
Abstract
Chlorpyrifos (CPF), methyl parathion (MPT), and malathion (MLT) are among the most extensively used organophosphate (OP) pesticides in India. DNA protein cross-links (DPC) and DNA strand breaks are toxic lesions associated with the mechanism(s) of toxicity of carcinogenic compounds. In the present study, we examined the hypothesis that individual and interactive genotoxic effects of CPF, MPT, and MLT are involved in the formation of DPC and DNA strand break. The DNA strand break was measured by comet assay and expressed as DNA damage index, while DPC estimation was carried out by fluorescence emission assay. The results showed that exposure of rat lymphocytes with CPF, MPT, and MLT caused significantly marked increase in DNA damage and DPC formation in time-dependent manner. MPT caused the highest damage, and these pesticides do not potentiate the toxicity of each other.
Keywords
Introduction
Pesticides are a very important group of environmental pollutants used in intensive agriculture for protection against diseases and pests. The estimated annual application is more than 4 million tons, but only 1% of this reaches the target pests. 1 It has been estimated that 3,000,000 cases of severe poisoning and 220,000 deaths are caused globally every year due to pesticides poisoning. 2 Besides being neurotoxic, 3 organophosphate insecticides (OPIs) exert their toxic effects on many other tissues and organs. A number of pesticides have been tested in a wide variety of mutagenic assays for gene mutation, chromosomal aberration, and DNA damage. 4 OP pesticides have demonstrated genotoxic, alkylating, and clastogenic properties; thus they are potentially mutagenic and clastogenic. 5,6
Pesticides induced oxidative stress in mammalian systems. 7 OP compounds, methyl parathion (MPT), malathion (MLT), monocrotophos, and acephate have been shown to be genotoxic. 8 –10 MLT is reported to induce oxidative stress, cytotoxicity, and genotoxicity in human liver carcinoma (HepG2) cells. 11 It has been demonstrated that DNA damage can occur in lymphocytes and hepatocytes from a single low-level exposure to diazinon and dichlorvos. This study shows that some, but not all of, OPs can cause DNA damage at levels subthreshold for acute toxicity. 12 Study on the effect of OPIs and their metabolite, oxon, on human spermatozoa revealed that OPIs alter semen quality, sperm chromatin, and DNA at different stages of spermatogenesis. 13 Chlorpyrifos (CPF)-induced DNA damage in rat brain and liver is also reported. 5 Oxidative stress and DNA damage are possibly linked to pesticides-induced adverse health effects in agricultural workers. 14 In vitro exposure of human blood lymphocytes with carbofuran induces significant DNA damage in concentration-dependent manner and increased frequency of micronuclei exhibit its genotoxic potential. 15 A study in occupational North Indian suburban population shows pesticides-induced biochemical alterations in antioxidant enzymes like superoxide dismutase, catalase, glutathione-S-transferase, and glutathione peroxidase. 16 Studies on soybean workers in Espumoso region who are exposed to a wide combination of fungicides, herbicides, and insecticides show high DNA damage in peripheral leukocytes. 17
The DNA damaging effect could follow from the induction of DNA strand breaks and/or alkali-labile sites (ALS), the latter from the formation of DNA–DNA and/or DNA protein cross-links (DPC). A DPC is created when a protein becomes covalently bound to DNA when cells are exposed to DNA-damaging agents. 18 The formation of DPCs was originally demonstrated for bacterial and mammalian cells that were heavily irradiated with ultraviolet light. 19,20 It was subsequently shown that DPCs are produced by a number of chemical and physical agents such as aldehydes, metal ions, 21 anticancer drugs, 22 and ionizing radiation. 23 Recent studies with defined DPC substrates have begun to unravel the molecular mechanism by which cells respond to this type of potentially deleterious DNA damage. 24 In bacteria, DPCs inhibit the replication of plasmid DNA in vivo, 25 –27 and replication forks stalled by DPCs are likely reactivated by RecBCD-dependent homologous recombination and the subsequent action of PriA helicase to continue DNA synthesis through DPCs. 26,28 DPCs containing small (but not large) cross-linked proteins (CLPs) are removed from DNA by nucleotide excision repair (NER). 26,28,29
CPF (O,O-diethyl O-3,5,6-trichloro-2-pyridyl phosphorothioate), MPT (O,O-dimethyl O-4-nitrophenyl phosphorothioate), and MLT (S-1,2-dicarbethoxy ethyl O,O-dimethylphosphorodithioate) are diethylorganophosphorothionate insecticides extensively used for agriculture and domestic purposes in India. Simultaneous exposure to two or more pesticides can take place in the environment and/or under occupational conditions; the investigation of interactions between pesticides is an important problem in modern toxicology. Since CPF, MPT, and MLT are widely used OP pesticides in India, their overlapping application may lead to combined exposure that may potentiate the effect of each other. Therefore, looking at the widespread use of these studied OP pesticides in agriculture and domestic use, the present study was aimed to evaluate the DNA damage and DPC formation induced by CPF, MPT, and MLT, individually and mixture in in vitro system using the peripheral blood lymphocytes of rats. Different agents induce DPCs by different mechanisms, and the biology of these lesions is complicated by several factors. However, present studies will also throw light on how studied pesticides induce DPC formation in mammalian system.
Materials and methods
Chemicals
Normal melting agarose (NMA), low-melting agarose (LMA), Triton X-100, ethylenediamine tetraacetic acid (EDTA), tris (hydroxymethyl)-aminomethane (Tris), ethidium bromide, endonuclease III, and formamidopyrimidine-DNA glycosylase, oxidized cytochrome C, phorbol 12-myristate 13-acetate, phenol red ,and hydrogen peroxide (H2O2) were purchased from Sigma Chemicals Inc. (St Louis, Missouri, USA). Sodium chloride (NaCl), disodium hydrogen phosphate, sodium dihydrogen phosphate, sodium hydroxide (NaOH), Hisep, dimethylsulfoxide (DMSO), Roswell Park Memorial Institute (RPMI) 1640, trypan blue, 4-(2-hydroxyethyl)-piperazine-1-ethanesulphonic acid buffer, potassium hydroxide, bovine serum albumin, and potassium chloride were purchased from Merck Chemicals (Germany). MPT, CPF, and MLT were kindly gift from Devidayal (Sales) Limited (Mumbai, Maharashtra, India).
Experimental animals
Adult male albino rats of Wistar strain (Rattus norvegicus), weighing 150 ± 20 g, were used in all experiments. Rats were obtained from animal facility division of All India Institute of Medical Sciences, New Delhi, India, and were maintained in the department animal room with a 12-h light/12-h dark cycle) and 25°±2°C temperature condition and were fed on standard pellet diet and tap water ad libitum. Rats were acclimatized for 1 week prior to the start of experiment. The care and maintenance of animals were as per the approved guidelines of the “Committee for the Purpose of Control and Supervision of Experiments on Animals” (India).
Lymphocytes isolation
Peripheral blood was collected from healthy male Wistar rats (150 ± 20 g) from eye orbital in vacutainer test tubes containing EDTA (Becton-Dickinson, Cedex, France). Blood samples were layered on the top of a Ficoll solution (1.077 g/mL), and the supernatant containing the leukocytes was removed after sedimentation of erythrocytes at 1000 g for 10 min at room temperature. Lymphocytes sediment at the interface of the Ficoll layers were collected and washed twice with phosphate-buffered saline (PBS), pH 7.4 at 20°C. The cell viability was checked by trypan blue exclusion test 30 and was found to be about 95%. The final concentration of the lymphocytes was adjusted to 1–3 × 105 cells/mL by adding RPMI 1640 to the single cell suspension.
Pesticide treatment
CPF (4 h median lethal concentration (LC50) = 0.2 mg/L), 31 MPT (4 h LC50 = 0.135 mg/L), 32 and MLT (4 h LC50 > 5.2 mg/L) 33 individually and in mixture were taken from DMSO stock solutions (LC50) and added to the lymphocyte suspension to give final concentrations of 1/10 and ¼ LC50 individually and in mixture (equivalent to 1/30 and 1/12 LC50 of individual pesticides). The control cells were treated with DMSO without the OPs, which did not affect the processes under study. To examine DNA double-strand breaks (DSBs) DNA single-strand breaks (SSBs), and DPC formation, the lymphocytes were incubated with pesticides for 2, 4, 8, and 12 h at 37°C in a carbon dioxide incubator, and the obtained results were compared with the control lymphocyte cells.
DNA SSB and DSB measurement
DNA SSB was measured by alkaline version of comet assay according to protocol followed by Sasaki et al. 34 After exposures, lymphocytes were mixed with LMA (2% in PBS) and layered on slides precoated with NMA (1% in PBS). After overnight incubation in cold lysis buffer at 4°C (2.5 M NaCl, 0.1 M disodium-EDTA, 0.01 M Tris-hydrochloric acid (HCl), 1% Triton X-100, 10% DMSO, pH = 10), slides are placed in an electrophoresis tank containing electrophoresis solution (0.3 M NaOH, 1 mM EDTA). Electrophoresis should be run for 30 min with constant voltage setting, at around 0.8 V/cm (measured between the electrodes across the platform carrying the slides). DNA DSB was measured by neutral version of comet assay having electrophoresis buffer (300.0 mM sodium acetate, 100.0 mM Tris-HCl) of pH 8.5, according to protocol followed by Fracasso et al. 35
After electrophoresis, slides were neutralized by adding Tris-HCl buffer (0.4 M, pH 7.5). The slides were rinsed in ethanol and dried for later analysis. The whole procedure was performed in dim light to minimize artifactual DNA damage. Just before visualization, each slide was stained with 50 µL of ethidium bromide (2 µg/mL), rinsed with water, and covered with a cover slip. A total of 50 cells were scored per slide (25 from each replicate slide). Analyses were performed on the basis of the type of comet visualized on the slide. The nuclei were counted and divided into five types as stage 0, 1, 2, 3, and 4 as shown in Figure 1. The DNA damage index was calculated as #0 + #1 + #2 + #3 + #4/ # of cell scored. Two independent experiments were conducted in each treatment. Slides were viewed under fluorescence microscope (Nikon Eclipse Ti Microscope, Tokyo, Japan). Analyses were performed at 100× magnification, with a Nikon Eclipse Ti microscope equipped with an excitation filter of 515–560 nm and barrier filter of 590 nm.
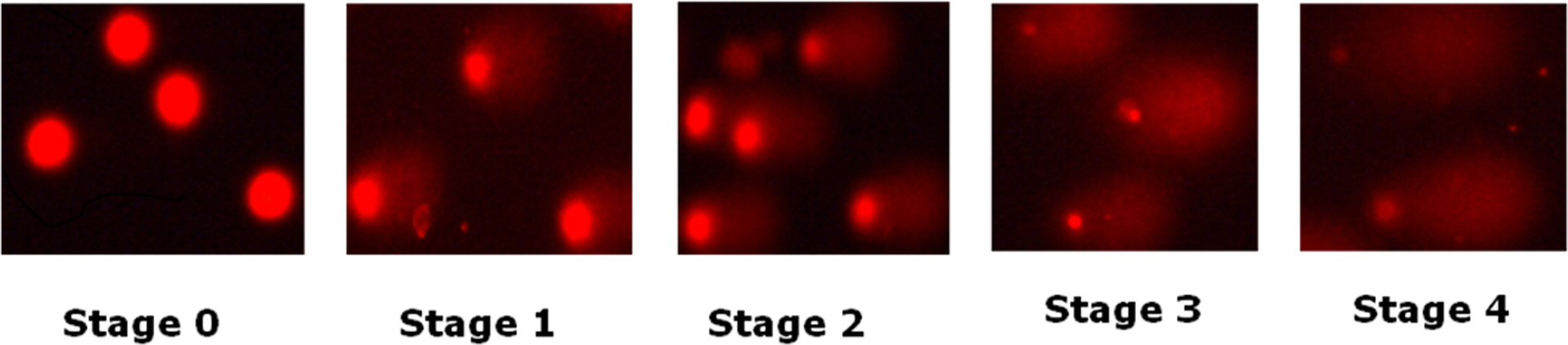
Showing different stages of comet.
DPC estimation
DNA DPCs were measured by the method followed by Zhitkovich and Costa. 36 Briefly, the assay is based upon the binding of sodium dodecyl sulfate (SDS) to proteins and its lack of binding to DNA. Fragments of protein-free DNA and protein-bound DNA were easily separated, since free DNA remained in the supernatant, while protein-linked DNA precipitated with protein-bound SDS when the cation was changed from sodium ion to potassium ion. Thus, the amount of DNA in the SDS pellet provides a direct measurement of DPC. DNA was mixed with 1 mL freshly prepared Hoechst dye reagent (200 ng/mL) at pH 7.5 to determine the amount of DNA in samples. Fluorescent measurements were made using a Spectrofluorimeter (Molecular Devices, Sunnyvale, California, USA) with 360 nm excitation and 450 nm emission filters. We calculated the amount of DPC as the ratio detected in exposed white blood cells to the total DNA in the same amount of cells.
Statistical analysis
Results are expressed as mean ± SE of six set of observation. Statistical analyses were performed using Statistical Package for Social Sciences Version 2.0 (SPSS Inc., Chicago, Illinois, USA) statistical software. All the statistical analyses were performed using one-way analysis of variance with post hoc Bonferroni’s multiple comparison test applied across the treatment groups. The value of p < 0.05 was considered statistically significant.
Results
Percentage viability estimation
Results of the present study showed that lymphocytes exposed to 1/10 and 1/4 LC50 equivalents of CPF, MPT, MLT and their mixture caused time-dependent decrease in the viability of rat peripheral blood lymphocytes (Table 1). Data show 7–12% and 15–18% decrease in cell viability after 2 h exposure of rat lymphocytes with 1/10 and 1/4 LC50 dose of CPF, MPT, MLT, and their mixture, respectively, when compared with control. After 4 h exposure of rat lymphocytes with 1/10 and 1/4 LC50 equivalent of CPF, MPT, MLT, and their mixture caused 10–12% and 16–27% decrease in cell viability, respectively, as compared to control. Incubation of rat lymphocytes with 1/10 and 1/4 LC50 equivalent of CPF, MPT, MLT, and their mixture for 8 h caused 18–22% and 21–27% decrease in cell viability, respectively, as compared to control. Similarly 12 h exposure of rat lymphocytes with 1/10 and 1/4 LC50 dose of CPF, MPT, MLT individually and in mixture caused 26–30% and 22–34% decrease in cell viability, respectively, as compared to control.
Estimations of percentage viability in rat’s lymphocyte after exposure of CPF, MPT, and MLT individually and in mixture for 2, 4, 8, and 12 h.a
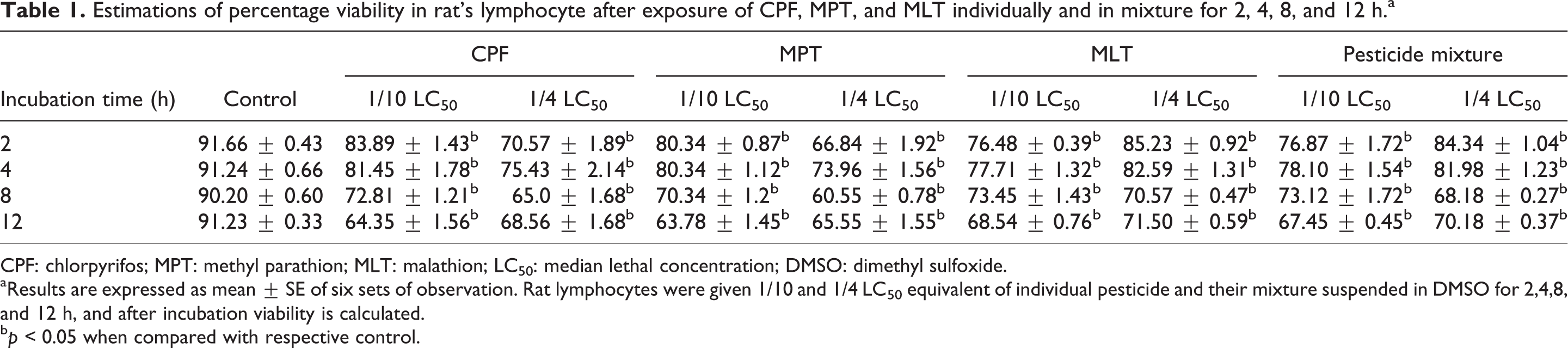
CPF: chlorpyrifos; MPT: methyl parathion; MLT: malathion; LC50: median lethal concentration; DMSO: dimethyl sulfoxide.
aResults are expressed as mean ± SE of six sets of observation. Rat lymphocytes were given 1/10 and 1/4 LC50 equivalent of individual pesticide and their mixture suspended in DMSO for 2,4,8, and 12 h, and after incubation viability is calculated.
b p < 0.05 when compared with respective control.
Estimation of DNA SSBs
Results of the DNA SSB or alkaline comet assay showed that strand breaks contributed to the observed increase in the damage index (in terms 0, I, II, III, and IV stage) in the presence of individual and combined OP exposure. The data showed that in vitro exposure of rat lymphocytes with 1/10 and 1/4 LC50 concentrations of CPF, MPT, MLT, and their mixture for 2 h caused 289–511% and 411–532% increase in DNA damage index, respectively, when compared with the control. When the lymphocytes were incubated with 1/10 and 1/4 LC50 concentrations of CPF, MPT, MLT, and their mixture for 4 h caused 386–422% and 477–905% increase in the DNA damage index, respectively, as compared to control lymphocytes. Similarly in vitro exposure of rat lymphocytes with 1/10 and 1/4 LC50 concentrations of CPF, MPT, MLT, and their mixture for 8 h and 12 h caused 338–981, 495–1114, 375–933, and 488–983% increase in DNA damage index, respectively, as compared to control (Table 2).
Estimations of DNA SSB in rat’s lymphocyte after exposure of CPF, MPT, MLT individually and in mixture for 2, 4, 8, and 12 h.a
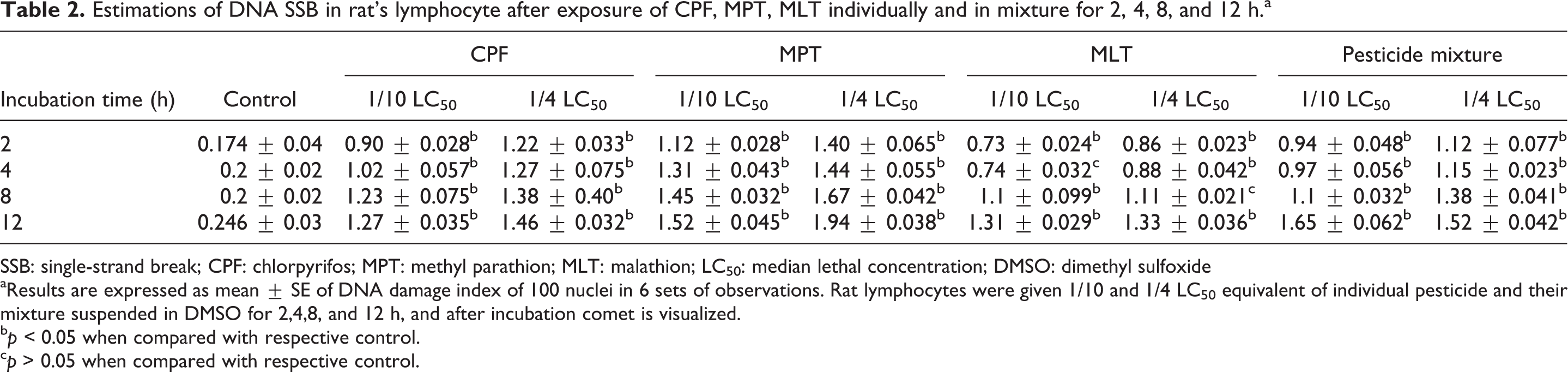
SSB: single-strand break; CPF: chlorpyrifos; MPT: methyl parathion; MLT: malathion; LC50: median lethal concentration; DMSO: dimethyl sulfoxide
aResults are expressed as mean ± SE of DNA damage index of 100 nuclei in 6 sets of observations. Rat lymphocytes were given 1/10 and 1/4 LC50 equivalent of individual pesticide and their mixture suspended in DMSO for 2,4,8, and 12 h, and after incubation comet is visualized.
b p < 0.05 when compared with respective control.
c p > 0.05 when compared with respective control.
Estimation of DNA DSBs
Results of the DNA DSB or neutral comet assay showed that there was no difference between DNA damage index in the alkaline and neutral comet assay, which indicates that strand breaks contributed to the observed increase in the damage index (in terms 0, I, II, III, and IV stage) in the presence of individual and combined OP exposure. The data showed that in vitro exposure of rat lymphocytes with 1/10 and 1/4 LC50 concentrations of CPF, MPT, MLT, and their mixture for 2 h caused 306–522 and 378–700% increase in DNA damage index, respectively, when compared with the control. When the lymphocytes were incubated with same concentration of pesticides for 4 h, 270–555 and 340–600% increase in the DNA damage index was observed in CPF, MPT, MLT, and their mixture-treated lymphocytes, respectively, as compared to control lymphocytes. Similarly in vitro exposure of rat lymphocytes with 1/10 and 1/4 LC50 concentrations of CPF, MPT, MLT, and their mixture for 8 h and 12 h caused 450–625, 455–725, 392–560, and 432–676% increase in DNA damage index, respectively, as compared to control (Table 3).
Estimations of DNA DSB in rat’s lymphocyte after exposure of CPF, MPT, MLT individually and in mixture for 2, 4, 8, and 12 h.a

DSB: double-strand break; CPF: chlorpyrifos; MPT: methyl parathion; MLT: malathion; LC50: median lethal concentration; DMSO: dimethyl sulfoxide.
aResults are expressed as mean ± SE of DNA damage index of 100 nuclei in 6 sets of observations. Rat lymphocytes were given 1/10 and 1/4 LC50 equivalent of individual pesticide and their mixture suspended in DMSO for 2,4,8, and 12 h, and after incubation comet is visualized.
b p < 0.05 when compared with respective control.
c p < 0.0001 when compared with respective control.
d p < 0.001 when compared with respective control.
e p > 0.05 when compared with respective control.
Estimation of DPC
Results of the present study showed that in vitro exposure of rat lymphocytes with 1/10 and 1/4 LC50 dose of OP pesticides, CPF, MPT, and MLT, alone or in combination for 2, 4, 8, and 12 h caused significantly marked increase in DPC formation, as evidenced by the increase in the florescence. After exposing the lymphocytes of rats for 2 h with 1/10 and 1/4 LC50 dose of CPF, MPT, MLT, and their mixture caused 12–47 and 21–57% increase in DPC formation as compared to control, respectively. Similarly, 4 h exposure of rats lymphocytes with 1/10 LC50 and 1/4 LC50 dose of CPF, MPT, MLT, and their mixture caused 0.4–19 and 0.5–41% increase in DPC formation as compared to control, respectively. Similarly, 8 and 12 h exposure of rats lymphocytes with 1/10 LC50 and 1/4 LC50 dose of CPF, MPT, MLT, and their mixture caused 20–56, 24–89, 83–106, and 88–148% increase in DPC formation as compared to control, respectively (Table 4).
Estimations of DPC formation in rat’s lymphocyte after exposure of CPF, MPT, MLT individually and in mixture for 2, 4, 8, and 12 h.a

DPC: DNA protein cross-link; CPF: chlorpyrifos; MPT: methyl parathion; MLT: malathion; LC50: median lethal concentration; DMSO: dimethyl sulfoxide; BSA: bovine serum albumin.
aResults are expressed as mean ± SE of six sets of observations. The amount of DPC is expressed as the ratio detected in exposed lymphocytes to the total DNA in the same amount of cells. Rat lymphocytes were given 1/10 and 1/4 LC50 equivalent of individual pesticide and their mixture suspended in DMSO for 2,4,8, and 12 h, and after incubation DPC is visualized.
b p < 0.05 when compared with respective control.
c p > 0.05 when compared with respective control.
d p < 0.001 when compared with respective control.
e p < 0.0001 when compared with respective control.
Discussion
Extensive use of pesticides in crop protection and for household purposes has resulted in their widespread distribution in the environment causing part of the worldwide environmental contamination. Besides their great contribution to the animal and human prevention from vectors of diseases, their use also creates many problems because of their toxicity to nontarget organisms, persistence, and combined effects with other agrobiochemicals and environmental factors. Because of the potential environmental impacts associated with the extensive use of OP pesticides and the target population exposed, an assessment of the effects and risks of accidental or chronic exposure to the pesticides is needed. CPF, MPT, and MLT are commonly used OP pesticides, known to cause DNA damage in tissues and cells of target organisms. However, no report is available in literature on the level of DNA damage when the combined exposure of these pesticides is given. This study was undertaken to evaluate the in vitro genotoxicity of CPF, MPT, and MLT when given individually or together in mixture using the single-cell gel electrophoresis (SCGE) or comet assay.
The biochemical and molecular mechanisms of CPF-, MPT-, and MLT-induced genotoxicity are not fully known, and the results of studies remain often inconclusive. Genotoxicity was reflected in the differential production of the DNA damage and its effects on cellular viability. In the present study, all pesticides significantly reduced rat lymphocyte viability. Particularly, we observed that at 1/4 LC50 concentrations of pesticides produced more than 60% killing of the cells and higher DNA fragmentation which was also reflected by presence of apoptotic nuclei or comets without nuclei (“clouds”). The present study clearly showed that 8 and 12 h exposure of rat lymphocytes with 1/10 1nd 1/4 LC50 dose of CPF, MPT, and MLT caused significantly high level of DNA damage. The number of damaged nuclei of type I, II, III, and IV increased resulting in many fold increase in the damage index on exposure with these pesticides. These results suggest the possibility of induction of DNA breakage and/or ALS and various species of oxidized purines and pyrimidines, which are easily detected by alkaline comet assay due to pesticide exposure. 37 Damage to cellular DNA by lipid peroxidation plays a major role in cell injury and altered cell functions leading to apoptosis. 38 Thus, the role of oxidative stress in DNA damage observed by alkaline comet assay or SCGE herein cannot be ruled out.
The role of reactive oxygen species (ROS) in the production of DNA SSBs is well known. 39 ROS interact with the biological molecules and disrupt the normal synthesis and repair of DNA. This disruption is primarily associated with ROS interaction with the biological molecules and disrupts the normal synthesis and repair of DNA. This disruption is primarily associated with inhibition/inactivation of key proteins as well as DNA repair enzymes induced by ROS damage to these biomolecules. 40 Typically most ROS have a short half-life and cause damage locally, but for example H2O2 has a relatively long half-life and can travel long distances causing DNA damage at distant sites. 41 There have been several investigations in vivo and in vitro on the correlation between toxicant-induced oxidative stress and DNA damage. These studies include the effect of monocrotophos in tissues of rats, 42 MLT on human liver carcinoma cells, 43 CPF on rat tissues and lymphocytes of mice, 44 cypermethrin, pendimethalin, and dichlorvos on Chinese hamster ovary (CHO) cells, 45 diazinon in mouse sperm cells, 46 and the herbicide α-2,4-dichlorophenoxyacetic acid in CHO cells 47 as well as several other environmental mutagens and metals. 48,49
Chromosomal aberration, sister chromatid exchange (SCE) analysis, micronucleus test, DNA elution test, diffusion assay, and comet assay and its modifications are the cytogenetic techniques used in controlled clinical trials to evaluate the primary DNA damage caused by physical and chemical agents. 50–51 The comet assay not only measures DNA lesions but also DNA nicks associated with repair activities occurring through base excision repair or NER. 52,53 It is reported that in vitro exposure of human peripheral blood lymphocyte with lead acetate may induce SSBs and DSBs in DNA as well as cause DPC formation. 54 The exposure of occupational workers to mixture of pirimiphos-methyl, CPF, temephos, and MLT may induce DNA damage, decrease in acetylcholinesterase activity, hepatotoxicity as well as nephrotoxicity. 55
OPIs are genotoxic, inducing gene mutation, deletion, chromosomal aberration, SCEs, and cell transformation. They can also induce DNA adducts such as hydroxymethyl adducts in DNA, CLP with DNA, and biologic macromolecules. Under its influence, structural proteins that normally do not bind to DNA become covalently cross-linked to DNA. In the present study, we found DPC formation in time- and dose-dependent manner following exposure to CPF, MPT, and MLT individually and in mixture in vitro condition as compared to control thus showing its carcinogenic effect. Because DPCs are irreversible and expected to interrupt DNA metabolic processes such as replication, repair, recombination, transcription, and chromatin remodeling, it may possibly lead to a loss of genetic material that may inactivate tumor suppressor genes (p53). DPC formation induced by CPF, MPT, and MLT are probably formed by direct interaction between DNA and proteins. The interaction of OP with DNA is most likely ionic. Indeed, the effect of agents that cause DPCs on DNA replication has been widely investigated. 18 Thus, all known DPC-inducing agents generate other forms of DNA damage in addition to DPCsand direct attribution to mutagenicity and carcinogenicity in exposed individuals.
Different types of tumors and the inhibition of tumor suppressor genes such as p53 (maintain normality of cells) emphasize that DPC may be one of the primary and early lesions of the carcinogenesis process. 56 Several studies have reported that the induction of DPCs by many agents correlates with genetic damage such as SCEs, transformation, and cytotoxicity. 57,58 Thus, DPCs may contribute to the genotoxic effects of many different DNA-damaging agents. It is well known that loss of DNA is an important genetic consequence in many genetic diseases such as Alzheimer’s and Parkinson’s disease as well as responsible for aging and cancer. The rare hereditary diseases characterized by genetic defects of DNA repair mechanisms includes ataxia telangiectasia, Nijmegen breakage syndrome, Werner syndrome, Bloom Syndrome, Fanconi anemia, xeroderma pigmentosum, Cockayne syndrome, and trichothiodystrophy. 59
Conclusion
The results of the present study suggest that CPF, MPT, and MLT can induce SSBs and DSBs in DNA as well as DPC formation. We suppose that the DNA SSBs and DSBs could arise from DNA degradation connected with the cell death. It is also possible that OPs could inactivate some proteins, for example, repair enzymes, so that DNA strand breaks could arise from their inactivation. Under our experimental conditions, we found significant increase in the mean DNA damage index and DPC formation after pesticide exposure compared with the control. Thus, the studied OP might generate oxidative lesions to the DNA bases in exposed rat lymphocytes and thus have genotoxic potential that may alter the expression of some enzymes and further study should be performed to measure their cytotoxic potential.
Research Highlights
In vitro exposure with OP pesticides induced DNA damage in rat lymphocytes. These pesticides caused DNA SSB and DNA DSB in rat lymphocytes. Combined exposure with these pesticides had not shown synergistic effects. In vitro exposure of rat lymphocytes with these pesticides induces DPC formation.
Footnotes
Funding
The financial support of Indian Council of Medical Research, New Delhi, India, in the form of individual research project to Prof. YKG is thankfully acknowledged.