
Abstract
Archaeon Aeropyrum pernix K1 is an obligate aerobic hyperthermophilic organism with C25,25-archeol membrane lipids with head groups containing inositol. Interactions of archaeosomes, liposomes prepared from lipids of A. pernix, with mammalian cells in vitro were studied. In vitro cytotoxicity was tested on five different cell lines: rodent mouse melanoma cells (B16-F1) and Chinese hamster ovary (CHO) cells, and three human cell lines—epithelial colorectal adenocarcinoma cells (CACO-2), liver hepatocellular carcinoma cell line (Hep G2) and endothelial umbilical vein cell line (EA.hy926). Archaeosomes were nontoxic to human Hep G2, CACO-2 and mildly toxic to rodent CHO and B16-F1 cells but showed strong cytotoxic effect on EA.hy926 cells. Confocal microscopy revealed that archaeosomes are taken up by endocytosis. The uptake of archaeosomes and the release of loaded calcein are more prominent in EA.hy926 cells, which is in line with high toxicity toward these cells. The mechanisms of uptake, release and action in these cells as well as in vivo functioning have to be further studied for possible targeted drug delivery.
Introduction
Archaea are unicellular microorganisms that are regarded as one of the three domains of life (apart from Bacteria and Eucarya). 1 They mostly inhabit extreme environments such as hot acidic springs and submarine volcanic fields (with high salinity, very high or very low temperature and pH, low oxygen concentrations and high pressure) and therefore possess unique structural and metabolic features to survive. 2 The lipid composition of cell membrane plays one of the key roles in overcoming destabilizing conditions of such extreme environments. Archaeal lipids have four unique characteristics that increase membrane stability in these conditions: (1) stereochemistry of glycerophosphate groups: sn-glycerol-1-phosphate defines sn-2 and -3 positions of hydrocarbon chain binding, (2) ether linkages between glycerol moiety and hydrocarbon chains, (3) high methyl branching of hydrocarbon chains (isoprenoids and isopranoids) and (4) rigid membrane-spanning bipolar lipids with tetraether core in several archaeal species. 3,4
For the past few decades, lipid bilayer vesicles (liposomes) have been extensively studied for drug delivery. The ability to incorporate hydrophilic, lipophilic and amphiphilic compounds into liposomes (to aqueous core or lipid bilayer) led to the development of drug, vaccine and gene delivery systems, protecting encapsulated drug from physiological media, better biodistribution of drugs, site- and time-specific unloading of cargo in therapeutic concentrations at the site of action and lowering toxic side effects of active compounds. Liposomes that are prepared from natural phospholipids are biologically inert and cause little or no antigenic, pyrogenic, allergic and toxic reactions. 5,6
A major disadvantage of liposomes as drug carriers however is their instability in a gastrointestinal tract and circulatory system; they release their cargo before reaching the target tissue or become prone to reticuloendothelial system elimination, mostly in liver and spleen. 7,8 Different approaches, such as adding poly(ethylene glycol) to liposomes to mask the surface, have been used to overcome this problem. 9 The discovery of new archaeal lipids that form liposomes that are stable in high temperature, low or high pH, resist even in the presence of phospholipases, bile salts and serum media 10 –14 and even in mixtures with conventional diester lipids 15 has led to the development of new potential drug, gene and vaccine delivery systems. Some Archaea species possess unusual bipolar tetraether lipids that bear two hydrophilic head groups located at both ends of a hydrophobic domain that stabilize and rigidify membranes of methanogens, thermophiles and psychrophyles. 16,17 Archaeosomes are also strong adjuvants for the induction of Th1, Th2 and CD8+ T cell responses to the entrapped soluble antigens and are therefore promising self-adjuvanting delivery vehicles for vaccines against intracellular infections and cancers. 18
Archaeon Aeropyrum pernix K1 is the first discovered obligate aerobic hyperthermophilic organism growing at temperatures up to 100°C. It was isolated from a coastal solfataric vent in Japan. It grows optimally at temperatures 90–95°C, pH 7.0 and salinity 3.5%. The core lipids of A. pernix cell membrane consist solely of C25,25-archeol (2,3-disesterterpanyl-sn-glycerol). C25,25-archaetidyl(glucosyl)inositol (AGI) has glucosyl inositol and C25,25-archaetidylinositol (AI) has inositol as polar head group components, representing 91 and 9 mol% of total lipids, respectively. 19 Although A. pernix lacks bipolar tetraether lipids, the archaeosomes prepared from A. pernix lipids show great stability at pH range from 4 to 12 and no apparent gel-to-liquid crystalline phase transition in the temperature range from 0 to 100°C. 20 –22 The length of the C25-isoprenoid hydrocarbon chain may be 20% longer than that of C20-isoprenoid or C18 straight-chain fatty acid, therefore it is assumed that C25,25-archeol-based lipid should be about 20% thicker of other Archaea, although the physiological significance of this state is yet unknown. 23
In the scope of a possible use of A. pernix archaeosomes as a potential drug delivery system we tested in vitro cytotoxicity on various cell lines, rodent and human. Liposomes that are candidates of the drug delivery system have to express the least cytotoxic effects as possible. In the previous studies, liposomes composed of lipids from different archaeal species have been found to be nontoxic or mildly toxic in vitro 24 –26 and nontoxic in vivo. 27 To our knowledge, the presented study is the first report of the interactions of liposomes constituted of A. pernix lipids and cultured mammalian cells.
Endocytotic uptake of A. pernix archaeosomes into cells was also observed, using a confocal fluorescence microscope. In addition, calcein release from archaeosomes inside cells was studied to establish a possible use of archaeosomes as “drug carriers”.
Methods
Archaeosomes
Cells of A. pernix were grown, harvested and lipids were extracted and purified as described previously. 22 Briefly, cells were grown in culture medium consisted of synthetic sea salt, trypticase peptone, yeast extract, sodium thiosulfate and 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid) buffer at pH 7.0. The A. pernix cells were grown in heavy-walled flasks with forced aeration at 92°C. After 40 h, the suspensions were cooled, centrifuged and washed. Lipids were isolated from lyophilized biomass essentially as described by Bligh and Dyer. 28 The lipids were fractionated with adsorption chromatography and polar lipid methanol fraction was used for the preparation of archeosomes. Dry lipid film was hydrated with potassium phosphate buffer (KPB) consisting of 250 nM sucrose, 10 mM phosphate (dipotassium phosphate /potassium hydrogen phosphate) and 1 mM MgCl2, pH 7.4 and vortexed to obtain multilamellar vesicles (MLVs) that were also transformed into small unilamellar vesicles (mean diameter: 60 nm) by sonication for 30 min with 15 min on–off cycles at 40% amplitude using a Vibracell Ultrasonics Disintegrator VCX750 (750 W, 20 kHz; Sonics and Materials, Newtown, Connecticut, USA). Vesicles filled with calcein were prepared by hydrating the dried lipids with 80 mM calcein (Sigma–Aldrich, St Louis, Massachusetts, USA) in KPB. Gel filtration on Sephadex G-50 (Pharmacia Fine Chemicals AB, Uppsala, Sweden) columns was used to remove extravesicular calcein. MLVs loaded with calcein were bath sonicated (Bandelin, Berlin, Germany) at 35 kHz for 15 min in order to obtain unilamellar vesicles (mean diameter: 200 nm) used for a confocal microscope. All the vesicles were sterile filtered.
Cell cultures
All cells were obtained from European collection of Cell Cultures (Salisbury, UK) except EA.hy926 that were purchased from American Type Culture Collection (Manassas, Virginia, USA). They were grown as an attached monolayer culture at 37°C, 5% carbon dioxide (CO2) atmosphere in a humidified chamber.
B16-F1 mouse melanoma cells were grown in Eagle’s minimum essential medium (EMEM) with
Chinese hamster ovary (CHO) cells were grown in Ham’s nutrient mixture F-12 (PAA Laboratories, Pasching, Austria), supplemented with 2 mM
Three human cell lines were also used for studies: epithelial colorectal adenocarcinoma cells (CACO-2), human liver hepatocellular carcinoma cell line (Hep G2) and human endothelial umbilical vein cell line (EA.hy926). EA.hy926 cell line was obtained by the hybridization of primary human umbilical vein endothelial cells with the A549/8 human lung carcinoma cell line but has maintained the phenotype of endothelial cells.
29
CACO-2, Hep G2 and EA.hy926 cells were grown as described before.
30
Briefly, for CACO-2, we used EMEM (Sigma–Aldrich) supplemented with 10% FBS, 1% penicillin/streptomycin, 1%
Cytotoxicity assay
Cytotoxicity was evaluated by the colorimetric 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay 31 (cell growth determination kit (CGD1), MTT based, Sigma–Aldrich), as described by the manufacturer. In brief, cells were seeded to 96-well plates (2000–10,000 cells per well) and allowed 2 h to attach to the surface. Different concentrations of archaeosomes were added to cells (5000–0.000256 µg/ml). After 4 days of incubation at 37°C in a CO2 incubator, the MTT reagent was added to cells and after 4 h purple formazan was extracted from cells by dimethyl sulfoxide and measured spectrophotometrically at 544 nm using a microplate reader (Tecan Infinite M200, Tecan Group Ltd, Männerdorf, Switzerland), and using 690 nm wavelength as a reference.
Statistical analysis was performed using Microsoft Excel (Microsoft Corp., Redmond, Washington, USA). Results from at least three independent experiments were averaged and presented as mean ± SEM. One-way analysis of variance was used to determine statistical differences between control and treated cells and p < 0.05 was considered significant.
Cell labeling and confocal fluorescence microscopy
Cells were seeded to 8-well µ-slides (Ibidi GmbH, Martinsried, Germany), 10,000 cells per well (B16-F1 and CHO), or on round cover glass slides placed in 24-well plates, 20,000 cells per well (CACO-2, Hep G2 and EA.hy926). Cells were then washed and stained by membrane label Vybrant DiI (Vybrant® DiI cell-labeling solution, Molecular Probes/Invitrogen/Life Technologies Corp., Grand Island, New York, USA), following a manufacturer’s protocols (DiI diluted 1:200 in phosphate buffered saline (PBS) for 30 min at 37°C). Cells were then washed three times and used for incubation with archaeosomes. Calcein-encapsulated archaeosomes were added to stained cells at a concentration of 100 µg/1 ml of culture medium and incubated for 10 and 30 min, 1, 2 and 24 h at 37°C in a CO2 incubator. Cells were then washed three times with PBS, fixed with 4% paraformaldehyde and mounted on a glass slide.
B16-F1 and CHO cells were studied by a confocal laser scanning fluorescence microscope (CLSM) Leica TCS SP5 (Leica Microsystems GmbH, Wetzlar, Germany) using 63× water immersion objective and λ EX 488 nm and λ EM 515 nm for calcein and λ EX 561 nm and λ EM 590 nm for Vybrant DiI. CACO-2, Hep G2 and EA.hy926 cells were studied with Nikon confocal microscope system C1si (Nikon Instruments Europe B.V., Kingston, UK), using the same excitation and emission wavelengths.
Results
Cytotoxicity
Cytotoxicity of archaeosomes was tested on five cell lines (Figures 1 to 3): rodent mouse melanoma cells (B16-F1) and CHO cells (Figure 1), as well as three human cell lines—epithelial colorectal adenocarcinoma cells (CACO-2), liver hepatocellular carcinoma cell line (Hep G2; Figure 2) and endothelial umbilical vein cell line (EA.hy926; Figure 3). The archaeosomes were moderately toxic to both rodent cell lines (median lethal dose (LC50) for B16-F1 and CHO cells is around 625 µg/ml) but showed no toxicity to human CACO-2 and Hep G2 cells. However, archaeosomes exhibited strong toxic effect on Ea.hy926 cell line (LC50 around 0.8 µg/ml).
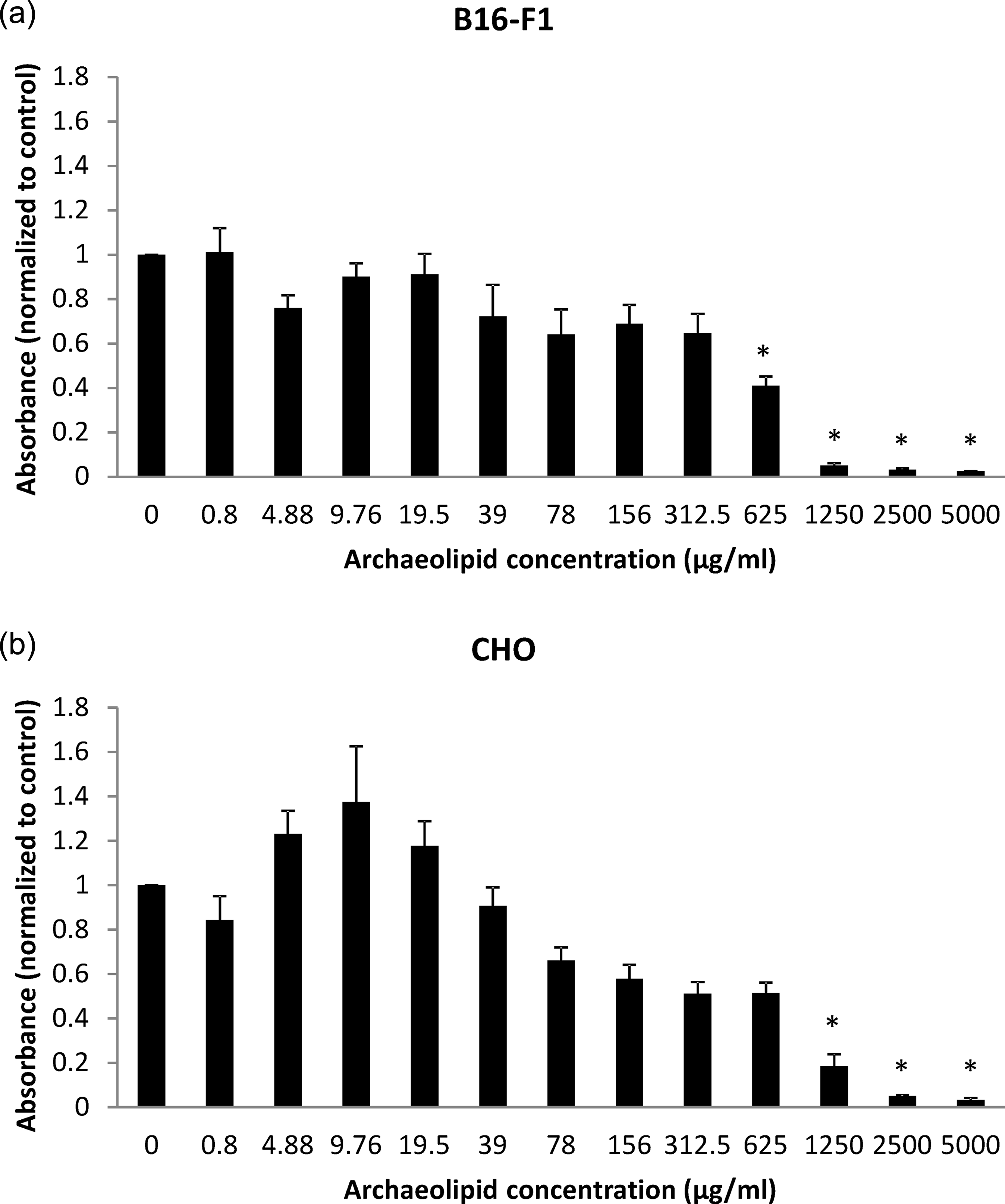
A dose-dependent cytotoxic effect of archaeosomes on rodent cell lines: B16-F1 (a) and CHO (b), measured by MTT cytotoxicity assay. Data are presented as mean ± SEM of at least three independent experiments. Significant differences from controls are designated by asterisks (*p < 0.05).

A dose-dependent cytotoxic effect of archaeosomes on human cell lines: CACO-2 (a) and Hep G2 (b), measured by MTT cytotoxicity assay. Data are presented as mean ± SEM of four independent experiments. Significant differences from controls are designated by asterisks (*p < 0.05).

A dose-dependent cytotoxic effect of archaeosomes on human umbilical vein cell line EA.hy926 measured by MTT cytotoxicity assay. Data are presented as mean ± SEM of four independent experiments. Significant differences from controls are designated by asterisks (*p < 0.05).
Confocal microscopy of archaeosome uptake
The archaeosomes were added to B16-F1, CHO, CACO-2, Hep G2 and EA.hy926 cells. After different incubation times, the cells were fixed and observed under a CLSM (Figure 4). We can see archaeosomes as green dots in the images. The archaeosomes interact with cells in three distinct phases (Table 1). First, they adsorb onto the cell surface ((1) e.g. as in Figure 4, CACO-2, 10 min). Then, they are transferred to the cell interior ((2) e.g. as in Figure 4, CACO-2, 2 h), and finally, they release their load (in our case calcein) into the cytoplasm ((3) e.g. as in Figure 4, EA.hy926, 24 h). Time course and intensity of the uptake of archaeosomes and release of calcein are however different in different cell types (Table 1). In B16-F1, CHO, CACO-2 and Hep G2 cells, the release of calcein into the cytoplasm is observed after 24 h of incubation with archaeosomes, while in EA.hy926 cells this phenomenon can be seen already after 30 min when intensive green coloring of the cytoplasm occurs. In the case of B16-F1, CHO, CACO-2 and Hep G2 cells, intact archaeosomes inside cells persist even after 24 h (Figure 4, CACO-2, 24 h). B16-F1 cells accumulate the least amount of archaeosomes.

Laser scanning confocal microscope images of cell lines CACO-2 and EA.hy926 incubated with archaeosomes loaded with calcein (green). Cells are labeled with Vybrant® DiI cell-labeling solution for membranes (red) prior incubation with archaeosomes. Cell types are shown on left side and incubation times of cells with archaeosomes (10 min, 2 and 24 h) are shown on top. Longitudinal sections are shown.
Interactions between archaeosomes from Aeropyrum pernix lipids, and cells.
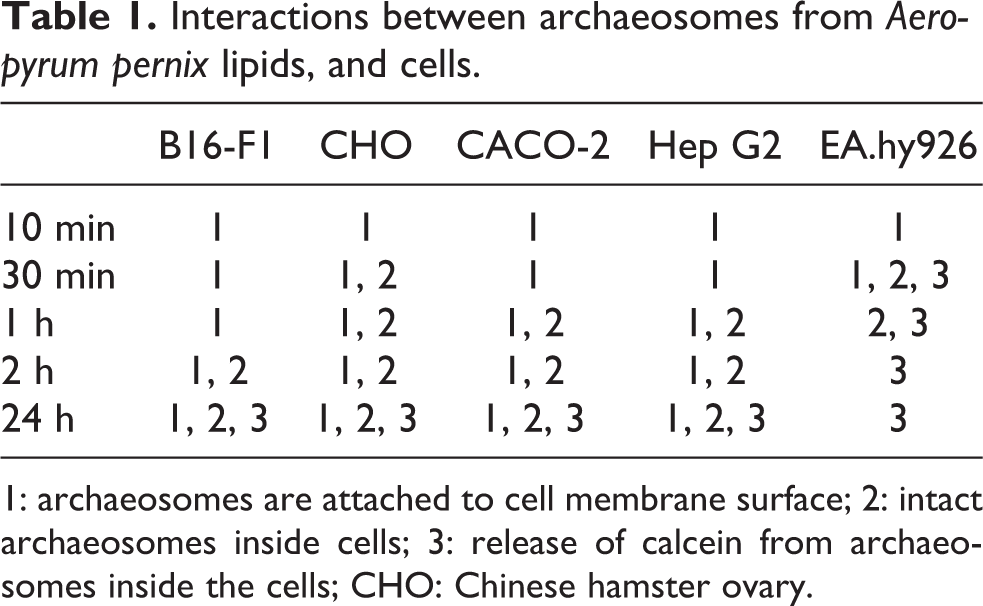
1: archaeosomes are attached to cell membrane surface; 2: intact archaeosomes inside cells; 3: release of calcein from archaeosomes inside the cells; CHO: Chinese hamster ovary.
Discussion
For a possible use of A. pernix archaeosomes as a potential drug carrier system, we studied cytotoxicity and uptake of archaeosomes using rodent and human cell lines. Archaeosomes were loaded with fluorescent calcein. Our results showed no in vitro toxicity of A. pernix archaeosomes to human CACO-2, Hep G2 and mild toxicity to rodent B16-F1 and CHO cells, as tested with the MTT cytotoxicity test. To our knowledge, the presented study is the first report of the interactions of A. pernix archaeosomes and mammalian cells in vitro. This is in line with the previous toxicity studies of liposomes from different archaeal species that have been found to be nontoxic or mildly toxic in vitro and in vivo. 24 –27 Rather surprisingly, our archaeosomes showed strong cytotoxic effect on human endothelial umbilical vein cell line EA.hy926.
Although most of the liposomes are inert and lack toxicity toward cells, toxicity studies of liposomes on various cell lines have been reported in the literature. Filion and Phillips 32 found that liposomes composed of cationic lipids and fusogenic dioleoylphosphatidylethanolmine are highly toxic in vitro toward macrophages. This was in line with the observation that higher amounts of lipids were associated with the cells of macrophage origin than the hepatocytes. 33 On the other hand, fibroblasts were sensitive toward two different phosphatidylcholine liposomes. 34
Our results show that archaeosomes composed of lipids from archaeon A. pernix are taken up by all tested cells: rodent mouse melanoma cells (B16-F1 and CHO cells, and human epithelial colorectal adenocarcinoma cells (CACO-2), liver hepatocellular carcinoma cell line (Hep G2) and endothelial umbilical vein cell line (EA.hy926). Total polar lipids of A. pernix are composed solely of two lipids, AI and AGI, and both of them have inositol in their head group. 19 Similarly, Tolson et al. found that liposomes from archaeon Methanosarcina mazei that contained a high content of phosphatidylinositol interacted strongly with macrophages. 24 The mechanism of the uptake is still to be elucidated. We can clearly see intact archaeosomes inside the cells so fusion is unlikely to be the uptake mechanism, therefore one of the endocytotic pathways is most likely responsible for the uptake of archaeosomes.
Numerous in vitro studies have already revealed that liposomes can be internalized by cells with endocytosis. 35 –37 After binding to the cell surface, the liposomes are internalized by phagocytosis in phagocytic cells, and in nonphagocytic cells the main endocytotic pathway is clathrin-mediated or caveolae-mediated endocytosis, although liposomes can enter the cells also by macropinocytosis or other pathways (reviewed in the literature 38 ). Charged liposomes (either positively or negatively charged) are endocytosed faster and to a greater extent than neutral liposomes by endocytotic cells, although different cells have different liposome-binding sites on their surface. 39,40
Phagocytic uptake of liposomes prepared from ether lipids extracted from various archaeal species was several times greater than the uptake of liposomes composed of conventional lipids (dipalmitoylphosphatidylcholine, dimyristoylphosphatidylcholine: dimyristoylphosphatidylglycerol:cholesterol). It has also been shown that interactions of archaeosomes with the nonphagocytic cells were significantly lower than in macrophages. 24 Results of Sprott and his colleagues suggest endocytosis of archaeosomes, rich in surface-exposed phosphoserine head groups via a phosphatidylserine receptor. Archaeosomes composed of certain other archaeal lipids (lacking phosphoserine head groups) are suggested to enter the cells after energy-independent surface adsorption. 41,42
In EA.hy926 cells, we can see calcein released in the cytoplasm shortly after adding archaeosomes loaded with calcein to the medium. A fast release of large amounts of calcein from archaeosomes can be related to higher rate of archaeosome uptake and release in these cells due to fast endocytosis. This is in line with high cytotoxicity of archaeosomes of A. pernix toward EA.hy926 cells. Anionic liposomes bind to different receptors at the plasma membrane. 43 –45 EA.hy926 cells are human macrovascular endothelial cell lines 29 that are used as an in vitro experimental model for studying vascular functions such as adhesion 46 and angiogenesis. 47 It expresses numerous surface molecules characteristic of human vascular endothelium (reviewed in the literature 48 ). These specific surface molecules might be responsible for more prominent uptake of archaeosomes in EA.hy926 cells. Once inside the EA.hy926 cells, archaeal lipids could fuse with cell membranes and contribute to more rigid and stable state of membranes or affect other cell mechanisms that lead to cell death. In other cells, more archaeosomes stay intact in endosomes and do not release calcein to the same extent as in EA.hy926 cells. The role of specific properties of EA.hy926 cells in archaeosomal uptake need to be further explored to elucidate the observed cytotoxicity. Cell surface–receptor binding, competitive inhibition assays and studies of kinetics of endocytosis have to be carried out in the future. Intact archaeosomes that persist in cells could be electroporated with the use of nanosecond electric pulses that affect internal cell membranes to release their cargo. 49,50 In this way, a selective release of archaeosome content could be achieved.
Archaeosomes from A. pernix lipids are potential drug carriers that can resist extreme environmental conditions such as high temperatures and acidic or basic pH, 21 although it is not yet known how the A. pernix archaeosomes act in vivo. Negatively charged A. pernix archaeosomes could be prone to rapid opsonization and uptake by the reticuloendothelial system as other anionic liposomes. 51 By adding different molecules to the surface of archaeosomes, their blood circulation time could however be prolonged 52 and they could be prepared to be used for targeted drug delivery. 53
To conclude, our in vitro study shows that archaeosomes composed of lipids from archaeon A. pernix are taken up by a variety of cells: B16-F1, CHO, CACO-2, Hep G2 and EA.hy926 cells. Since A. pernix archaeosomes are very stable in extreme environments (pH range from 4 to 12 and the temperature range from 0 to 100°C) and capable of encapsulating biologically active compounds, they are potential drug delivery system. They have proved to be nontoxic to CACO-2 and Hep G2 cells and only mildly toxic to B1-6F1 and CHO cells. However, they are very toxic to EA.hy926 endothelial umbilical vein cell line, which is related most probably to a fast endocytosis and/or intracellular release and action in this type of cells. The mechanisms of uptake, release and action in these cells as well as in vivo functioning archaeosomes have to be further evaluated for possible targeted drug delivery.
Footnotes
Acknowledgments
The authors would like to thank Dr Mojca Benčina for her invaluable help with confocal microscopy.
Funding
The study was supported by the Slovenian Research Agency (ARRS; Grant numbers J2-3639, P2-0249, P4-0121 and MR-1000-06-310166). Research was conducted in the scope of the EBAM European Associated Laboratory (LEA).