
Abstract
Serrapeptase (SP) and nattokinase (NK) are proteolytic enzymes belonging to serine proteases. In this study, we hypothesized that SP and NK could modulate certain factors that are associated with Alzheimer’s disease (AD) pathophysiology in the experimental model. Oral administration of aluminium chloride (AlCl3) in a dose of 17 mg/kg body weight (bw) daily for 45 days induced AD-like pathology in male rats with a significant increase in brain acetylcholinesterase (AchE) activity, transforming growth factor β (TGF-β), Fas and interleukin-6 (IL-6) levels. Meanwhile, AlCl3 supplementation produced significant decrease in brain-derived neurotrophic factor (BDNF) and insulin-like growth factor-1 (IGF-1) when compared with control values. Also, AlCl3 administration caused significant decline in the expression levels of disintegrin and metalloproteinase domain 9 (ADAM9) and a disintegrin and metalloproteinase domain 10 (ADAM10) genes in the brain. Histological investigation of brain tissue of rat model of AD showed neuronal degeneration in the hippocampus and focal hyalinosis with cellular as well as a cellular amyloid plaques formation. Oral administration of SP or NK in a rat model of AD daily for 45 days resulted in a significant decrease in brain AchE activity, TGF-β, Fas and IL-6 levels. Also, the treatment with these enzymes produced significant increase in BDNF and IGF-1 levels when compared with the untreated AD-induced rats. Moreover, both SP and NK could markedly increase the expression levels of ADAM9 and ADAM10 genes in the brain tissue of the treated rats. These findings were well confirmed by the histological examination of the brain tissue of the treated rats. The present results support our hypothesis that the oral administration of proteolytitc enzymes, SP and/or NK, would have an effective role in modulating certain factors characterizing AD. Thus, these enzymes may have a therapeutic application in the treatment of AD.
Introduction
In humans, Alzheimer’s disease (AD) develops over many years and it comprises a chain of subtle alterations in brain function, finally leading to the impairment of memory and cognition. 1 The cognitive disability includes the impairment of behaviour, visual-spatial skills, speech and motor ability, depression, delusions, hallucinations, aggressive behaviour and ultimately, increasing dependence upon others before death. 2 Genetic and neuropathological evidence suggests that the accumulation of amyloid-β (Aβ) peptides in the brain is a key event in the pathophysiology of AD. Amyloid fibrils are an ordered protein aggregate with a lamellar cross-beta structure. Enhancing amyloid clearance is one of the targets of the therapy of AD. 3 Therefore, the therapeutic potential of several approaches aimed at lowering the level of Aβ in the brain is currently being investigated. 4
Proteases are ubiquitous enzymes that are crucial for cell growth, development, apoptosis, protein turnover and cell cycle regulation during life process. 5 Several serine proteases have been well characterized in the brain. Accumulating evidence demonstrates that some serine proteases play very important roles in the neural development, plasticity and neuroregeneration in the brain. 6 The basis of these functions is the unique correct three-dimensional structures of the proteins, and all the information necessary for proteolysis are encoded by the amino acid sequence of protease. 7 Diseases such as cystic and even some forms of cancer are believed to result due to defective protein folding. 8
Serrapeptase (SP) is a proteolytic enzyme belonging to serine proteases. It is derived from the non-pathogenic enterobacteria Serratia E15. It is produced in the intestines of silkworms to break down cocoon walls. It has an anti-inflammatory effect. 9 This enzyme is believed to induce the degradation of insoluble protein products like fibrin, biofilm and inflammatory mediators. 10 Therefore, SP has also been used for the treatment of a number of inflammatory conditions. 11,12
Nattokinase (NK), a bacterial serine protease derived from Bacillus subtilis, has been reported to have potent fibrinolytic activity. 13 It is considered to be one of the most active functional ingredients found in natto, a traditional Japanese food prepared from the fermented soybeans. 14 This enzyme is reported to directly digest fibrin especially in its cross-linked form. 15 It has a fourfold greater thrombus-dissolving activity than plasmin and is shown to potentiate endogenous fibrinolysis by cleavage and to inactivate plasminogen activator inhibitor 1 (PAI-1), 16 leading to efficient lysis of the detrimental coagulation of blood in the body. 15 NK not only dissolves blood clots 17 but also degrades amyloid fibrils. 3
In this study, we hypothesized that SP and NK could correct certain factors including proteins, growth factors, apoptotic factor, inflammatory factor and genes that are associated with AD pathophysiology in the experimental model. To test this hypothesis, we investigated the effects of SP and NK on brain cholinesterase activity, brain-derived neurotrophic factor (BDNF), transforming growth factor β (TGF-β), insulin-like growth factor-1 (IGF-1), Fas and interleukin-6 (IL-6) levels. The study was extended to examine the effects of these enzymes on the expression levels of disintegrin and metalloproteinase domain 9 (ADAM9) and disintegrin and metalloproteinase domain 10 (ADAM10) genes. Histological investigation of brain tissue was also carried out to confirm the biochemical and molecular genetics observations.
Materials and methods
Aluminium chloride (AlCl3) was purchased from Sigma Co. (California, USA). Its molecular weight is 133.34.
The two proteolytic enzymes, SP (Doctors Best Serrapeptase 40,000 serratio units per veggie cap.) and NK (Doctors BEST Nattokinase 2000 FUs per veggie cap) were purchased from a dietary supplement market in USA.
Reagents for real time-polymerase chain reaction (RT-PCR) were purchased from Invitrogen (Carlsbad, USA) and Fermentas (Leon-Rot, Germany).
The human doses of the two proteolytic enzymes used were converted into rat equivalents with the help of conversion table described by Paget and Barnes. 18
A total of 66 adult male albino rats weighing 120–150 g were enrolled in the current study. The rats were obtained from the Animal House Colony of the National Research Centre, Cairo, Egypt. The animals were housed in polypropylene cages in an environmentally controlled clean air room with a temperature of 24 ± 1°C, an alternating 12 h light–dark cycle, a relative humidity of 60 ± 5% and free access to tap water and food. Rats were allowed to adapt to these conditions for 2 weeks before beginning the experimental protocol. The animal experimental protocol has been approved by the Institutional Animal Ethics Committee of Medical Research, National Research Centre, Egypt. AD-like pathology model was achieved in rats by the oral administration of AlCl3 in a dose of 17 mg/kg bw 19 daily for 45 days. The animals used in the present study were classified into six groups; G1: healthy control group (negative control); G2: AD group (positive control); G3: AD group treated orally with water suspension of SP in a dose of 10.8 U/kg bw daily for 45 days; G4: AD group treated orally with water suspension of SP in a dose of 21.6 U/kg bw daily for 45 days; G5: AD group treated orally with the water suspension of NK in a dose of 360 FU/kg bw daily for 45 days and G6: AD group treated orally with water suspension of NK in a dose of 720 FU/kg bw for 45 days daily.
At the end of the experimental period, the animals were fasted overnight, sacrificed by decapitation under diethylether anaesthesia. The whole brain of each rat was rapidly and carefully dissected, thoroughly washed with isotonic saline, dried on a filter paper and weighed. The whole brain of a certain number of animals in each group was fixed in formalin buffer (10%) for 24 h for histological examination and the whole brain of the rest number of animals in each group was mid-sagitally divided into two portions; the first portion was immediately stored in liquid nitrogen for gene expression analysis, while the second portion of the brain was weighed and homogenized immediately to give 10% (w/v) homogenate in ice-cold medium containing 50 mM Tris-HCl (pH 7.4) and 300 mM sucrose. 20 The homogenate was centrifuged at 1800g for 10 min in a cooling centrifuge at 4°C. The supernatant (10%) was stored at −80°C till further analysis to assess cholinesterase activity, BDNF, TGF-β, IGF-1, Fas as well as IL-6 levels.
Biochemical analyses
Brain cholinesterase activity was determined kinetically using a kit purchased from Biostc Co (Cairo, Egypt), according to the method of Young. 21 While, brain BDNF level was assayed by enzyme linked immunosorbent assay (ELISA) technique using kit purchased from RayBiotech, Inc. (Norcross, USA), according to the method described by Barde. 22 Brain TGF-β and IGF-1 levels were determined by ELISA technique using a kit purchased from DRG instrument GmbH (Marburg, Germany), according to the methods described by Kropf et al. 23 and Lewitt et al. 24 respectively. Meanwhile, brain Fas level was detected by the ELISA technique using kit purchased from Diaclone a Tepnel Co. (France), according to the method described by Iio et al. 25 Finally, brain IL-6 level was assayed by ELISA technique using kit purchased from RayBiotech, Inc, according to the method of Bauer and Herrmann. 26 Quantitative estimation of total protein content in the brain was carried out according to the method of Lowry et al. 27 to express the concentration of different brain parameter per milligram protein. 28
Gene expression analysis
RNA extraction
Brain tissues of rats in each group were used to extract the total RNA. Total RNA was isolated from 100 mg of tissue by the standard TRIzol extraction method (Invitrogen, USA) and recovered in 100 μl molecular biology grade water. In order to remove any possible genomic DNA contamination, the total RNA samples were treated by DNA-free™ DNase and removal reagents kit (Ambion, Austin, Texas, USA) following the manufacturer’s protocol. The RNA concentration was determined by spectrophotometric absorption at 260 nm.
First-strand cDNA synthesis
To synthesize the first-strand complementary DNA (cDNA), 5 µg of the complete poly(A)+ RNA isolated from rat brain tissue was reverse transcribed into cDNA in a total volume of 20 µl using 1 µl oligo (poly(deoxythymidine)18) primer. 29 The composition of the reaction mixture was 50 mM MgCl2, 10X reverse transcription (RT) buffer (50 mM KCl; 10 mM Tris-HCl; PH 8.3), 200 U/µl reverse transcriptase (RNase H free), 10 mM of each dNTP and 50 µM of oligo (dT) primer. The mixture of each sample was centrifuged for 30 s at 1000g and transferred into the thermocycler (Biometra GmbH, Göttingen, Germany). The RT reaction was carried out at 25°C for 10 min, followed by 1 h at 42°C, and finished with denaturation step at 99°C for 5 min. Later, the reaction tubes containing RT preparations were flash-cooled in an ice chamber until being used for DNA amplification through RT-PCR. 30
RT-PCR assay
The first strand cDNA from each brain sample was used as a template for the semi-quantitative RT-PCR with a pair of specific primer in a 25-µl reaction volume. The sequences of specific primer and product sizes are listed in Table 1. β-Actin was used as a housekeeping gene for normalizing messenger RNA levels of the target genes. The reaction mixture for RT-PCR consisted of 10 mM dNTP’s, 50 mM MgCl2, 10X PCR buffer (50 mM KCl; 20 mM Tris-HCl; pH 8.3), 1 U/µl Taq polymerase and autoclaved water. The PCR-cycling parameters of the studied genes (ADAM9, ADAM10 and β-actin) were performed as the PCR condition summarized in Table 1. The PCR products were then loaded onto 2.0% agarose gel, along with the PCR products derived from β-actin of the different rat samples. Each reaction of the RT-PCR was repeated with the samples obtained from eight rats, generating new cDNA products at least eight times per each group.
Primers and PCR-thermocycling parameters

PCR: polymerase chain reaction; ADAM9: disintegrin and metalloproteinase domain 9; ADAM10: disintegrin and metalloproteinase domain 10.
Histological investigation
Brain tissues previously fixed in formalin buffer (10%) for 24 h were washed under tap water for 20 min. Then, the serial dilutions of alcohol (methyl, ethyl and absolute ethyl) were used for dehydration. Specimens were cleared in xylene and embedded in paraffin at 56°C in hot air oven for 24 h. Paraffin beeswax tissue blocks were prepared for sectioning at 4-µm-thick using slidge microtome. The obtained tissue sections were collected on glass slides, deparaffinized and stained by hematoxylin and eosin stains 31 for histological examination through the light microscope.
Statistical analysis
The results were expressed as mean ± SE. Data were analyzed by one-way analysis of variance and were performed using the Statistical Package for the Social Science (SPSS) program, version 11 followed by least significant difference to compare the significance between groups. 32 Difference was considered significant at p < 0.05.
Results
Biochemical results
The data in Table 2 showed the effect of treatment with SP and NK on brain cholinesterase activity in the model of AD. The untreated AD group rats showed significant increase in brain cholinesterase activity in comparison with the negative control group. In contrast, the groups of AD group treated with low or high dose of SP (10.800 or 21.600 U/kg bw) or NK (360 or 720 Fu/kg bw) showed significant decrease in brain cholinesterase activity in comparison with the untreated AD group.
Effect of treatment with SP and NK on brain cholinesterase activity in the model of ADa

NK: nattokinase; SP: serrapeptase; AD: Alzheimer’s disease.
aSignificant change at p < 0.05 in comparison with the negative control group.
bSignificant change at p < 0.05 in comparison with the AD group.
Table 3 showed the effect of treatment with SP or NK on brain BDNF, TGF-β and IGF-1 levels in AD group. The untreated AD group showed significant decrease in brain BDNF and IGF-1 levels in concomitant with significant increase in brain TGF-β level in comparison with the negative control group. On the other hand, treatment with low or high dose of SP or NK caused significant increase in brain BDNF and IGF-1 levels associated with a significant decrease in brain TGF-β level in comparison with the untreated AD group.
Effect of treatment with SP and NK on brain BDNF, TGF-β and IGF-1 levels in the model of ADa
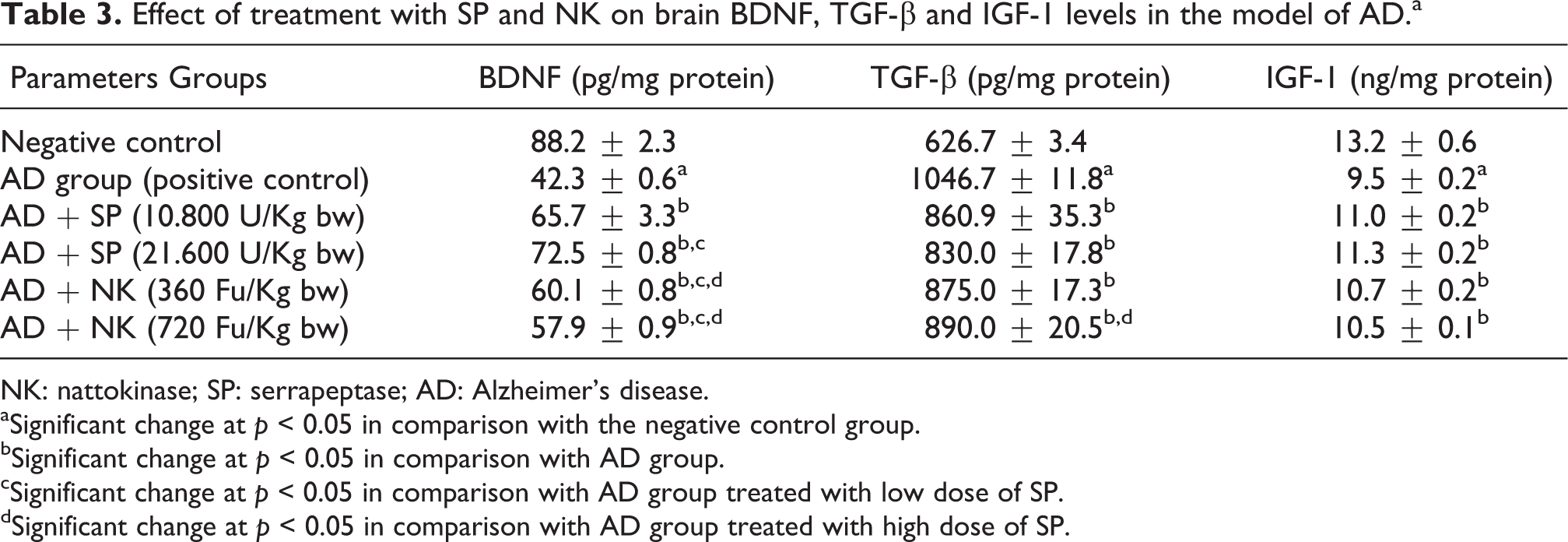
NK: nattokinase; SP: serrapeptase; AD: Alzheimer’s disease.
aSignificant change at p < 0.05 in comparison with the negative control group.
bSignificant change at p < 0.05 in comparison with AD group.
cSignificant change at p < 0.05 in comparison with AD group treated with low dose of SP.
dSignificant change at p < 0.05 in comparison with AD group treated with high dose of SP.
Noteworthy, treatment with a high dose of SP caused significant increase in brain BDNF level in comparison with the group of rats treated with low dose of SP. On the other hand, treatment with low or high dose of NK produced significant decrease in brain BDNF level in comparison with low or high dose of SP. Meanwhile, the group of rats treated with high dose of NK showed significant increase in brain TGF-β level in comparison with the group of rats treated with high dose of SP.
The data in Table 4 showed the effect of treatment with SP and NK on brain Fas and IL-6 levels in the model of AD. The untreated AD group showed significant increase in brain Fas and IL-6 levels in comparison with the negative control group. On the contrary, treatment with low or high dose of SP or NK caused significant decrease in brain Fas level in comparison with the untreated AD group. Also, treatment with high dose of SP produced significant decrease in brain IL-6 level in comparison with the untreated AD group. Noteworthy, treatment with low dose of NK showed significant elevation in brain Fas level when compared with the group of rats treated with high dose of SP. Meanwhile, treatment with high dose of NK caused significant increase in brain Fas level when compared with the groups of AD treated with low or high dose of SP as well as low dose of NK.
Effect of treatment with SP and NK on brain Fas and IL-6 levels in the model of ADa
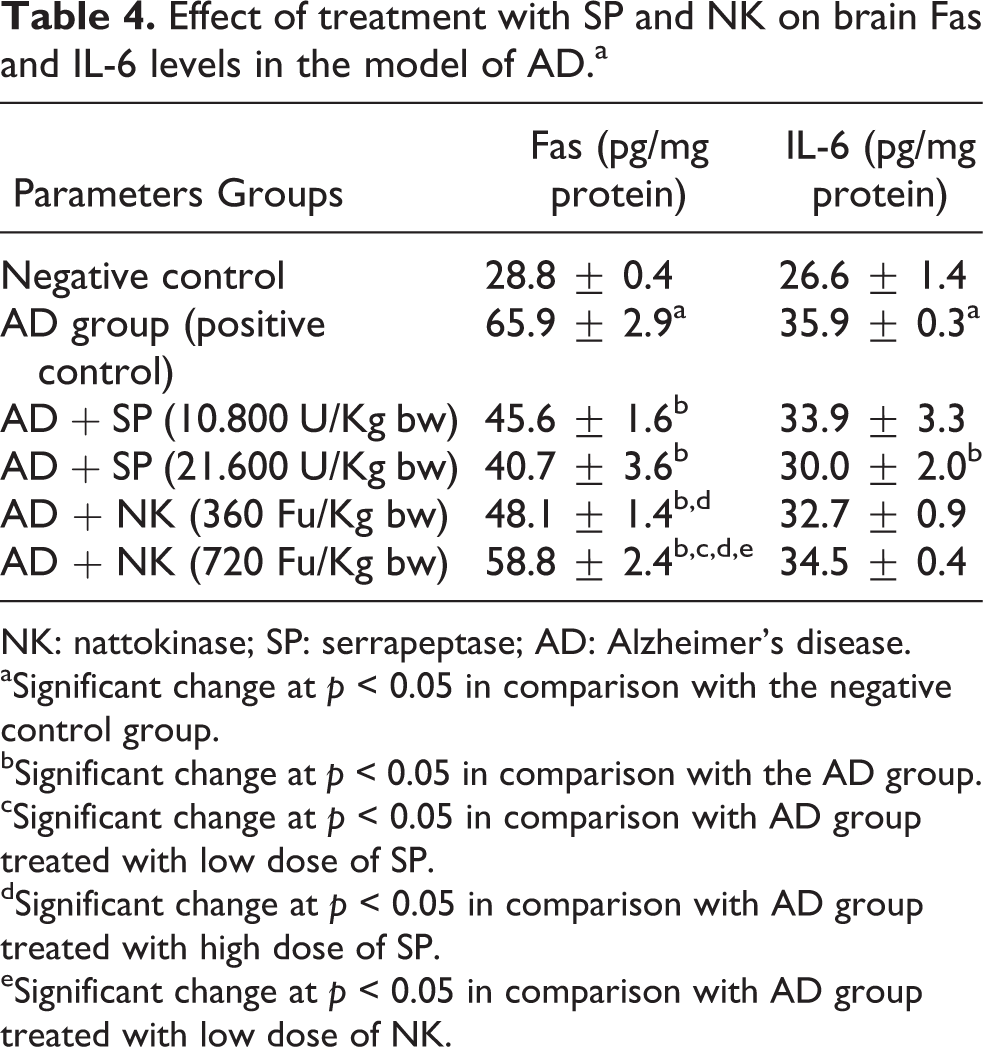
NK: nattokinase; SP: serrapeptase; AD: Alzheimer’s disease.
aSignificant change at p < 0.05 in comparison with the negative control group.
bSignificant change at p < 0.05 in comparison with the AD group.
cSignificant change at p < 0.05 in comparison with AD group treated with low dose of SP.
dSignificant change at p < 0.05 in comparison with AD group treated with high dose of SP.
eSignificant change at p < 0.05 in comparison with AD group treated with low dose of NK.
Molecular genetic results
Expression of ADAM9 and ADAM10 genes
The results of the expression analysis of ADAM9 and ADAM10 genes in different studied groups are represented in Figures 1 and 2.
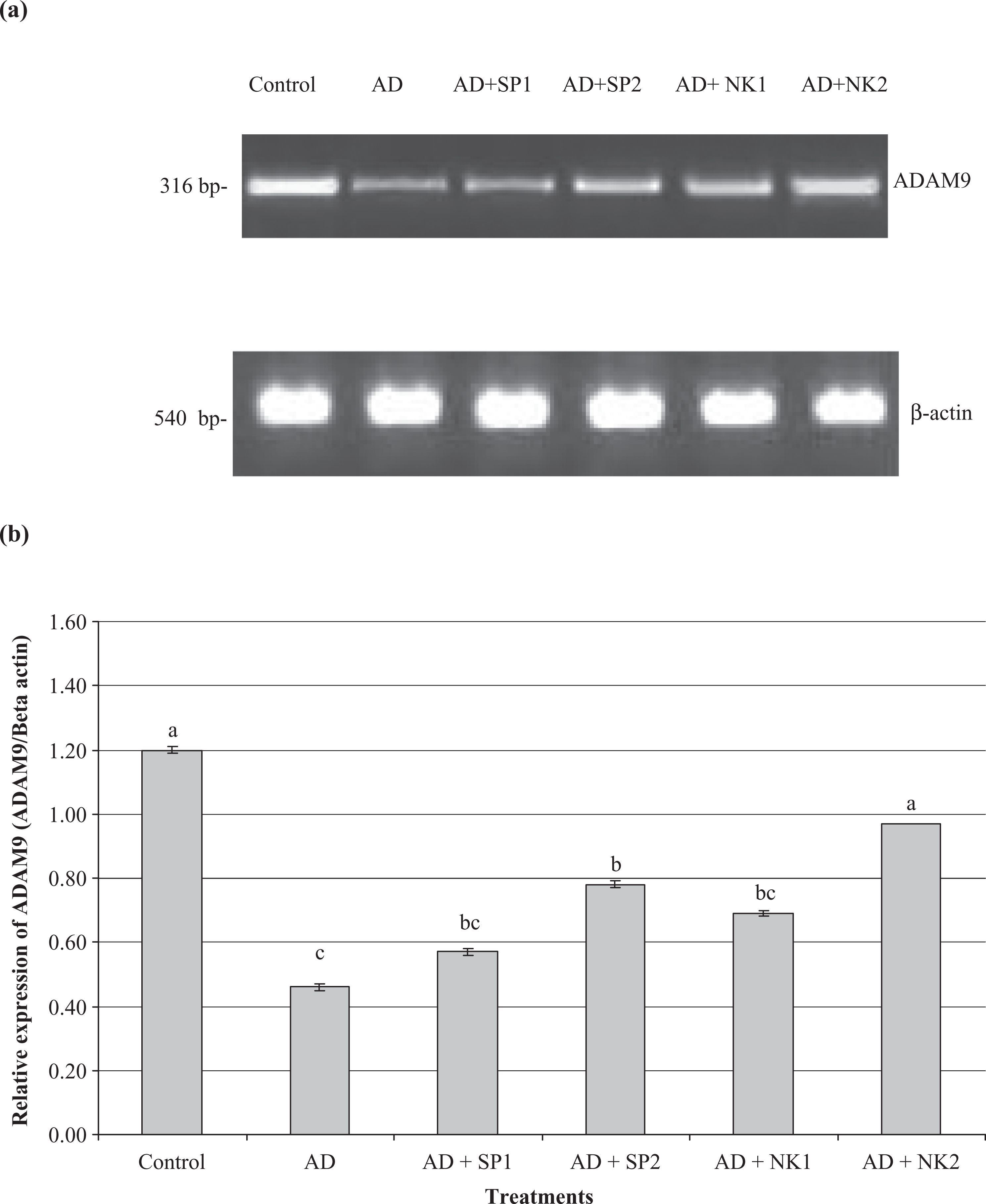
Semiquantitative RT-PCR of ADAM9 gene in brain tissue of AD group treated with SP and NK at low (a) and high (b) doses for 45 days. Superscripts with different letters within each column means significantly different (p < 0.05). RT-PCR: real time-polymerase chain reaction; ADAM9: disintegrin and metalloproteinase domain 9; SP: serrapeptase; NK: nattokinase; ADAM10: disintegrin and metalloproteinase domain 10; AD: Alzheimer’s disease.

Semi-quantitative RT-PCR of ADAM10 gene in brain tissue of AD group treated with SP and NK at low (a) and high (b) doses for 45 days. Superscripts with different letters within each column means significantly different (p < 0.05). RT-PCR: real time-polymerase chain reaction; SP: serrapeptase; NK: nattokinase; ADAM10: disintegrin and metalloproteinase domain 10; AD: Alzheimer’s disease.
The current results revealed that the expression level of ADAM9 gene was significantly decreased in the brain tissues of AD group in comparison with the negative control group (Figure 1). In comparison with the expression level of ADAM9 in the brain tissue of AD group, low doses of SP and NK did not affect the expression level of ADAM9 gene in brain tissue of the treated AD group significantly, while the expression level of ADAM9 gene in the brain tissue of these groups was still higher than that in the untreated AD group. Using high doses of SP or NK increased the expression level of ADAM9 in the brain tissue of the treated AD group significantly when compared with that of the untreated AD group. Significant increase in the expression level of ADAM9 gene in the brain tissue of AD group treated with high dose of NK when compared with that in AD group treated with the low dose of NK as well as low or high dose of SP (Figure 1).
Regarding the expression level of ADAM10 gene in the brain tissue, the results of the present study revealed that the expression level of ADAM10 gene was significantly decreased in AD group when compared with the negative control group (Figure 2). Low dose of SP did not affect the expression level of ADAM10 gene in the brain tissue of the treated AD group significantly when compared with that in the untreated AD group, although the expression level of ADAM10 gene in the brain tissue of this group was still higher than that in the untreated AD group (Figure 2). However, the treatment of AD group with the low dose of NK was able to increase the expression level of ADAM10 gene in the brain tissue significantly when compared with that in the untreated AD group. On the other hand, using a high dose of SP or NK increased the expression level of ADAM10 in the brain tissue of the treated AD group significantly when compared with that in the untreated AD group. Similarly, treatment of AD group with low dose of SP produced significant decrease in the expression level of ADAM10 gene in the brain tissue when compared with that in AD group treated with high dose of NK, low dose of NK as well as high dose of NK (Figure 2).
Histological results
Our histological study showed that there is no histopathological alteration observed in the meninges, cerebral cortex and cerebrum (Figure 3(a)), hippocampus (Figure3(b)), cerebellum (Figure 3(c)) and medulla oblongata (Figure 3(d)) of the brain of healthy control rats (negative control group).

(a) Photomicrograph of brain section of rat in the healthy control (negative control) group showing the normal histological structure of the meninges (m) cerebral cortex (CC) and cerebrum (C). (H&E, ×40). (b) Photomicrograph of brain section of rat in the healthy control (negative control) group showing the normal histological structure of the hippocampus (hp). (H&E, ×40). (c) Photomicrograph of brain section of rat in the healthy control (negative control) group showing the normal histological structure of the cerebellum (Cr) (H&E, ×40). (d) Photomicrograph of brain section of rat in the healthy control (negative control) group showing the normal histological structure of the medulla oblongata (mo). (H&E, ×40). H&E: hematoxylin and eosin stain.
Oral administration of AlCl3 in a dose of 17 mg/kg bw for 45 days induced AD-like pathology model (AD group). This event is confirmed by our histological investigation of brain tissues of rats in this group (Figures 4(a) to (g)). These figures showed the appearance of focal gliosis in the cerebral cortex (Figure 4(a)) associated with neuronal degeneration in the hippocampus (Figure 4(b)). The hippocampus also showed focal hyalinosis with cellular (Figure 4(c)) as well as a cellular amyloid plaques formation (Figure 4(d)). Encephalomlacia was detected in the cerebral matrix with the degeneration of the surrounding neurons (Figure 4(e)), while the blood vessels showed perivascular cuffing (Figure 4(f)). Also, there was degeneration in the Purkenje cells of the cerebellum (Figure 4(g)).
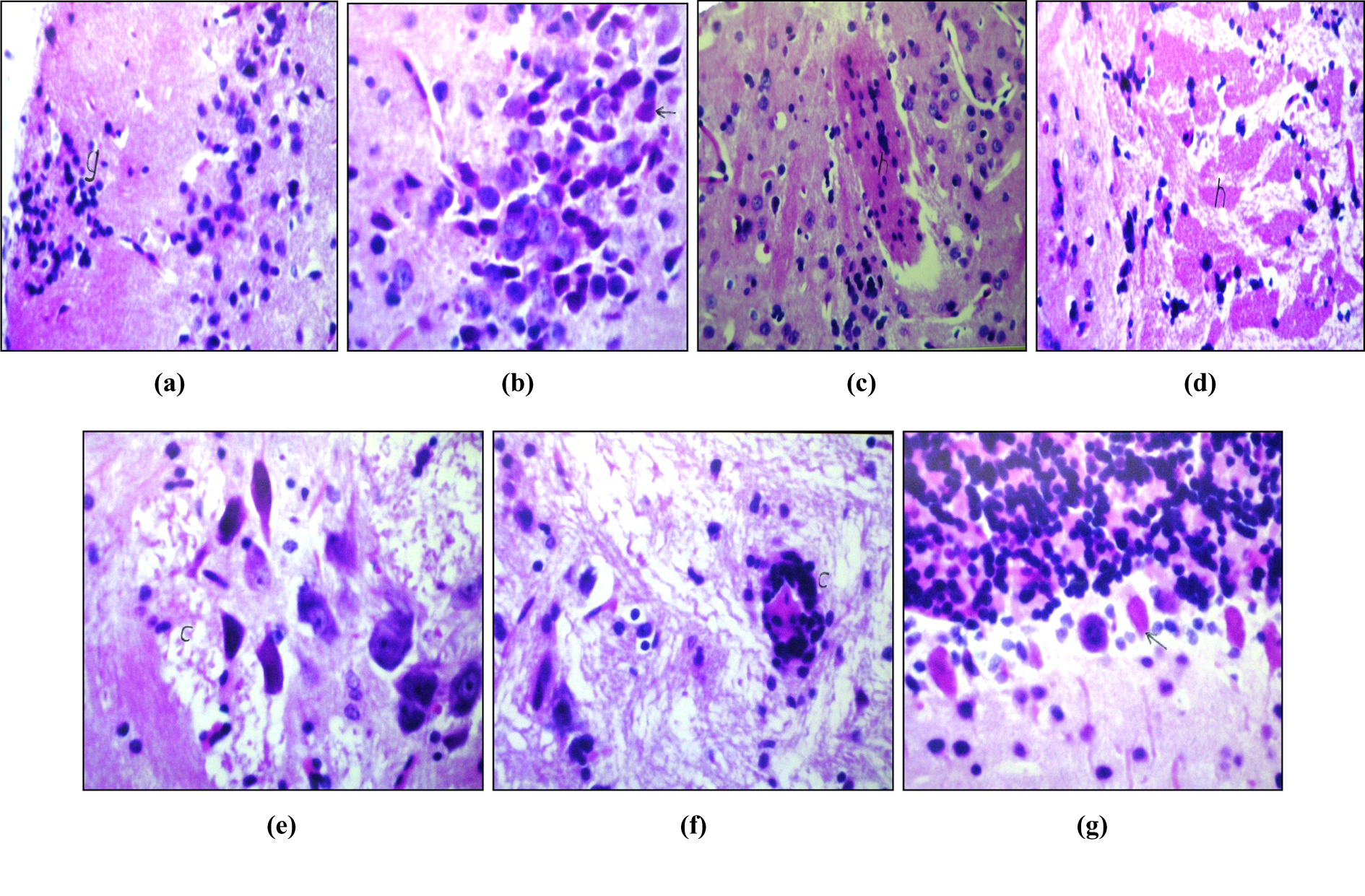
(a) Photomicrograph of brain section of rat (AD group) showing focal gliosis in cerebral cortex (g) (H&E, ×64). (b) Photomicrograph of brain section of rat (AD group) showing neuronal degeneration in the hippocampus (arrow) (H&E, ×80). (c) Photomicrograph of brain section of rat (AD group) showing focal hyalenosis with cellular Plaque formation (h) in hippocampus (H&E, ×80). (d) Photomicrograph of brain section of rat (AD group) showing hyalenosis with a cellular plaque formation and encephalomalacia (h) (H&E, ×64). (e) Photomicrograph of brain section of rat (AD group) showing encephalomalacia and neuronal degeneration (C) (H&E x 160). (f) Photomicrograph of brain section of rat (AD group) showing pesivascular caffing (C) (H&E ×160). (g) Photomicrograph of brain section of rat (AD group) showing degeneration in purkenji cells (arrow) in cerebellum (H&E, ×160). AD: Alzheimer’s disease; H&E: hematoxylin and eosin stain.
Treatment of AD group rats with the low dose of SP (10.8 U/kg bw) for 45 days resulted in an appreciable improvement in the histological feature of the different brain areas in concomitant with the disappearance of amyloid plaques in the hippocampus. There was only focal gliosis in the cerebrum (Figure 5(a)). Treatment of AD group with the high dose of SP (21.6 U/kg bw) led to a complete dissolution of amyloid plaques in the hippocampus. Only, the presence of focal gliosis (Figure 5(b)) and perivascular cuffing (Figure 5(c)) was observed in the cerebrum.

(a) Photomicrograph of brain section of rat in AD group treated with low dose of SP (10.8 U/kg bw) daily for 45 days showing focal gliosis in cerebrum (g) (H&E, ×40). (b) Photomicrograph of brain section of rat in AD group treated with high dose of SP (21.6 U/kg bw) daily for 45 days showing focal gliosis in cerebrum (g) (H&E, ×40). (c) Photomicrograph of brain section of rat in AD group treated with high dose of SP (21.6 U/kg bw) daily for 45 days showing pesivascular caffing (arrow) (H&E, ×40). AD: Alzheimer’s disease; H&E: hematoxylin and eosin stain; SP: serrapeptase.
Treatment of AD group with a low dose of NK (360 Fu/ kg bw) for 45 days revealed an obvious improvement in the histology of different brain areas associated with the lysis of amyloid plaques in the hippocampus. Focal gliosis was observed only in the striatum of the cerebrum (Figure 6(a)). Treatment of AD group with a high dose of NK (720 Fu/kg bw) resulted in the disappearance of amyloid plaques in the hippocampus. The cerebrum showed focal gliosis (Figure 6(b)) as well as diffuse gliosis in the striatum of the cerebrum (Figure 6(c)).

(a) Photomicrograph of brain section of rat in AD group treated with low dose of NK (360 Fu/kg bw) daily for 45 days showing focal gliosis in striatum of cerebrum (g) (H&E, ×40). (b) Photomicrograph of brain section of rat in AD group treated with high dose of NK (720 Fu/kg bw) daily for 45 days showing focal gliosis in cerebrum (g) (H&E, ×40). (c) Photomicrograph of brain section of rat in AD group treated with high dose of NK (720 Fu/kg bw) daily for 45 days showing diffuse gliosis in striatum of cerebrum (arrow) (H&E, ×64). AD: Alzheimer’s disease; H&E: hematoxylin and eosin stain; NK: nattokinase.
Discussion
Several proteins can form amyloid fibrils in vivo, which are related to various diseases, such as AD. Enhancing amyloid clearance is one of the targets of the therapy of these amyloid-related diseases. 3 In this study, we investigate whether the proteolytic enzymes, namely SP and NK, could ameliorate certain factors involved in AD pathophysiology in an experimental rat model in an attempt to explore new modalities for the management of AD.
Protoelytic enzymes are a group of enzymes that break the long chain-like molecules of proteins into shorter fragments (peptides) and eventually into their components called amino acids. 33
The current study demonstrated that the AlCl3 administration (AD-like pathology model) revealed a significant increase in brain acetylcholinesterase (AchE) activity. This result coincides with that of Zhang et al. 34 The two suggested mechanisms for aluminium-induced stimulation of AchE activity in the brain are: (1) aluminium can induce caspase-1 activation and IL-1β secretion. 35 IL-1β has been found to promote the activity and expression of AchE both in cell cultures and in vivo. 36 (2) aluminium can augment the accumulation of insoluble Aβ protein. 37 Aβ induced elevation in the AchE activity through the induction of lipid peroxidation in neuronal membranes due to the production of hydrogen peroxide (H2O2). 38 H2O2 may have a direct action on the AchE enzyme as it acts as a modulator (may be allosteric) in the activity of functionally important proteins, receptors and enzymes. 39 Based on these findings, aluminium’s action on AchE activity may be related to the involvement of aluminium in the aetiology of AD pathological process, 40 as the impairment of cholinergic neurotransmission became a well established fact in AD. 41
In the present study, the induction of AD-like pathology in rats via AlCl3 supplementation resulted in a significant decrease in brain BDNF level. Inflammatory stimulation strongly decreases the generation and release of the neurotrophic factors, which further contributes to neurodegeneration. 42 Aluminium has been found to induce the inflammatory genes leading to the production of a number of proinflammatory cytokines. 43 Parallel to the increasing generation of these cytokines, a dramatic decrease in the nerve growth factor and BDNF was demonstrated after aluminium intoxication. 44 The second possible mechanism by which aluminium could decrease brain BDNF in the present study is related to aluminium-induced Aβ formation in the brain. Aβ has been shown to reduce BDNF content and function 45 via imparing BDNF retrograde transport. Thus, Aβ could reduce BDNF signalling and this may underlie the synaptic dysfunction observed in AD patients. 46 In view of neuroprotective effects of BDNF, the demonstrated decrease in BDNF in AD patients may contribute to the development of this neurodegenerative disease due to a lack of neurotrophic support. 47 These observations assess the contribution of aluminium in the pathophysiology of AD.
The induction of AD-like pathology in rats by AlCl3 administration in the present study produced a significant elevation in brain TGF-β level. This finding could be explained by the stimulatory effect of aluminium on Aβ deposition in the brain, as it has been suggested that Aβ could induce a remarkable inflammatory cascade leading to the production of inflammatory cytokines. 48 Also, the experimental studies on microglial cells stimulated by 1–42 peptide of Aβ showed high transcription level of many proinflammatory cytokine genes such as Tumor necrosis factor-alpha (TNF-α), monocyte chemoattarctant protein-1 (MCP-1), IL-8 and TGF-β. 49,50 These observations are in consistent with other reports that showed elevations of TGF-β expression and immunoreactivity in AD patients. 51 Our findings are greatly supported by the study of Salins et al. 52 who stated that TGF-β level was markedly upregulated in cortical brain regions of mouse model of familial AD.
Our results showed a significant decrease in brain IGF-1 level in AD-like pathology model due to aluminium administration. On the basis of decreased IGF-1 concentrations in AD, some authors have suggested that disrupted IGF-1 input into the brain may be involved in the pathogenesis of amyloidosis and that changed the IGF-1 signalling may potentially lead to amyloidosis. 53 IGF-1 resistance at the blood–brain barrier is a pathogenic event in AD, and the local alterations in insulin like growth factor binding proteins (IGFBPs) may lead to a loss of IGF-1 input at the blood–brain barriers in AD. 54 Moreover, as the increased levels of oxidative stress are associated with the aging process, 55 and also impair neuronal IGF-1 signalling, 56 oxidative stress may constitute a mechanism for IGF-1 resistance in the aging brain. This mechanism could also involve in our AD-like pathology model as aluminium is a well known pro-oxidant that induces oxidative stress in the brain tissue 57 leading to IGF-1 resistance in AD brain.
Significant increase in brain Fas level has been detected after 45 days of AlCl3 administration to induce AD-like pathology model in the current study. A number of studies suggested a possible connection between oxidative stress and increases in Fas/FasL expression in brain astrocytes, microglia and neurons. 58 –60 Moreover, Fas and FasL have been reported to participate in Aβ-induced neuronal death. 61 Morishima et al. 62 provided evidence that Aβ induces FasL expression in a Jun N-terminal kinase (JNK3)-dependent manner. In agreement with our results Choi et al. 63 reported that Fas and FasL are elevated in the compromised AD brains.
The present findings indicated that our AD-like pathology model showed a significant increase in brain IL-6 level. It has been assumed that Aβ induced by aluminium administration activates microglia and astrocytes upregulating cytokines production including IL-6. 48 Also, Aβ could induce IL-6 expression in several types of brain cells. 64,65 Furthermore, IL-6 enhances neuronal damage induced by Aβ. 66 The destructive processes including neurodegneration gliosis and progressive neurological diseases are mediated by the overexpression of many cytokines including IL-6. 67 Apelt and Schliebs 68 reported that in patients with AD, IL-6 is increased locally around amyloid plaques.
AD is characterized by the excessive deposition of amyloid β-peptides (Aβ peptides) in the brain. In the non-amyloidogenic pathway, the amyloid precursor protein (APP) is cleaved by the α-secretase within the Aβ peptide sequence. Proteinases of the ADAM family (a disintegrin and metalloproteinase) are the main candidates as physiologically relevant α-secretases. 69
Three members of the ADAM family, ADAM9, ADAM10 and ADAM17, can act as α-secretases in various cell lines. 70,71 ADAM10 can cleave APP-derived peptides at the main α-secretase cleavage site between the positions 16 and 17 of the Aβ region and it is expressed in mouse and human brain. 72,73
In the present study, a significant decrease in expression levels of ADAM9 and ADAM10 genes was demonstrated in AlCl3-administered rats, which further confirmed the implication of aluminium in the pathogenesis of AD. This finding indicates that aluminium could promote the amyloidogenic pathway in the brain that results in Aβ deposition as shown in our histological study (Figures 4(a) to (g)).
The data in the present work revealed that the treatment of AD group with either SP or NK resulted in significant modulation in the different studied biochemical parameters. The detected positive effects of these enzymes could be attributed, at least in part, to the ability of SP 74 and NK 75 to cross the blood–brain barrier.
The proposed mechanisms for the beneficial effects of SP and/or NK may be related to the ability of SP to induce the degradation of insoluble proteins. 10 Similarly, NK possesses high amyloid-degrading ability, which suggests the usefulness of this enzyme in the treatment of amyloid-related diseases. 3 Thus, these enzymes might dissolve Aβ protein that accumulates in the brain due to aluminium neurotoxicity. This explanation was well documented by our histological findings (Figures 5(a) and 6(c)). As mentioned above, Aβ plays an important role in the activation of AchE activity in the brain besides its reducing effect on brain BDNF content. Therefore, the degradation of Aβ by these enzymes leads to the inhibition of brain AchE activity and restoration of BDNF brain level. Moreover, the dissolution of Aβ by SP and/or NK enzymes results in the blocking of the inflammatory cascade and preventing the production of the proinflammatory cytokines such as TGF-β and IL-6 in the brain. Also, the elimination of Aβ deposits helps in correcting the disruption of brain IGF-1 level and managing the process of apoptosis and neuronal death due to the suppressing of Fas expression and in turn its level in the brain.
SP has been found to have anti-inflammatory effect 9,76 and the efficacy of SP in the treatment of a number of inflammatory conditions has been reported. 11,12 This property of SP represents an additional mechanism by which SP could correct the activity of brain AchE and the levels of BDNF, TGF-β and IL-6 in the brain.
SP and NK are belonging to proteolytic enzymes family. These enzymes have been reported to have free radical-scavenging property, 77 which enables these enzymes to reduce reactive oxygen species accumulation in the brain. By this way, both SP and NK could elevate IGF-1 and suppress Fas levels in the brain.
Regarding the effect of SP and/or NK on the expression levels of ADAM9 and ADAM10 genes in the brain tissue, our results revealed that the high doses of these enzymes could increase the expression levels of ADAM9 and ADAM10 genes. Also, the low dose of NK could elevate the expression level of ADAM10 gene in the brain tissue of the treated animals. These results indicate that these enzymes may have the ability to shift the amyloidogenic pathway to the non-amyloidogenic one via increasing the expression level of ADAM9 and ADAM10 genes in the brain. To the best of the author’s knowledge, this is the first published study that highlighted the efficacy of SP and NK in alleviating the generation of Aβ in the brain via promoting the activity of α-secretase-like action.
Several studies investigating the systemic use of SP 10,12,78 and NK 79 have demonstrated no adverse effects. In the present study, the lack of a dose dependence may confirm the safety of these enzymes as they are recommended as dietary supplements.
In conclusion, the results of the present work support our hypothesis that the oral administration of SP or NK would have beneficial effects against AD. Thus, they may represent good remedies for the management of AD via their proteolytic activity, antioxidant capacity and anti-amyloidogenic effect.
Footnotes
Funding
This research received no specific grant from any funding agency in the public, commercial, or not-for-profit sectors.