SOD: superoxide dismutase; CAT: catalase; GPx: glutathione peroxidase; GR: glutathione reductase; GST: glutathione-S-transferase; G6PD: glucose-6-phosphate dehydrogenase.
aValues are given as mean ± SD from six rats in each group. Values with different superscript letters (b to d) in the same column differ significantly at p < 0.05 (DMRT).
Effect of GSP on Cd-induced alterations in erythrocyte membrane-bound ATPases
Figure 5 shows the effect of GSP on Cd intoxicated rat erythrocyte membrane-bound Na+/K+-ATPase, Mg2+-ATPase and Ca2+-ATPases levels in control and experimental rats. The levels of erythrocyte membrane-bound ATPases were found to be significantly (p < 0.05) decreased when compared with that of the normal group. Pre-administration of GSP along with Cd had significantly (p < 0.05) improved the levels of erythrocyte membrane-bound ATPases when compared with the Cd-treated group. Administration of GSP alone to rats did not show any significant alterations in erythrocyte membrane-bound ATPases when compared with the normal rats.

Effect of GSP on RBC membrane-bound ATPases (Na+/K+-ATPase, Mg2+-ATPase and Ca2+-ATPases) of control and experimental rats. Values are expressed as mean ± SD for groups of six rats in each. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance; Na+: sodium ion; K+: potassium ion; Ca2+: calcium ion; RBC: red blood cell.
Erythrocyte iron, H2O2 and calcium levels
Figure 6 depicts the effect of GSP on erythrocyte iron, H2O2 and Ca2+ in treated rats. The levels of erythrocyte iron, H2O2 and Ca2+ were significantly (p < 0.05) decreased in Cd-treated rats when compared with the control group. Pre-administration of GSP along with the Cd had counteracted the effect of Cd and significantly (p < 0.05) improved the levels of free iron, calcium and H2O2 in erythrocytes of rats as compared to the Cd alone treated rats.

Effects of GSP on Cd induced changes in free iron, calcium and H2O2 in erythrocytes of control and experimental rats. Values are expressed as mean ± SD for six rats in each group. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance; H2O2: hydrogen peroxide; Cd: cadmium.
Effects on erythrocyte osmotic fragility
Changes in RBCs haemolytic pattern after they’re exposed to different concentrations of NaCl solution were measured to determine the erythrocyte osmotic fragility of all treated groups. As shown in Figure 7, the osmotic fragility of RBCs of Cd group was significantly higher than that of the Cd-pretreated GSP group and normal groups at each tested NaCl solution.

Effect of GSP on erythrocyte osmotic fragility in control and experimental rats. Values are expressed as mean ± SD for groups of six rats in each. Statistical significance was determined by one-way ANOVA followed by post hoc test. Values are given as mean ± SD for six rats in each group. Values not sharing a common superscript letter (a to c) differ significantly at p < 0.05 (DMRT). DMRT: Duncan’s multiple range test; GSP: grape seed proanthocyanidins; ANOVA: analysis of variance.
Effect of GSP on DNA damage of rat blood lymphocytes treated with Cd
The comet assay is a rapid, sensitive and versatile method for the quantification of DNA damage in individual cells. A significant (p < 0.05) increase in tail length, % tail DNA and olive tail moment were observed with Cd-treated rats when compared with the control rats, as shown in Figure 8(b). GSP had a significant (p < 0.05) inhibitory effect on Cd-induced DNA damage. No significant damages were observed between control and rats treated with GSP alone. Figure 8(a) shows the representative photomicrographs of comets stained with ethidium bromide at 200× magnification showing the DNA migration pattern in lymphocytes, where the control rats show no DNA migration. Whilst extensive DNA migration was observed in Cd-treated rats when compared with the control rats. GSP pre-administered rats with Cd intoxication shows the minimal DNA migration when compared with the Cd alone treated rats. GSP alone administered rats show no DNA migration.

(
Morphological evaluation
Figure 9 shows the effect of Cd and GSP on histology of erythrocytes. Normal architecture, round, biconcave discs having a central pallor was observed in the control group, as shown in Figure 9(a). In the Cd-treated group (Figure 9(c)), there were remarkable membrane destruction and haemolytic changes in erythrocytes, few RBC were irregularly spaced projections, schistocytes (star), stomatocytes (S) and echinocytes (arrow head) were noticed. Several RBCs possessing no area of central pallor spherocytes were noticed (arrow). In the GSP-pretreated Cd group, as shown in Figure 9(d), these changes exhibited a gradual recuperation and were less seen than in the Cd-treated group. The rat received GSP alone showed a normal picture similar to that of the control Figure 9(b).

Effect of GSP on erythrocyte histopathology in control and experimental rats. (a) shows normal RBCs with central pallor. (b) GSP-treated group showed normal RBC as seen in the control group. (c) Cd-treated group showed the presence of acanthocytes, spherocytes (arrow), echinocytes (arrow head), stomatocytes (S) and schistocytes (star), fragmented and faded RBC were seen in the majority. GSP pretreated rats (d) showed the recovered architecture of RBC as compared to Cd-treated group, but some faded cells with no resurgence were also seen. ×1100. Bars = 5 μm. GSP: grape seed proanthocyanidins; RBC: red blood cell; Cd: cadmium.
Protective effect on RBC structural morphology
On histological examination, RBC in the control group (Figure 10(a)), displayed a specific biconcave disk shape. The administration of Cd (Figure 10(c)) showed a marked increase in echinocyte (E), discocyte (D), spherocytes (S), schizocytes (SZ) and acanthocytes (A). Many RBCs with thin and long extensions of their membranes were found, so-called myelinic figures, whilst the discocyte (D), acanthocytes (A) and the RBCs with other aberrant shapes were the most frequent abnormal RBCs observed in Figure 10(d). In Figure 10(e), GSP-pretreated Cd group displayed a normal, recovered biconcave disk shape, but little acanthocytes (A) were also observed, whilst the most frequent abnormal RBCs were slightly crenated by GSP. The rat received GSP alone (Figure 10(b)) showed a normal morphological picture similar to that of the control.
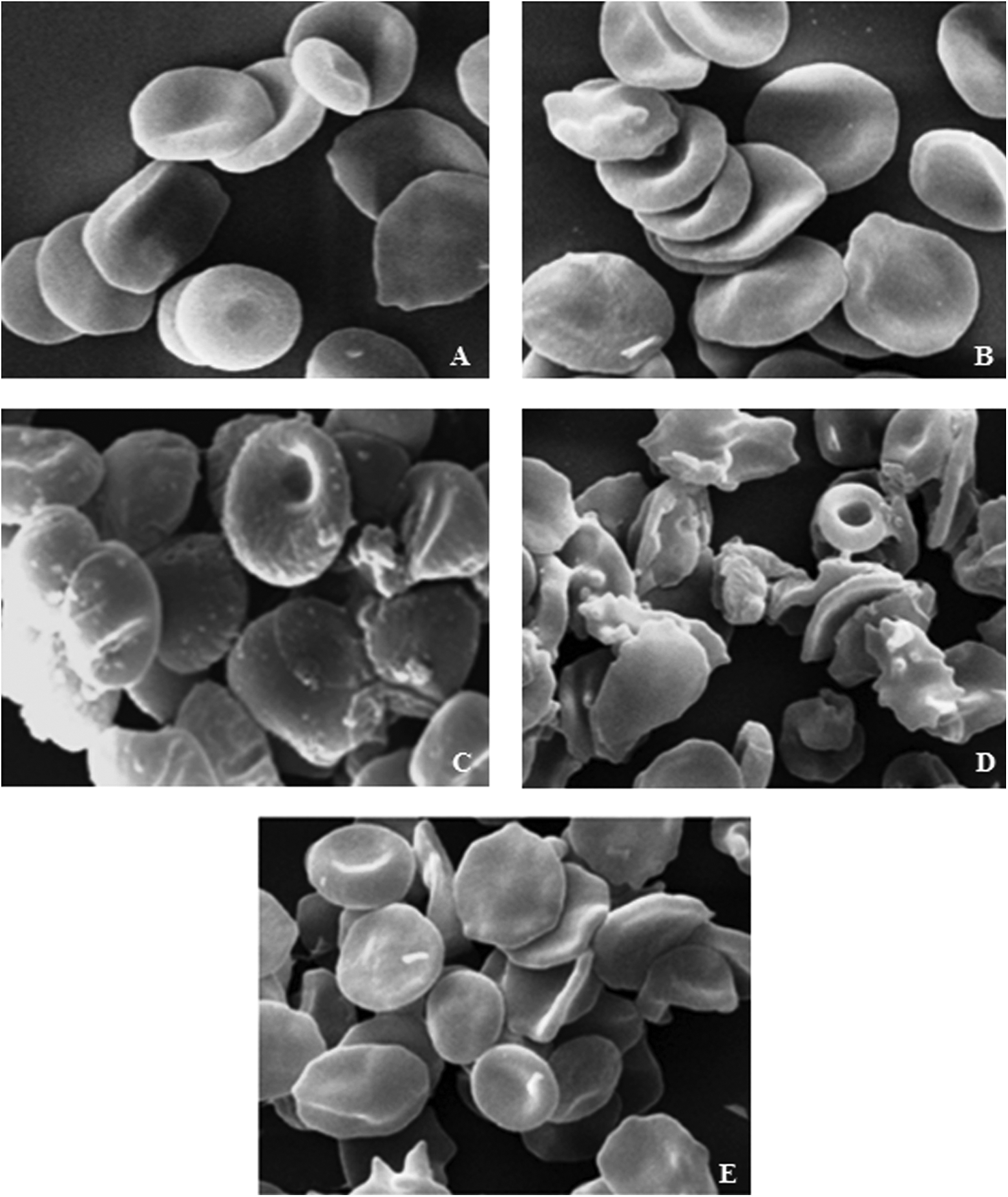
Scanning electron micrograph images from the erythrocytes of treated rats. Control (a) and GSP (b) showed normal micrographs of RBC; (c and d) Cd group show the presence of acanthocytes (A), fragmented RBC-schistocytes (SZ), spherocytes (D), discocyte (D) and echinocytes (E). Most of the erythrocytes showed a biconcave shape. GSP pretreated rats (e) showed the recovered RBCs. ×3000. Bars 10–20 nm. RBC: red blood cell; GSP: grape seed proanthocyanidins.
Discussion
Effective metabolism and functioning of erythrocytes required a balancing array of enzymes, proteins, carbohydrates, lipid anions and cations in the cells. Degenerative changes in haemoglobin, cell membranes and enzymes required for normal erythrocyte functions are because of the imbalance between the erythrocyte components due to the inadequate ability of erythrocytes to cope up with oxidative stress induced by Cd. From prevention of the oxidative reactions, there is a great deal of interest in the use of GSP as an effective antioxidant in the preventive strategies. BW changes between various treatments are used to analyse the deterioration of the rat’s general health condition. In Cd-treated rats, the reduction in BW gain and decreased water and food intake clearly reflects the Cd-induced metabolic alterations and oxidative stress, which influences the absorption of food, shows growth retardation and decreased water and food intake in Cd-treated rats. GSP pre-treated rats showed a significant restoration of the altered BW changes induced by Cd.
The blood is a vital fluid, which contains a homeostatic balanced concentration of RBCs, WBCs and PLTs. ROS have been implicated in the mechanism of damage of the RBCs in Cd-treated rats with the result of haematological complications, which mainly consist of abnormalities in the function, morphology and metabolism of haematological parameters. 43 In our study, a substantial diminution in the levels of WBC, RBC, PLT and HCT and decreasing concentration of HGB, MCH, MCV and MCHC was found in Cd-exposed rats. MCV (Table 2) signifies that the size of RBCs was reduced, indicating microcytic anaemia due to iron deficiency. 44 Due to Cd-induced oxidative stress, a large amount of O2 .− and H2O2 is liberated from the neutrophils into circulation, which leads to the influx of ROS in erythrocytes. Exposure to Cd induces anaemia associated with a decrease in RBCs counts and induction of oxidative damage and lipid peroxidation in blood and RBCs. Also Cd damages the synthesis of erythropoietin, a hormone used to promote the formation of RBCs. 43 Decrease in WBC, MCHC, MCV and MCH might suggest the decrease in immunity in fighting against foreign substances, showing the further ability of Cd intoxication to suppress the activity of haematopoietic tissues, impair erythropoiesis and accelerate erythroclasia because of the altered RBCs membrane permeability, increased RBCs mechanical fragility and/or defective Fe metabolism. 45 In our findings, the pretreatment of GSP in Cd-treated rats showed a significant renewal of these haematological parameters by bringing them back to near normal levels due to the presence of the 3- and 5-hydroxyl groups with 4-oxo group functions as electron donors to form bonds with electrophilic ions, thereby help in the recoupment of the antioxidant defense system and protecting heme from Cd-induced oxidative stress.
AST, ALT and ALP secreted from the lysosomes are one of the markers for assessing cellular damage 46 and are considered among the most sensitive markers of erythrocytic injury. 67 When the integrity of the lysosomal membrane changes and/or the membrane of the lysosomes is ruptured by deleterious influence, these acid and alkali dehydrolyses enter the blood stream, inducing a transient increase in the activity of lysosomal enzymes into the plasma. In the present study, increased levels of AST, ALT, ACP, γGT and LDH caused by Cd are due to increased lipid peroxidation, which has the ability to cause the erythrocyte damage. Cd causes structural and functional damage to the cell membrane and increases the membrane permeability, leading to the leakage of cytoplasm enzymes into the blood. 7 In our finding, GSP regulates the action of these enzymes by stabilizing the cellular membranes, which prevent the leakage of these enzymes into the blood via its hydrogen donation capacity from the phenol to the free radical-containing compounds. 7 This indicates that GSP has the capability to effectively stabilize the cellular membranes and protects the eythrocytes against the toxic effects of Cd.
Lipid peroxidation is said to be the outcome of free radical-mediated chain oxidative reaction of membrane polyunsaturated fatty acids. It is a basic cellular deteriorating process induced by oxidative stress and occurs rapidly in tissues rich in highly oxidizable polyunsaturated fatty acids. 47 Cd administration in the experimental rats resulted in a substantial increase in the degree of plasma TBARS and hydroperoxides, MDA and PC, in erythrocyte membranes through the propagation of lipid peroxidation and the changes in the cellular antioxidant defense system that coincides with the results of Mahua et al. 48 The oxidative stress induced by Cd yields various LH and secondary products, which ultimately results in the structural disruption of membrane lipid bilayer and produce deleterious effects on the natural processes of membrane-bound proteins. Administration of GSP markedly attenuated the elevation of TBARS and other lipid peroxidative derivatives in plasma and erythrocyte membranes as GSP scavenges the toxic free radicals and other reactive intermediates by binding with Cd cations that effectively reduces their absorption and the extent of their oxidative damage and thereby limiting the propagation of lipid peroxidation. 7
NO is recognized as a physiological messenger molecule generated from
In the present study, there was a significant decrease in the levels of GSH, vitamins C and E in the plasma and erythrocyte membranes in Cd-intoxicated rats. Our results are in line with the findings of Mahua et al. 48 During Cd exposure, free radical generation is enhanced and GSH acts to counteract these excess oxyradicals, resulting in the depletion of GSH level as it forms complexes with GSH through the free sulphydryl group and depletes the intracellular sulphydryl groups (SH groups), thereby decreasing its level in the tissues of rats. 52 Vitamins C and E are the antioxidants having synergistic action in scavenging oxygen-derived free radicals, and these vitamins are likely to be the most susceptible to free radical oxidation. The decrease of vitamin C in Cd-treated rats damages the cell membranes since they are implied in the regeneration of α-tocopherols. Our results indicate that pretreatment of GSP increased the level of thiol group by the protection of SH groups in GSH from oxidative damages via the free radicals quenching action of the di-OH (catechol) structure in the B ring of proanthocyanidin. 53 GSP was capable of reversing the decline of vitamin C and vitamin E, through its potent antioxidant and metal chelating activity that minimizes the usage of these antioxidants, thus restoring their levels. The chain breaking action of proanthocyanidins within the core of the phospholipid bilayer saves vitamin E and recycles it from the tocopheroxyl radical through an H-transferring mechanism, thereby behaving as a sacrificial antioxidant. Moreover, the metal chelating property of GSP increases vitamin E content by preventing the involvement of haemoglobin iron in lipid peroxidation processes. 54
SOD, CAT and GPx participate an integral role in the antioxidative stress defenses of the cells by protecting against ROS toxicity and lipid peroxidation. SOD catalyzes the conversion of superoxide radicals to H2O2, whilst CAT converts H2O2 into H2O. Thus, these antioxidant enzymes can alleviate the toxic effects of ROS. In this study, erythrocyte SOD, CAT and GPx activities were decreased in the Cd-treated group, which may be assigned to the saturation of SOD during the process of converting O2 − to H2O. 55 The CAT decrease may involve CAT saturation or inhibition during the breakdown of free radicals and H2O2. 55 Inhibition of GPx activities is accompanied by a depletion of GSH, which may result in oxidative stress. Reduced GSH, in conjunction with GPx and GST, is responsible for the GSH redox cycles that maintain the redox status of tissues and protect structural and regulatory proteins against ROS-induced damage. 56 G6PD is an important enzyme of hexosemonophosphate shunt. It converts one molecule of glucose-6-phosphate into 6-phosphogluconolactone in the presence of Mg2+, Mn2+ and Ca2+ ions, and subsequently, NADP+ is reduced to NADPH. A subsequent reduction of the G6PD activity in Cd-induced rats showed an impaired generation of NADPH, which is required for the reduction of GSSG to GSH. 56 The formation of Cd–SH complex with SH groups of the enzyme leads to a decrease in the activities of GR and G6PD and depletion of GSH level. GSP pretreatment increased the level of these enzymatic antioxidants, probably due to the protection of SH groups in GSH from oxidative damages due to the presence of two hydroxy groups in the 30 and 40 positions on the B ring, resulting in stability to the radical formed mainly in the 3′ position.
In the present study, a significant decrease in the activities of membrane-bound ATPases in the erythrocyte was observed in Cd-treated rats. The membrane-bound Na+/K+-ATPase and Ca2+/Mg2+-ATPase are responsible for the transfer of Na+/K+ and Mg2+/Ca2+ across the cell membranes at the expense of ATP by hydrolysis. 57 Cd-induced lipid peroxidation by free radicals decreases the activity of Na+/K+-ATPase and Ca2+/Mg2+-ATPase. In addition, Na+/K+-ATPase and Ca2+/Mg2+-ATPase are SH-containing enzymes and these −SH groups are indispensable to their activities. 57 Cd inhibits the ATPase activity due to its binding with −SH groups at the active sites of the enzymes, either due to oxidation by free radicals or due to conjugation by reactive chemicals that produced deleterious effects on the membrane proteins. The maintenance of the cation gradient by these enzymes is of fundamental importance in the control of hydration, cellular volume, nutrient uptake, cell fluidity, growth and differentiation. 58 Administration of GSP in Cd-intoxicated rats in our study significantly reduced the lipid peroxidation in erythrocytes and sustained the activities of membrane-bound enzymes by preventing the oxidation of −SH groups through the free radical quenching action of di–OH (catechol) structure in the B ring of GSP.
In the present study, Cd increased the concentration of free iron and H2O2 within erythrocytes, which increases the toxic hydroxyl radical (OH.) and induces the lipoperoxidation of erythrocytes membrane and enhancement of its permeability to calcium, which ultimately lead to erythrocyte death and anaemia. H2O2 acted as a positive regulator of erythrocytes free iron and calcium. Although the role of calcium channels in free iron uptake by the erythrocytes remains unclear. Nandini and Anuradha have speculated about differential calcium regulation by H2O2 or other ROS via either L-type or T-type calcium channels. 59 In our study, GSP counteracted Cd-induced increase in H2O2 and free iron and restored the Ca2+ elevation in erythrocytes as compared to Cd-treated rats. According to our findings, these effects could be due to the presence of functional groups involving both hydroxyl groups of ring-B and the 5-hydroxy group of ring-A in GSP, which used to chelate Cd and thus prevent the catalytic breakdown of H2O2 and its ability to chelate free iron, calcium and scavenge H2O2.
Erythrocyte osmotic fragility test refers to the propensity of erythrocytes to haemolyse when they are subjected to osmotic stress by being placed in a hypotonic solution. Administration of Cd results a significant increase in RBC osmofragility due to the increased lipid peroxidation-mediated reactive oxidative damage to the erythrocyte membrane. Increased oxidative stress and decreased antioxidant status result in increased erythrocyte deformability, RBC membrane lipoperoxidability and consequently lead to an increased osmotic fragility and reduced lifespan in Cd-treated rats. This shows that erythrocytes are less stable in hypotonic solution and might be an indication of membrane fluidity and intravascular haemolysis. Collectively, Cd intoxication might lead to anaemia as a consequence of suppressing the activity of haematopoietic tissues, impaired erythropoiesis, accelerated erythroclasia because of the altered RBCs membrane permeability, increased RBCs mechanical fragility and/or defective Fe metabolism. 45 GSP pretreated rats significantly reduced the RBC osmofragility due to the presence of tannins in grape seeds. Tannins bind to the membrane surface of cells and induce cluster and raft formation 60 that may prevent the erythrocytes swelling, release of haemoglobin and decreases membrane fluidity in both the hydrophobic and hydrophilic parts of erythrocyte membranes, this shows that GSP reduced the haemolysis to some extent, as compared to Cd-treated rats.
In the present study, the response of lymphocyte DNA after treatment with Cd and GSP was evaluated by the comet assay. In our study, the level of DNA damage was significantly increased in the lymphocytes of Cd-treated rats. Cd-induced oxidative stress through the generation of reactive oxygen and nitrogen species leads to the lipid peroxidation and DNA damage. 61 In the present study, Cd leads to the formation of OH. and O2 .−, which has the potential to react with DNA and cause the formation of strand breaks that can be detected by comet assay. According to Whiteman et al., semiubiquinones present in mitochondria are prone to transfer one electron to molecular oxygen to form superoxide ions, providing a potential mechanism for Cd-induced generation of ROS, which results in DNA damages, such as DNA strand breaks and base modifications. 62 It is eminent that Cd at low concentrations affects the incision step of the repair process. The growth in the comet tail length of the Cd-exposed lymphocytes is due to DNA strand breaks induced by Cd. 61 GSP prevented the DNA oxidative damage in various tissues; however, the exact mechanism by which proanthocyanidins protected against Cd-induced DNA damage in lymphocytes is not well characterized. One probable account for the protection against DNA damage is the treatment with proanthocyanidins would allow the interception of free radicals generated by Cd before they turn over DNA and induce oxidative damage. Moreover, Ray et al. 63 suggested that in vivo protection of DNA damage by proanthocyanidins might be due to detoxification of cytotoxic radicals and presumed contribution to DNA repair.
The morphological and ultrastructural examination revealed many changes in RBC of Cd-treated rats. Cd-induced oxidative stress damages cell membrane, leading to the alterations in cell rigidity and form. This oxidative stress leads to echinocyte formation because of membrane damage, which in turn contributes to the change in whole RBC structural conformation and its functioning. 64 In the present study, fragmented RBCs (schistocytes) were observed in Cd-treated rats due to alterations in the cytoskeleton (membrane proteins and/or lipids) of RBC, thus affecting the surface area of the cell. Echinocytes and discocyte are formed when the compound inserts into the inner monolayer, acanthocytes and spherocytes are produced when it locates into the outer moiety. Cd damages the membrane structure, ion permeability and cell metabolism of erythrocytes, thus may cause morphological damage in erythrocyte formation. Altogether, these changes induced by Cd were effectively attenuated by the pretreatment with GSP via the presence of flavan-3-ol molecule, which contains both hydrophobic and hydrophilic residues and allow these compounds to interact with phospholipid head groups and be adsorbed onto the surface of membranes. These interactions can lead to the modification of membrane properties, leading to changes in the regulation of membrane-bound enzymes and receptors. 65
GSP prevents the ROS formation via hydrogen donation from the phenol to the radical-containing compound. The electronic configuration of GSP allows free release of electrons to R. such as superoxide anion, hydroxyl and NO radicals with which the free radical reactivity of ROS is reduced by binding with the GSP. Also the presence of electron-donating groups attached to the aromatic ring of GSP such as –CH3 and –OH ought to increase the ease of hydrogen atom calculation, enable them to undergo redox reaction, scavenge the free radicals more easily and protect them against the oxidative insult imposed by Cd. 66
In conclusion, the present study shows that the treatment of rats with Cd caused oxidative stress-mediated damage in the erythrocyte and lymphocytes of rats. Cd causes anaemia, loss of membrane function by enhancing the lipid peroxide concentration as well as alterations in the enzymatic and non-enzymatic antioxidants. GSP has the ability to provide protection against Cd-induced oxidative damage in rat erythrocytes and DNA damage in lymphocytes possibly through its direct as well as indirect antioxidant activity, free radical scavenging and metal-chelating properties. Although the possible rescue mechanism of GSP on erythrocytes is not fully apparent, further studies are necessary to ascertain the exact mechanism of GSP protection against Cd-induced oxidative damage.
Footnotes
Acknowledgments
The authors would like thank to the Professor and Head of the Department of Zoology, Annamalai University for providing laboratory facilities and UGC MANF (MANF-2013-14-MUS-JAM-23333), New Delhi.
Conflict of interest
The authors declared no conflicts of interest.
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.