
Abstract
Pollen analysis was conducted on core materials that were deposited over the last 2000 cal years BP in Mkhuze wetland, KwaZulu-Natal. The objective was to reconstruct past vegetation and infer past changes in climate or possible human disturbances by enhancing both pollen and non-pollen proxy data in the summer rainfall region. Palynological results show a dominance of Poaceae (grasses >70%) that suggests a predominance of grassy vegetation in and around the swamp with some woodland and forest elements from the surroundings. Between 1700 and 1200 cal years BP, fungal spores, cryptogams, wetland plants, Podocarpus, and other forest pollen elements declined. Bushveld tree pollen of Spirostachys increased, indicating a decrease in moisture availability (humidity) and a change from forest to an open savanna vegetation. A peak of microscopic charcoal and Poaceae with a corresponding decrease in Spirostachys pollen around 800–500 cal years BP suggests an increase in wildfires under dry conditions that may have been caused by anthropogenic forest clearance for farming by the Iron Age people. The increase in microscopic charcoal, charred cuticles, a peak of Amaranthaceae pollen, and a corresponding decrease in trees and grasses in the last 280–200 cal years BP may be due to regional drought. The appearance of exotic Pinus pollen during the most recent period corresponds to the introduction of alien vegetation by European settlers. We compared changes in the pollen record of the Mkhuze freshwater swamp with a coeval section of a previously published longer pollen sequence in the nearby Lake St Lucia estuary to confirm if regional vegetation trends can also be observed in the section. Wider regional environmental trends were also assessed using other non-pollen proxies like isotopic data for comparison. Results align with δ18O, d13C and N15 studies, although some, especially the latter, may not suggest parallel moisture conditions over the distances involved. This indicates that diverse environmental conditions prevailed in the region in the last c. 2000 years BP.
Introduction
Paleoenvironmental studies in Southern Africa, utilizing pollen analysis, have yielded fundamental insight into the changes in vegetation cover, which can be further expanded by examining wetlands in this region where pollen-bearing deposits are typically limited (Coetzee, 1967; Effiom et al., 2024; Ekblom, 2008; Ekblom et al., 2014; Finch and Hill, 2008; Neumann et al., 2010, 2011, 2014, 2025; Quick et al., 2016, 2018; Scott, 1982; Scott et al., 2012, 2022; Scott, 1989, 1999; Scott and Lee-Thorp, 2004; Scott and Neumann, 2018; van Zinderen Bakker, 1957, 1982; van Zinderen Bakker and Coetzee, 1988).
Wetlands, which serve as sources of proxy evidence for past environmental changes utilized in pollen analysis, have experienced significant degradation and biodiversity loss in KwaZulu-Natal over the past century. This is attributed to intensified agricultural practices, urban expansion, and climate change (Kotze et al., 1995; Kotze and O’Connor, 2000). To enhance the understanding of past regional vegetation changes under the influence of climate change and human activity in light of the imminent future loss of this resource, we present a comprehensive palynological analysis of the Mkhuze core along with a regional comparison of both pollen and non-pollen data, including δ18O, δ 13C and δN15 studies at Cold Air Cave and Marakabi, representing areas at a distance away toward the interior of the summer rainfall region (Chase et al., 2022; Holmgren et al., 2003).
The northeastern sub-humid coast of KwaZulu-Natal, South Africa, consists of an extensive but under-utilized wetland scattered on the coastal plain with potential for pollen studies. Its potential is due to its many sedimentary environments, especially peat deposits (Grundling et al., 1998; Botha et al., 2018; Figure 1). Here we follow up on other pollen records for this region (e.g., Effiom et al., 2024; Finch and Hill, 2008; Mazus, 2000; Neumann et al., 2008, 2010).
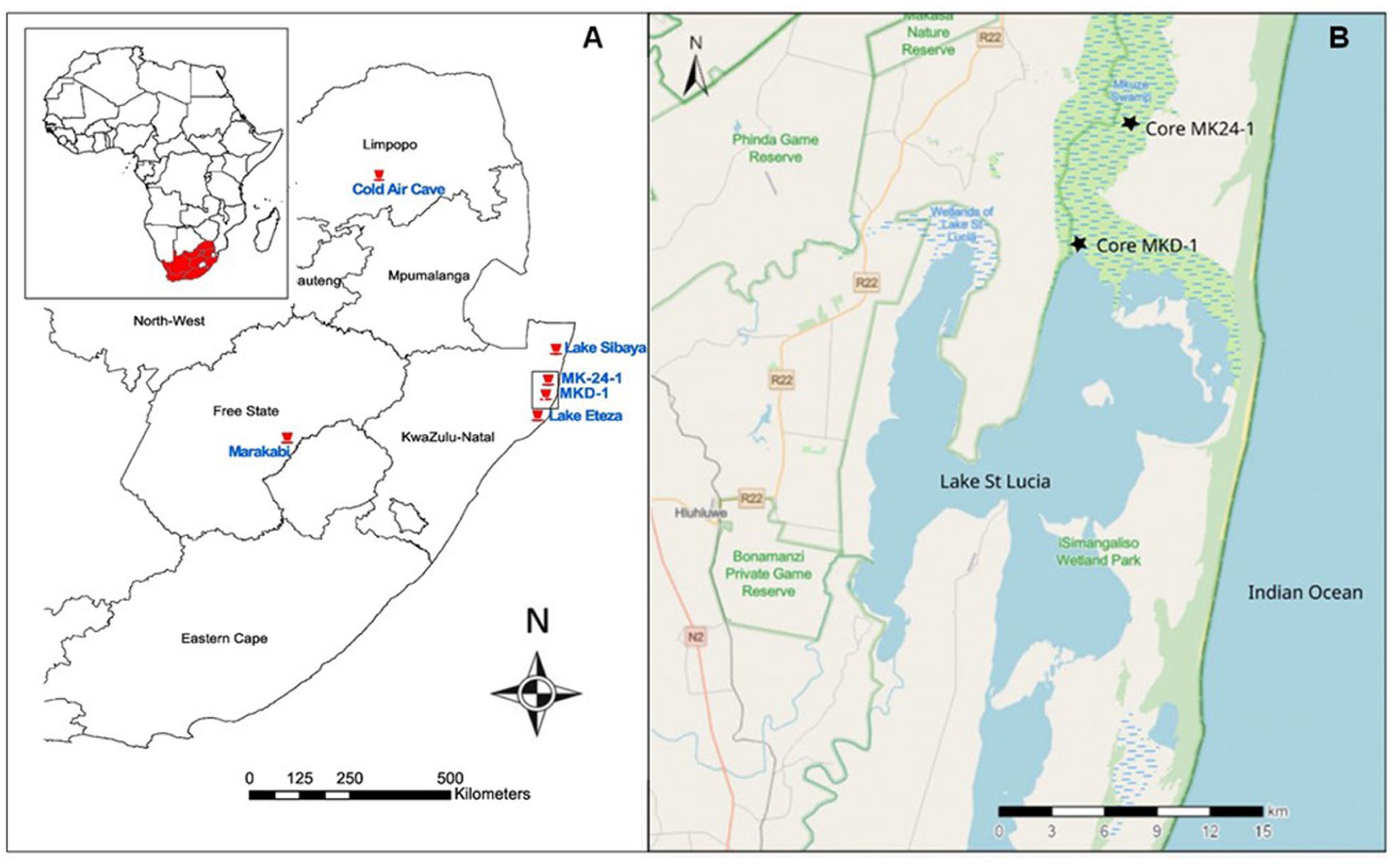
(a) Map showing the study site (Core MK24-1; this study) in relation to some other proxy sequences discussed here, viz., MKD-1 (Effiom et al., 2024), Lake Sibaya (Neumann et al., 2008), Lake Eteza (Neumann et al., 2010), Cold Air Cave (Holmgren et al., 2003) and Marakabi (Chase et al., 2022) and (b) Location of Core MK24-1 in the Mkuze swamp and the nearby palynological site MKD-1 in Lake St. Lucia.
The study of Mazus (2000) focused only on Podocarpus pollen and showed a decline in forest in northern KwaZulu Natal during the late-Holocene. Ekblom (2008) studied the forest–savanna dynamics covering the last 1400 years in the coastal lowland of southern Mozambique at Chibuene. She reported that although the expansion of savannas at the cost of forests has been attributed to human influence, in fact, the decline of forest is correlated with repeated dry spells associated with the “Little Ice Age” (LIA 500–250 cal years BP). In another study by Ekblom et al. (2014), in the coastal forest and Miombo woodland over the last 1600 years in the Vilankulo region, about 7 km north of Chibuene, southern Mozambique, they reported that the changes/variability in Brachystegia, as it declined after 1250 cal years BP, seemed to be driven by hydrological changes related to climate variability rather than land use changes. Neumann et al. (2008), in their study at Lake Sibaya, suggested that the early Iron Age had a moist climate. They also showed that an increase in algae and cereal pollen since 150–300 cal years BP reflected human activities. From Lake Eteza sediments, Neumann et al. (2010), who confirmed Mazus’ work, inferred that the decrease in Podocarpus pollen during the late-Holocene and the corresponding increase in Chenopodiaceae/Amaranthaceae and grass pollen coincided with a return to lower sea level and drier conditions. Effiom et al. (2024), working on sediments from Lake St. Lucia (MKD-1), suggested that the decline and disappearance of Podocarpus in the late-Holocene was due to dry conditions. According to Neumann et al. (2025), vegetation in Delagoa Bight in southern Mozambique shifted from late-Pleistocene open, cool-adapted communities to more humid Holocene savanna-forest assemblages with widespread Podocarpus over the last c. 16,600 years, followed by a late-Holocene trend toward drier conditions linked to climate and sea level fluctuations. In summary, all pollen records in the region indicate a drier (drought) climate in the late-Holocene.
Non-pollen proxies study in the IOCB and in the wider summer rainfall region disagree on drought periods for the last 2000 cal years BP (Chase et al., 2022; Holmgren et al., 2003; Humphries et al., 2020; Strobel et al., 2022). These studies used carbon, oxygen and nitrogen isotopes, geochemical data (leaf waxes, hemicellulose sugar, hydrogen, and carbon isotopes etc.), in a multiproxy approach. Several reported that drought phases occurred before the last 2000 cal years BP, with Holmgren et al. (2003) showing an increase in temperature between 1000 and 2000 cal years BP. Chase et al. (2022) reported a steady rise in humidity during the late-Holocene (the last 6680 cal years BP). In other cases, covering a wide region from the coast to the interior, the opposite was reported by Miller et al. (2019) at Mfabeni using stable carbon and hydrogen isotopes of plant-waxes, and House et al. (2022) using charcoal at Wonderwerk Cave. They suggested that drier conditions prevailed during the last c. 2000 years BP. Due to the above conflicts, there is a need to carry out further palynological analyses in the IOCB, such as this study on the Mkhuze swamp, and compare the findings with both palynological data and selected non-pollen data available for the last 2000 cal year BP, which will contribute to the Late-Holocene paleoenvironmental data. Moreover, studies by Gensel et al. (2022) on the origin, transport, and retention of fluvial sedimentary organic matter in the Mkhuze wetland indicated that organic matter originates from the surrounding catchment areas and is deposited throughout the wetland system, extending to the Mkhuze swamp. In contrast, the downstream region of Lake St. Lucia exhibits locally derived signals rather than integrated signals from the river catchment. Therefore, results from the Mkhuze swamp core (MK24-1), which is freshwater, will be compared with the Lake St. Lucia core (MKD-1), which is brackish and is about 60 km apart (see Figure 1b). The comparison will be done to see if the cores correlate both in diversity of palynomorphs and reconstructed vegetation changes, as it will provide more information about the depositional environment.
Study site
Background
As part of the Mkhuze wetland system, the Mkhuze River originates from the midland catchment below the Drakensberg Escarpment and is the largest contributor of sediment and freshwater to the northern basin of Lake St Lucia (Hansmann, 1993; Whitfield and Taylor, 2009). The Mkhuze swamp, located in the Indian Ocean Coastal Belt Biome (IOCB), is a wetland system of 450 km2 consisting of streams and floodplain pans that developed along the lower reaches of the river (Ellery et al., 2003). The swamp and river are utilized by rural people that depend on natural resources, subsistence farming, and grazing on the floodplain (Andrén, 2001; Koaleli, 1999; Stormanns, 1987). The wetland forms part of the IOCB that houses the southernmost extension of the tropical East African coastal forest trees, lianas, and epiphytes (Mucina et al., 2006). The biome is highly diverse in terms of flora and fauna, but the ecosystem is susceptible to effects of human disturbance and climate change (Lawes et al., 2007; Smith Ehlers et al., 2017).
The Mkhuze swamp was chosen for the present study as it is a significant ecosystem due to its species richness and heterogeneity as well as the fact that it has been relatively undisturbed by anthropogenic activities in recent times and that it serves as a refugium for a diverse group of wildlife and plants (McCarthy and Hancox, 2000; Stormanns, 1987).
Climate and geology
Along the coastal belt of northern KwaZulu-Natal and southern Mozambique, the climate is humid and subtropical with mild, dry winters and wet, hot summers (Ellery et al., 2003). Monthly average precipitation is the highest in February and the lowest in July, and generally, high precipitation occurs from October to March and usually less from April to September (Nomadseason, 2024). The mean temperature during summer is 25°C and 18°C in winter, with the area experiencing over 247 days of 25°C mean temperature (Nomadseason, 2024). Average rainfall is approximately 1000 mm along the coast and 600 mm at 17 km inland in the Mkhuze Game Reserve (Ellery et al., 2003).
The catchment area of the Mkhuze River covers approximately 5000 km2 with the upper catchment comprising rocks of the Karoo Supergroup that vary from sandstone to shale and include numerous dolerite intrusions in the interior as well as volcanic rocks of the Jozini Formation in the Lebombo Mountains (Ellery et al., 2003). Sandy or reworked marine deposits are a minor component east of the Lebombo Mountain (Botha et al., 2018; Watkeys et al., 1993).
Most of the water supplied to the Mkhuze swamps is from rainfall, local streamflow, and groundwater seepage, including southward flow from some floodplain pans and swamps (Ellery et al., 2003).
Vegetation
The riverine forest along the Mkhuze River comprises trees such as Rauvolfia caffra, Ficus sycomorus, Voacanga thouarsii, and Blighia unijugata, which are found in isolated patches across a narrow band along the river (Neal, 2001; Stormanns, 1987). The open grasslands that border the riverine forest are covered by Echinocloa pyramidalis, and the floodplains are fringed by stands of Vachellia xanthophloea. Phragmites australis, Phragmites mauritianus, Cyperus papyrus, Cyperus dives and patches of Typha capensis dominate the floodplain pan fringes and depressions while infrequently flooded Cynodon dactylon grasslands dominate the distal end of the system (Neal, 2001).
Materials and methods
Core collection and radiocarbon dating
Core MK24-1 (27°51.365′S 32°28.760′E) was recovered from the Mkhuze swamp close to the Mkhuze River Delta which is a bayhead delta that discharges into the northern-most part of Lake St. Lucia (Figure 1b). It was retrieved using a 5 cm diameter clear plexiglass tube, which was pushed and hammered into the swamp. This core is 90 cm long. The cored sediments were subsampled to give a total of 30 subsamples for palynology. Pollen analyses were done on the sediments between 0.5 and 91.5 cm of core MK24-1.
Three sediment samples were submitted for radiocarbon dating, and radiocarbon ages were calibrated to calendar years using the Southern Hemisphere calibration SHCal20 (Hogg et al., 2020); a Bayesian age-depth model was developed using Bacon 2.2 software (Blaauw and Christen, 2011).
Pollen analysis
Pollen grains were extracted from the sediment following the standard method for sediments (Erdtman, 1969; Faegri and Iversen, 1966). The samples were treated with 40% HF overnight to dissolve silicates, rinsed, and followed by the addition of 10% KOH to remove humic acid as well as other organics. During this process, the sample was boiled for 5 min in distilled water, then centrifuged and decanted. Mineral separation with ZnCl2 (specific gravity 2) was carried out before acetolysis. Acetolysis is the removal of lipids and proteins, including the inner organic pollen wall (intine) and cytoplasm, using acetic anhydride and sulfuric acid in the ratio of 9:1 (Erdtman, 1960). After rinsing and centrifugation, the residue was mounted on microscopic slides in glycerin jelly. An Olympus BX51 stereomicroscope was used for the pollen analysis, and photographs were taken using a 100x oil immersion objective and an Olympus C30 digital camera.
Pollen grain identifications were supported by comparison with modern pollen reference collections at the Evolutionary Studies Institute, University of the Witwatersrand, and Department of Plant Sciences, University of the Free State; pollen atlases such as Bonnefille and Riollet (1980), Scott (1982), Gosling et al. (2013), Schüler and Hemp (2016), and Effiom et al. (2025); and online African pollen databanks (Lezine, 2005). Recovered micro charcoal fragments were also analyzed and grouped as “charcoal >100 µm and charcoal <100 µm.” This was done to separate the long and short distance derived charcoal in the study site (Vachula et al., 2018), which helps to reconstruct fire history in the environment.
Statistical analysis
The pollen diagram was calculated and drawn using Strat.Plot in rioja (R software), which is designed for managing and graphing stratigraphic data, especially paleontological data (Juggins, 2012; Juggins and Juggins, 2019). The pollen diagram was further divided into zones based on the internal variation within and between groups of palynomorphs at different depths/ages using Constrained Incremental Sums of Squares (CONISS, R studio: http://www.rstudio.com/). This method helps identify pollen assemblages and abundance or sparsity zones, which in turn provides information about changes in vegetation across the different cores. Finally, principal component analysis (PCA) was performed on prominent palynomorphs and non-pollen palynomorphs (NPPs) percentages using the correlation function in the PAST program (Hammer et al., 2001). The data set represents all the most prominent pollen and non-pollen palynomorph types (those whose totals exceed 4%), like cryptogams and fungal spores, to ordinate the main local variation of taxa in MK24-1.
Pollen results were further compared with other non-pollen proxy evidence from Marakabi and Cold Air Cave (Chase et al., 2022; Holmgren et al., 2003) to determine if wider regional climate events can be observed in MK24-1.
Results/discussion
Dating and sedimentation rate
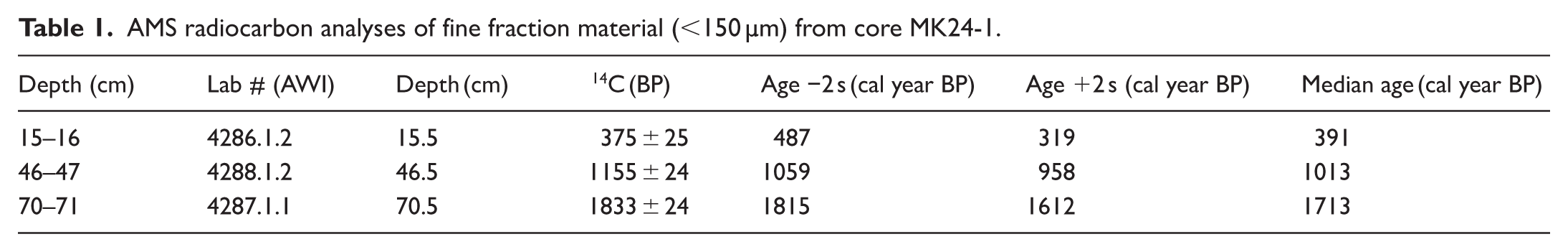
The radiocarbon dates were measured at depths between 15.5 and 70.5 cm (Table 1; Figure 2).
AMS radiocarbon analyses of fine fraction material (<150 µm) from core MK24-1.
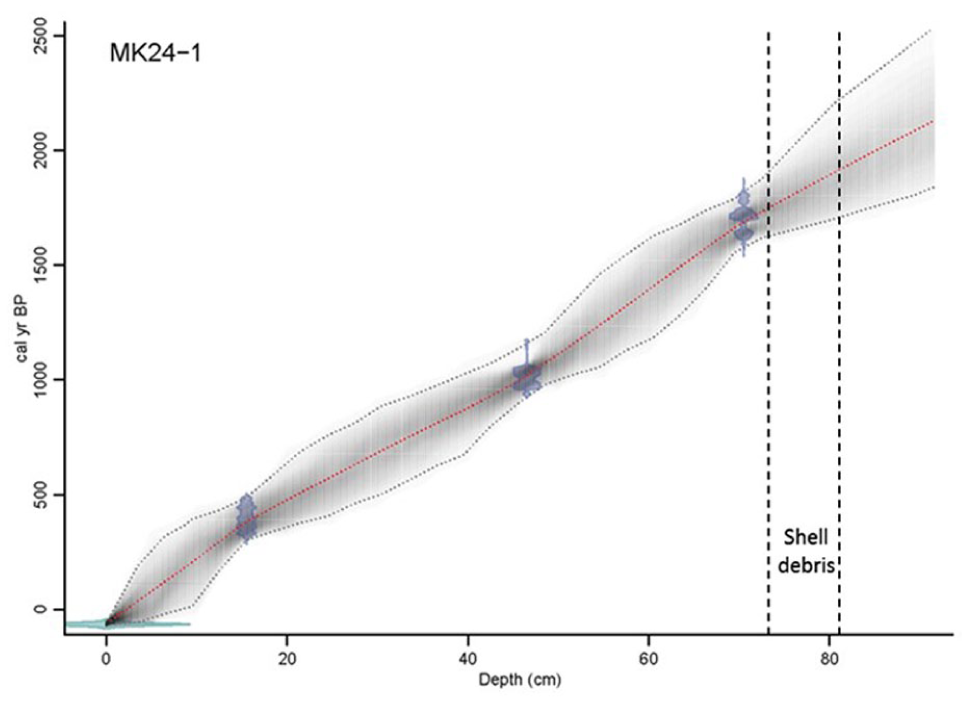
Age model using Bayesian statistics (Bacon software v2.2) for core MK24-1.
There is no radiocarbon date for the basal sediment due to the shell layer, which is conceived to be problematic for radiocarbon dating because of the inert instability of shell CaCO3 (Douka et al., 2010). MK24-1 sedimentation rate averaged ~0.01 cm/year from 1038 to 44 cal years BP and from 1753 to 1124 cal years BP but decreased to 0.02 cm yearly between 1078 and 1051 cal years BP. No sedimentation occurred between 1814 and 1784 cal years BP, coinciding with the shell debris (Figure 2). In the absence of sediment or organic matter deposition, biogenic materials such as mollusk shells accumulate. During this period of undisturbed conditions, the Mkhuze wetland maintained a stable and transparent water column, facilitating sunlight penetration (Schallenberg and Sorrell, 2009). The increase of light penetration fosters the proliferation of macrophytes, which subsequently offer habitat for invertebrates, including mollusks, thereby resulting in shell deposits (Schallenberg and Sorrell, 2009).
The lithological units of the Mkhuze swamp core are described in Figure 3. The base of the core (92–82 cm) consists of dark clay sediment rich in organic matter (humus). This deposit is sharply overlain by a shell bed layer and sand ( Figure 3). From 74 to 68 cm, there is reddish clay sand (rich in iron oxide), which can generally be associated with the weathering of crystalline and metamorphic rocks in areas of high rainfall (Bhargava, 2022). The sand-layered grained horizon is seen between 68 and 64 cm and is overlain by clay sand (65–12 cm). The upper 11 cm of the sediment profile is characterized by blackish coarse soil, which is generally rich in humus, phosphorus, and ammonia (McAleese, 2021).

Lithology of core MK24-1.
Pollen analysis and principal component analysis
The palynomorphs percentages were grouped into broad ecological groups such as neophytes, forest/woodland elements, herbs/shrublets, wetland elements, and cryptogams (Figure 4). Four pollen zones, from the oldest to the youngest, Zones 1–4, were generated based on changes in composition and abundance of individual pollen types as determined by CONISS (Figure 4 and Table 2). Principal Component Analysis (PCA) loadings of selected pollen taxa for MK24-1 are shown in Figure 5, and the scores are shown in Figure 6.
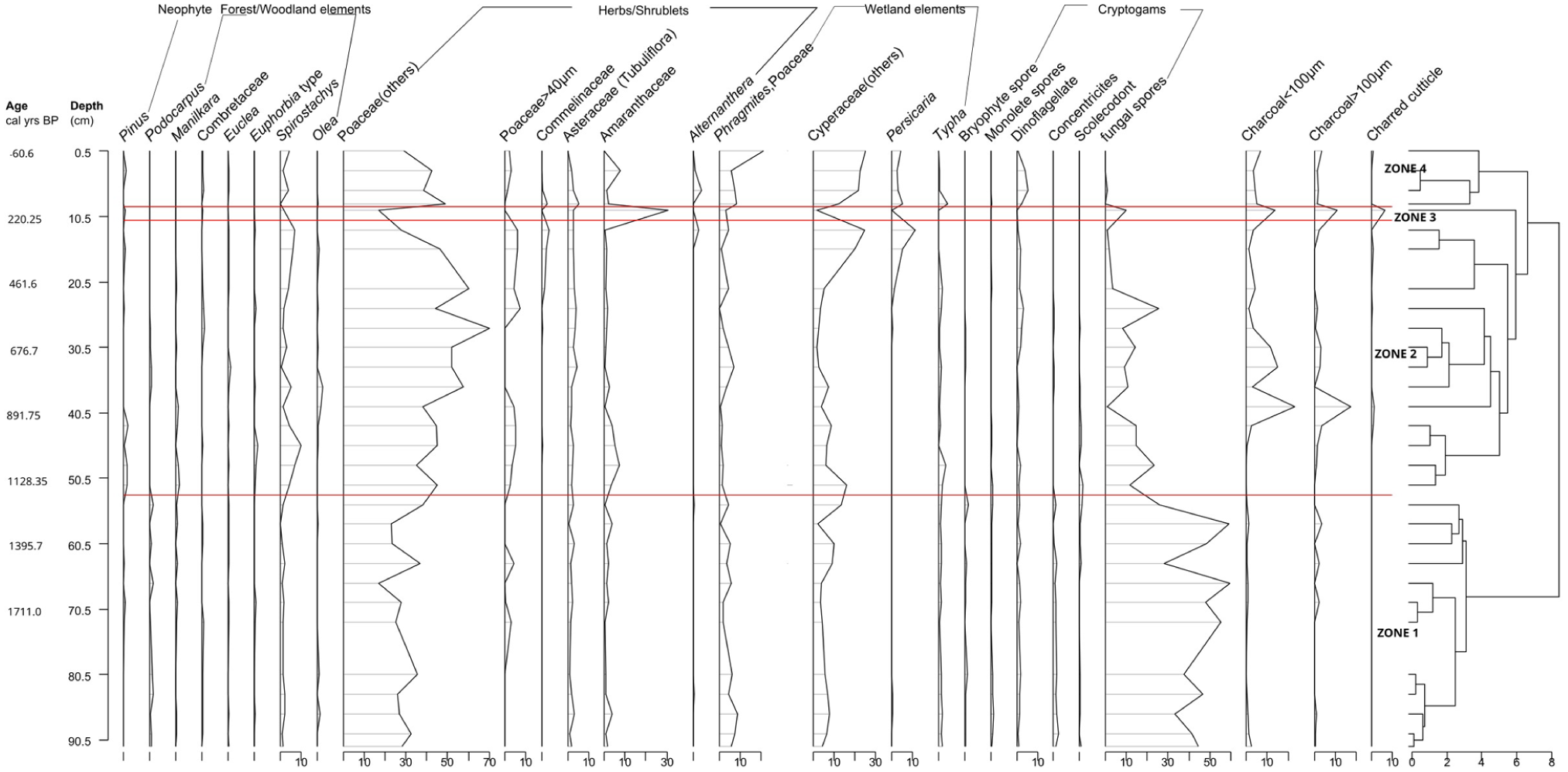
Pollen and cryptogamic spore percentage diagram for MK24-1 based on the pollen sum, plotted according to depth and showing the four pollen zones and groupings.
Summarized paleoenvironmental interpretation for MK24-1.

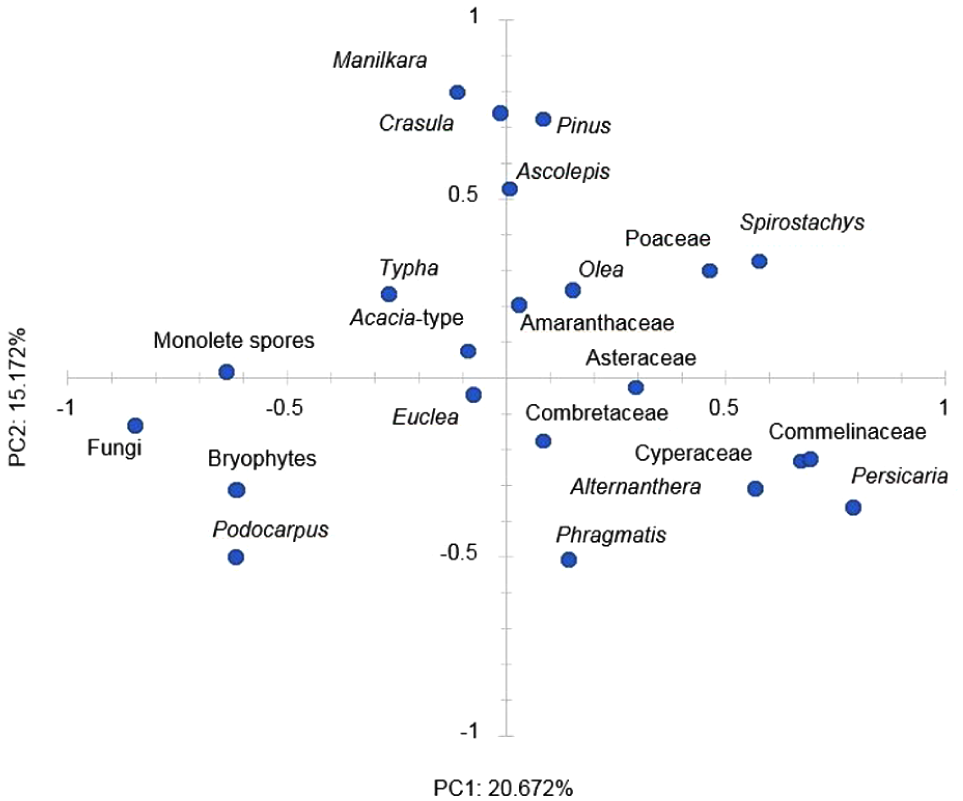
PCA biplot for PC1 and PC2 with variances of the most prominent palynomorph taxa (pollen and NPP’s) using correlation in the PAST program.
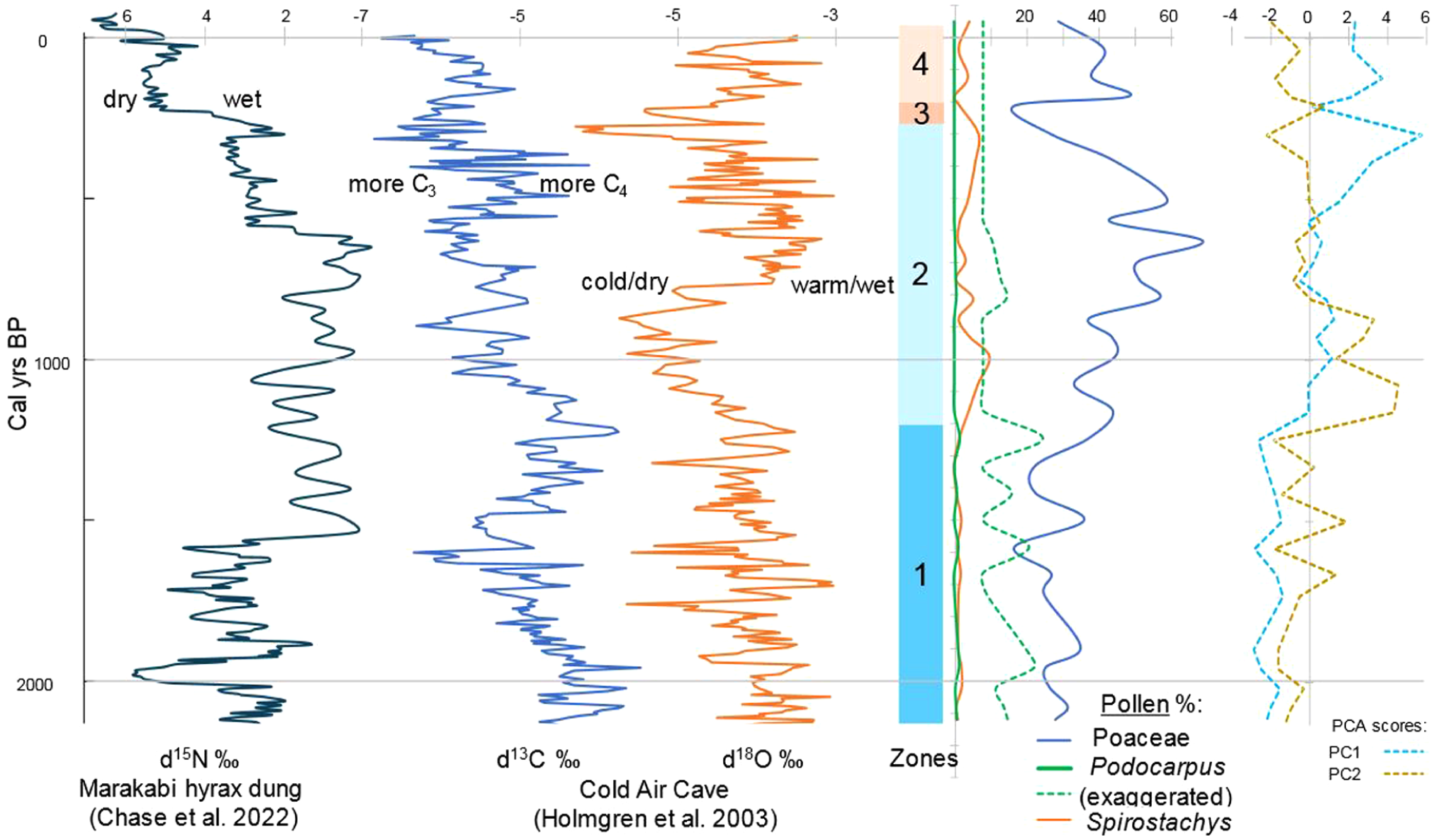
Pollen zones, Poaceae, Podocarpus and Spirostachys pollen %’s for MK24-1 and PC1 and PC2 scores for the most important palynomorphs and NPPs in MK24-1 on the right compared with isotopic contents of the Marakabi and Cold Air Cave localities (Chase et al., 2022; Holmgren et al., 2003) on the left.
PC1 accounts for 20.671%, while PC2 accounts for 15.172% of the variation in pollen composition. PC1 reveals a negative loading for Podocarpus, cryptogams, Euclea, Crassula, Manilkara, Typha, and fungal spores and a positive loading for Spirostachys, Asteraceae, Poaceae, Amaranthaceae, Combretaceae, Commelinaceae, Phragmites, Cyperaceae, and Persicaria.
In Figure 5, the distribution along PC shows a dichotomy between cryptogams, fungal spores, and Podocarpus negative loadings against most other palynomorphs with positive loadings, resulting in negative scores of this factor in the lower part (Zone 1) of the sequence as seen in Figure 6 up to c. 1200 cal year BP. PC2 shows a mixture of dryland and moisture-loving types, of which the loadings also cannot easily be assigned to environmental conditions, but its association with Podocarpus and Phragmites tentatively indicates relatively moist conditions in Zone 1.
Vegetation history
The Mkhuze wetland sequence is characterized by Poaceae pollen (20%–70%), indicating a dominance of grassy environments interspersed with trees. Spirostachys pollen dominated the arboreal spectra, and based on pollen percentages of taxa, we can deduce the persistence of grasses; hence, vegetation consists of woodland/grassland mosaics. This agrees with Burney’s (1988) studies of modern pollen rain in Madagascar, as proportions of arboreal pollen (AP) and non-arboreal pollen (NAP) were used to distinguish between taxa linked to grasslands (with grass pollen ⩾55%) and forest/grassland mosaics (Burney, 1988; Julier et al., 2021). The existence of wetland species, including Typha and sedges, signifies the continuous presence of a wetland, which has fluctuated and expanded in recent times.
Paleoenvironmental interpretations for Mkhuze swamp core
The pollen zones are described below and summarized in Table 2.
Zone 1 (1700–1200 cal years BP)
A moderately warm freshwater environment is suggested by the presence of Podocarpus pollen (less than 5%), various forest components and cryptogams, Poaceae (30%–40%), Phragmites, Cyperaceae (7%–8%), fungal spores (60%), Pseudoschizaea, and Typha (Finch and Hill, 2008; Scott, 1999). Cryptogams indicate local moisture (Kranner et al., 2008; Vanderpoorten and Goffinet, 2009).
Zone 2 (1200–280 cal years BP)
Pollen from Spirostachys increased and peaked at 10% at c. 980 cal years BP, but then fell to < 5% at 800–500 cal years BP. In parallel, pollen from Poaceae increased and peaked at 70%, and charcoal at 15%. Additionally, there was a decrease in cryptogams, fungal spores, Typha, Pseudoschizoaea, and Phragmites pollen, with fungal spores declining from 60% to ⩽30%. While the latter decreases indicate a drier climate with less precipitation, a substantial increase in Spirostachys indicates a more open woodland vegetation. Spirostachys, Cyperaceae, and Persicaria all increased toward the end of the zone, whereas Poaceae decreased at approximately 284 years BP, suggesting a warm habitat with a disturbed swamp mosaic. Due to pollen seepage that occurs downward, Pinus is also present in this zone and is most likely a contaminant (Erdtman, 1921; Malmstrom, 1923). The identification and resolution of this contamination can be achieved with ease through comparison with the regional pollen sequence (Godwin, 1934). This has been observed in the core MKD-1 sequence, Lake Eteza, and Sibaya, where Pinus first appeared approximately 400 cal years BP (Effiom et al., 2024; Neumann et al., 2008, 2010).
Zone 3 (280–200 cal years BP)
The forest elements declined and disappeared; Poaceae, herbs, shrubs, and wetland elements also declined, whereas Amaranthaceae increased and peaked at 30% with increased charcoal and charred cuticles at ⩾10% during this period. This indicates drought leading to high evaporation and the drying out of the swamp; hence, the abundance of Amaranthaceae, as they point to a dry or saline environment (Dyer, 1975; Pooley, 2005; Scott, 1999). The corresponding increase in macro- and micro- charcoal and charred cuticles suggests local fires that may be caused by the dry weather conditions. This period is drier than zone 2.
Zone 4 (200–0 cal years BP)
The presence of Spirostachys and Pinus pollen and the increase in Poaceae, Phragmites, Cyperaceae, Persicaria, and Typha in this period indicate a relatively warm environment with increased precipitation and a disturbed wetland mosaic that is implied by the presence of Persicaria and other wetland taxa (Scott, 1999). Pinus indicates disturbances by the European settlers as they introduced and established large pine plantations in coastal areas of the Indian Ocean Coastal Belt biome in 1928–1929 and in the St. Lucia region since 1957 (Marwick, 1973; Thamm et al., 1996).
Mkhuze swamp core and Lake St. Lucia core comparison
The Mkhuze River is one of five rivers that supplies freshwater to the Lake St. Lucia estuary (see Figure 1; Taylor et al. (2006)). Gensel et al. (2022) reported that the Mkhuze Swamp captures the transported organic matter transported by the Mkhuze River, while Lake St. Lucia receives more locally derived signals. Hence, comparing pollen sequences from the Mkhuze Swamp and Lake St. Lucia is essential for distinguishing catchment-derived from local vegetation signals and clarifying the role of the Mkhuze River in shaping the sedimentary and ecological records of the system.
The Lake St. Lucia core MKD-1 (Effiom et al., 2024), which was taken from the Mkhuze River delta that drains into the northernmost part of Lake St. Lucia, showed more diversity in terms of pollen recovery but low quantity in terms of preservation when compared to the Mkhuze swamp core (MK24-1), which had low diversity but better preservation. This difference may be due to the nature of the depositional environment, as MKD-1 was taken at the mouth of the estuarine lake, trapping pollen from the wide environment, which is less biased than the input from the immediate local environment of MK24-1, which is more dominated by lower-diversity swamp elements. However, it has a higher salinity compared to the current core MK24-1 from the freshwater swamp. The estuarine lake has experienced a series of wet and dry cycles that may have affected the preservation of pollen in the cores. A similar scenario was reported by Campbell and Campbell (1994) in their experiments of palynomorph preservation in saline and freshwater sediments that had undergone wet and dry cycles.
Lake St. Lucia (MKD-1) differs from Mkhuze Swamp (MK24-1) in the recovery of foraminifera linings and mangrove species. This is because Lake St. Lucia is an estuarine lake and had experienced marine influence in the past and may possess a soil salinity level required for the germination of mangrove species as opposed to the freshwater swamp because mangroves prefer brackish water (Kathiresan and Bingham, 2001).
Apart from the differences in pollen preservation, diversity, and marine influence, the vegetation histories derived from the pollen data from core MKD-1 and MK24-1 are similar. Summaries (PC1 scores) of their pollen records show low values associated with Podocarpus assemblages beginning 2000 cal years BP (see Figure 6), followed by higher scores and an increase in Spirostachys, which later declined as Poaceae increased. Consequently, there is an inverse relationship between Podocarpus and Spirostachys (see Figure 6). The sequence also shows different environmental changes; for example, MK24-1 recorded local fires occurring between 800–500 and 280–200 cal years BP, with the earlier events presumed to be caused by anthropogenic activities such as clearing of the trees, opening it up for livestock grazing. This is evidenced by the reduction of Spirostachys pollen (woodland tree) and a corresponding increase and peak of grass pollen. The latter charcoal peak probably was caused by severe drought between 280 and 200 cal years BP, which is evidenced by the decline of several genera, with a corresponding increase in Amaranthaceae, charcoal, and charred cuticles. In contrast, MKD-1 showed little to no evidence of microcharcoal in the core. This could possibly be that the very fine charcoal is trapped and deposited in the swamp and does not reach the delta mouth of the river into the lake. Finally, disturbances from human activities can be seen in both cores over the past 200 cal years BP. Precipitation also increased during that period, as shown by the increase in Typha and wetland species. There is similarity in both cores based on pollen taxa and so vegetation characters.
In summary, the subtropical wetland vegetation represented by the Mkhuze swamp core (MK24-1) reflects a local signal. In contrast, the Lake St. Lucia core (MKD-1) serves as a regional signal due to the recovery of pollen grains indicative of mangroves, wooded grasslands, and coastal flora (see Table 3).
Core MKD-1 (Lake St. Lucia) and core MK24-1 (Mkhuze swamp): comparison of palynological results.

Regional environmental interpretation
Regional trend of podocarpus and spirostachys
Podocarpus and Spirostachys are indicators of forest and woodland/bushveld, respectively (Coates Palgrave et al., 1985; Lennox and Bamford, 2015; Mtsweni et al., 2005). Podocarpus is indirectly proportionate to Spirostachys; hence, a decrease in the forest tree (Podocarpus) corresponds with an increase in woodland trees such as Spirostachys. In the last 2000 cal years BP, both core MKD-1 and MK24-1 showed a decline in Podocarpus and an increase in Spirostachys, suggesting a change from a forested environment to a more open-woodland environment. This is evidenced at various locations along the KwaZulu-Natal coastline, as noted by Mazus (2000) and others, including Lake Eteza (Neumann et al., 2010; Scott and Steenkamp, 1996) and Lake Sibaya (Neumann et al., 2008). Recent findings in the pollen record from marine sediments off the coast near Maputo, Mozambique, confirm the regional significance of this change (Neumann et al., 2025). Based on the ratio of Podocarpus to Spirostachys pollen across the three sites, Lake St. Lucia (MKD-1), Lake Eteza, and Lake Sibaya (Effiom et al., 2024), it seems that Spirostachys prominence developed in a north-to-south direction so that declining Podocarpus communities persisted in the south for longer. While the main reason for the decline of Podocarpus may be climatic as a result of drought gradually spreading from the north, the impact of Iron Age herders should not be discounted as an influence on its ratio to Spirostachys, which would have been favored as firewood, while the taller Podocarpus trees would have been more difficult to fell.
Wider regional climatic implication
To consider the palynology of MK24-1 in terms of the wider regional environmental development, we compared our results with those of other proxy data (Figures 1 and 6). IOCB pollen data from the different sites suggest moderate to high tree/shrub percentages between 2000 and 700 cal years BP for different pollen sites in the region like MKD-1.
Relatively high but fluctuating temperatures ranging from cool, dry indications to warm, wet conditions in the interior are reported by Holmgren et al. (2003) and Chase et al. (2022) for the last c. 2100 years BP. From c. 2100 to 1400 cal years BP, the Podocarpus pollen presence probably occurred under moderately warm conditions, assuming temperature changes inferred from Holmgren et al. (2003) are regional. Our findings indicate relatively stable moisture levels that became less humid at the conclusion of Zone 1 (2000–1200 cal years BP), coinciding with the decline and retreat of Podocarpus and the growth of Spirostachys. Spirostachys pollen is notable, signifying fairly warm and dry conditions. This is consistent with the δ18O results at Cold Air Cave (Holmgren et al., 2003). The N15 observations of Marakabi by Chase et al. (2022) indicate a steady rise in humidity throughout the late-Holocene until around c. 200 years BP (Figure 6). Despite Mkhuze wetland and Marakabi both being situated in the southern Africa summer-rain region, the difference in moisture conditions may arise from different regional climate forcings between the areas.
The period between c. 1200 and c. 700 cal years BP coincides with the decrease in δ18O in the stalagmite at Cold Air Cave, suggesting a dry and colder environment (Holmgren et al., 2003; Figure 6). which can also be seen at the beginning of Zone 2, as there was a transition from a forested environment to an open vegetation (Figure 6). Neumann et al. (2010) also reported that dry conditions and low sea levels led to the decline of forest elements during this period. A similar trend in pollen sequence was seen at Lake Sibaya and Lake St Lucia (Effiom et al., 2024; Neumann et al., 2008).
Between c. 700 and c. 300 years BP, δ18O increased, suggesting that the environment became warm and wet, which corresponds with an increase in C4 grasses at Cold Air Cave (Holmgren et al., 2003) and an increase and peak of grasses in Zone 2 at the Mkhuze swamp. However, between c. 300 and c. 200 cal years BP, δ18O declined sharply, suggesting that the environment became cold and dry, corresponding with the Northern Hemisphere’s Little Ice Age (LIA; 500–250 cal years BP). Ekblom (2008) and Ekblom et al. (2014) reported that there have been repeated dry spells associated with the LIA in the Indian Ocean Coastal Belt Biome, and this can be supported by drier conditions indicated in Zone 3 (MK24-1; see Figures 4 and 6). The opposite environment was reported at Marakabi as N15 declined, suggesting that the environment at Marakabi was wet during that time (Chase et al., 2022). The last c. 200 years BP witness a return of wetter conditions at the Mkhuze swamp which corresponds with an increase in δ18O at the Cold Air Cave, suggesting a warm and wet condition (Holmgren et al., 2003). However, N15 increased at Marakabi, indicating that drier conditions prevailed in Marakabi in the last c. 200 years BP (Chase et al., 2022) as opposed to the warm and wet conditions that prevailed at the other sites (Effiom et al., 2024; Holmgren et al., 2003; Neumann et al., 2008, 2010).
The early appearance of Pinus pollen in the Mkhuze sequence probably can be attributed to downward contamination of deposits during coring (pollen seepage). Since the sample is a swamp core, the moisture content of the sediment may have caused a seepage of the Pinus pollen. Hence, the early occurrence of Pinus is not likely to be due to the European spreading of the plant. Prehistoric pine expansion is reflected in other sequences like Lakes Sibaya, Eteza, and St. Lucia (Effiom et al., 2024; Neumann et al., 2008, 2010) when pine pollen dramatically increased since the arrival of European settlers (van Wilgen and Richardson, 2012) and when conditions became more like those in the present regime and where human influence is clearly indicated by other exotic and possibly cereal pollen.
Conclusion
Pollen analysis of a short Mkhuze swamp core (MK24-1) shed further light on the fluctuation of forest and development of grassland savanna, revealing a relatively warm environment with low moisture levels (humidity) for the last 1200 cal years BP at the time when Iron Age herders began to occupy the coastal area. Moisture-loving Podocarpus and cryptogam spores retreated, and fungal spores decreased as the environment became progressively warmer and drier with open savanna woodland from 1200 to 280 cal years BP, as suggested by prominent grass pollen ratios, culminating in a severe drought event between 280 and 200 cal years BP that resulted partly in the desiccation of the swamp and the occurrence of local fires. The pollen record shows that whereas all other species declined and some even disappeared, Amaranthaceae, charcoal, and charred cuticles experienced a comparable spike in number. During the last 200 years, a rather warm environment with high precipitation, wetland mosaic, and anthropogenic activities was evident, as shown by the presence of and rise in Typha, Pinus, Persicaria, and other wetland taxa. Similar patterns and vegetation characteristics were detected in a nearby pollen sequence from Lake St. Lucia for the last 2000 cal years BP (core MKD-1). Although similar, core MK24-1 differed from core MKD-1 environmentally, as MK24-1 experienced two periods of local fires, of which the earlier was likely caused by anthropogenic effects and or climate change. The apparent difference in the environmental changes was due to the location of the cores and associated sediment input. MKD-1 was collected offshore of the Mkhuze River delta, and MK24-1 in the swamp; hence, it is more susceptible to human activities and climate change, as this location is more vulnerable to changes in the quantity and quality of their water supply. Wider regional environmental comparison of the Mkhuze core aligns with other pollen sites in the region and δ18O and d13C studies at Cold Air Cave (Holmgren et al., 2003); however, it contradicts N15 findings at Marakabi (Chase et al., 2022), indicating that diverse environmental conditions prevailed in the region in the last c. 2000 years BP.
In conclusion, our findings show a rapid change from a forested environment with low moisture levels (humidity) to a warm and dry open woodland environment between 1100 and 200 cal years BP but became wetter during the last c. 200 years.
Footnotes
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The first author is grateful to the Paleontological Scientific Trust (PAST), Johannesburg, South Africa, for funding her PhD and GENUS: DSI-NRF Center of Excellence in Palaeosciences for supporting her postdoctoral research. She is also thankful to the German Federal Ministry of Education and Research (BMBF, Bonn, Germany) under the project “Tracing Human and Climate Impacts in South Africa” (TRACES, project number: 03F0798A) coordinated by Enno Schefuss and Mattias Zabel, for financing the laboratory expenses for this project.
Declaration of conflicting interests
The authors declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.