
Abstract
We present the first subfossil chironomid record from the Arabian Peninsula from a sediment core from Lake Gayal el Bazal, southeastern Yemen, providing insights into hydrological changes over the last ~1100 years. Shallow-water chironomid morphotypes, Dicrotendipes nervosus-type and Polypedilum nubeculosum-type, indicating low lake levels and productive habitats with abundant macrophyte cover, suggest two dry intervals between ca. 1575 and 1670 CE and 1770–1930 CE. These periods fall within the Little Ice Age (LIA), suggesting a link between lower global temperatures and weakened monsoonal activity. Since ca. 1930 CE, Tanytarsus cf. formosanus-type Bazal is dominant, indicating higher lake levels and lower productivity. This is supported by rising Ti/Al values, suggesting enhanced runoff and increased precipitation. A rise in Ceratopogonidae Dasyhelea-type during the last three decades suggests a decrease in lake size, likely reflecting anthropogenic water withdrawal since ca. 1989. Our findings support the notion of a dry LIA, affecting parts of Eastern Africa, southwestern Saudi Arabia, and extending to other latitudes such as the Tibetan Plateau. This points to large-scale weakening of the boreal summer monsoons and latitudinal southwards shifts of the Intertropical Convergence Zone. A pluvial period during the 17th century could be connected to a positive Indian Ocean Dipole event. Reconstructed increasing moisture during the pre-industrial era, which contrasts modern-day precipitation measurements, could be explained by seasonal variability with moisture increase during the early summer and autumn. Overall, our record emphasizes the interplay of climate variability and anthropogenic activities in shaping water resources and their sustainability in extremely dry low-latitude regions such as the Arabian Peninsula.
Introduction
Lake sediments can provide information on the surrounding terrestrial environment and climate history (Mills et al., 2017), including changes in precipitation, temperature, and human impacts (Halfman et al., 1992; Urrutia et al., 2010). In the Arabian Desert, the few numbers of lakes and semi-perennial streams (wadis) are the primary sources of freshwater, which are particularly vulnerable to environmental changes (Parth et al., 2023). Any changes in climate or human activities that affect these water resources can have significant impacts on the available quantity and quality of freshwater in the region. Therefore, paleolimnological studies can provide critical insights into the history of environmental changes in such a climatically sensitive area, and guide management practices for sustainable use of water and other resources in similar extreme environments worldwide.
Aquatic midge larvae such as Chironomidae (“non-biting”) and Ceratopogonidae (“biting”) are among the most abundant invertebrates in freshwater ecosystems around the world (Walker, 2001). Due to their rapid response to ecosystem changes by shifting their assemblages toward more tolerant species, they have been used for decades to trace changes in many environmental parameters, such as temperature, precipitation, nutrient status, or human impact (Brooks, 2006; Langdon et al., 2010). Their chitinous head capsules (HC) can be well preserved in lake sediments and may provide high-resolution climatic and environmental records, even for time spans of over hundreds of thousands of years (Rigterink et al., 2024). Previous work using subfossil chironomidae is mainly restricted to the Holarctic, especially regions which are rich in lakes and lacustrine systems, such as Europe, North-America, or Asia. Although recent work has also used chironomids to investigate tropical archives, especially Central and South America (Hamerlík et al., 2022), studies from sub-tropical regions, such as the Middle East and the Arabian Peninsula, have not yet employed chironomids. However, chironomids have shown extraordinary reactions to changes in moisture, especially in semi-arid to arid regions (Echeverría-Galindo et al., 2023; Eggermont et al., 2006) and are therefore especially suited to track fluctuations in precipitation and hydrological systems (Aubert et al., 2017, 2019; Eggermont et al., 2007).
Moreover, reliable records from these vulnerable regions (Figure 1a) provide valuable information on hydro-climate variability in relation with centennial- to millennial-scale latitudinal migration of the Intertropical Convergence Zone (ITCZ; Schneider et al., 2014), shifts in the position and strength of the tropical and sub-tropical monsoons, and reorganization of zonal circulation systems such as the IOD (Indian Ocean Dipole) and ENSO (El Nino-Southern Oscillation). Latitudinal migration of the ITCZ and the tropical rain belt is linked to the strength of the monsoon, which is the main driver of summer precipitation in northern Africa and Arabia (Fleitmann et al., 2003, 2007, 2022). Regional precipitation is also influenced by the IOD, the positive mode of which brings increased sea surface temperature and precipitation over much of the western tropical Indian Ocean region. We suggest that our chironomid record may reflect different components of the global climate system, which vary accordingly to the magnitude and strength of the ITCZ latitudinal migration through time.
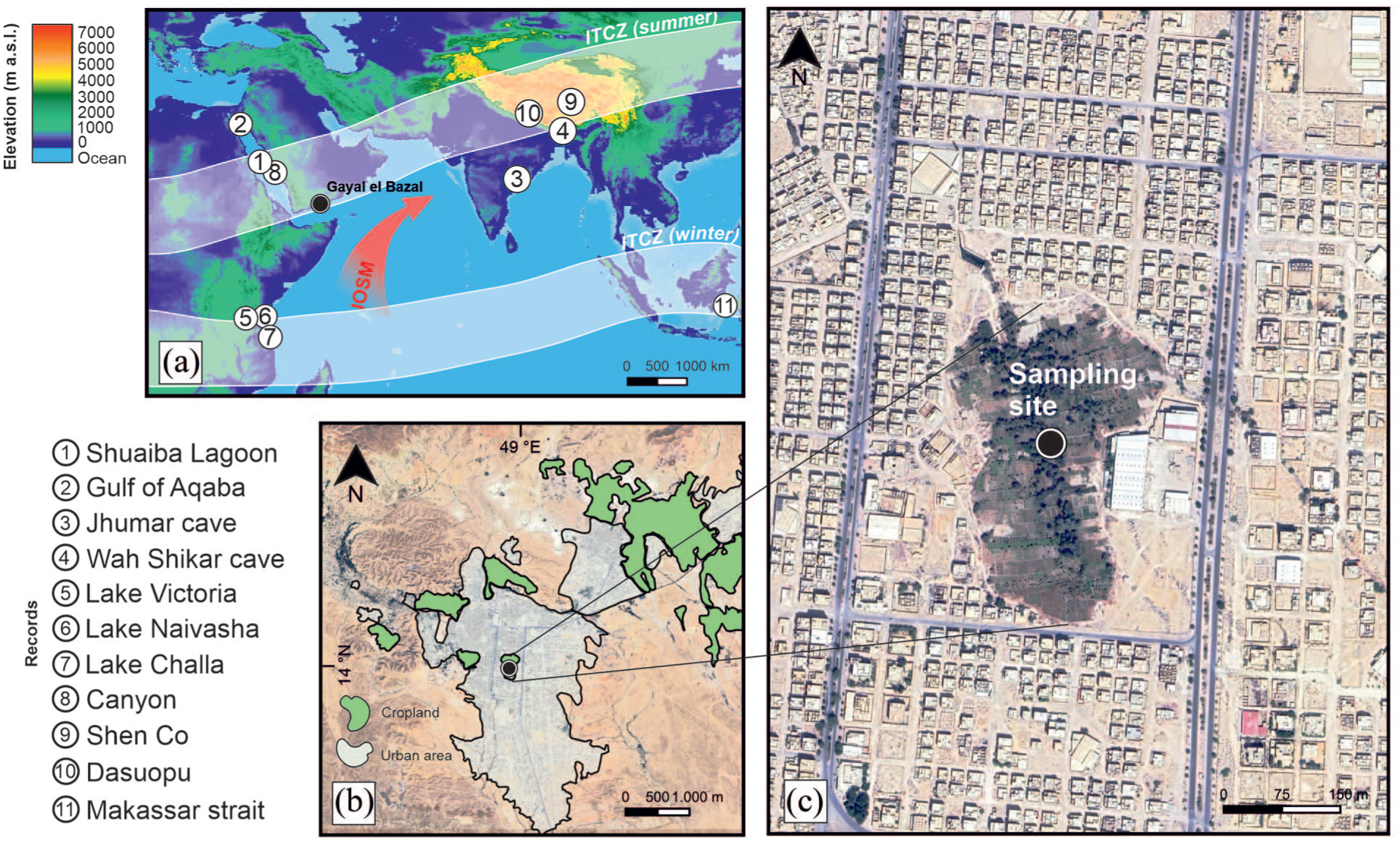
(a) Elevation map showing the extent of the Intertropical Convergence Zone (ITCZ) from 20°E to 1200°E during summer and winter (Lechleitner et al., 2017) with the Indian Ocean Summer Monsoon (IOSM) as the major atmospheric system providing rain to the Arabian Peninsula (Fleitmann et al., 2007; Petraglia et al., 2020). Circled numbers represent paleoclimate records displaying hydrological changes during the last millenium in the ITCZ region: (1) Abu-Zied and Bantan (2015), (2) Lamy et al. (2006), (3) & (4) Sinha et al. (2011), (5) & (6) & (7) Tierney et al. (2013), (8) Bryan et al. (2019), (9) Rigterink et al. (2022), (10) Thompson et al. 2000), (11) Oppo et al. (2009) lake (black dot); (c) Enlargement with the location of sediment core GBW from Lake Gayal el Bazal. Base maps of Gayal el Bazal were created with Google Satellite 2021, QGIS plugin. The World Digital Elevation Model (ETOPO1) was provided by the European Environment Agency (https://www.eea.europa.eu/data-and-maps/data/world-digital-elevationmodel-etopo5).
Well-dated, continuous paleo-records are available from the Nefud (Northern Arabia) and Rub al Khali (Southern Arabia) regions to the Hajar Mountains (Oman, South-Eastern Arabia) and the Arabian Sea coast (Woor et al., 2022). These records, including limnic (Parker et al., 2016; Parton et al., 2018), aeolian (Atkinson et al., 2011), fluvial (Berger et al., 2012), speleothems (Fleitmann et al., 2011), and marine sediment cores (Böll et al., 2015; Sirocko et al., 1993), provide valuable insights into the climatic history of the Arabian Peninsula from the Pleistocene to the Holocene. Paleoclimate records from Southern Arabia, particularly those covering the recent past and the last millennia, however, remain limited (Woor et al., 2022).
Two major temperature anomalies characterize the climatic variability of the last millennium: the Medieval Climate Anomaly (MCA, ca. 1000–1200 CE), and the so called “Little Ice Age” (LIA, ca. 1250–1860 CE). These intervals were characterized by global multi-decadal to centennial temperature changes (Neukom et al., 2022; Wanner et al., 2022) suggested to be caused by solar forcing (Steinhilber et al., 2009) and changes in tropical volcanic activity (Crowley, 2000; Mann et al., 1998) as well as land clearance and land use changes (Mann, 2002; Otto-Bliesner et al., 2016). The climate history of the Middle East and East Africa over the last millenium has been highly variable. Sedimentary and alluvial deposits from Saudi Arabia and Syria reveal a clear trend to warmer and wetter climate during the MCA and colder and drier climate during the LIA (Bantan et al., 2019; Kaniewski et al., 2011). The Horn of Africa however, seems to have been unaffected by temperature changes (Tierney et al., 2015). East Africa shows heterogeneous patterns with wet conditions in easternmost equatorial Africa and drought in the interior (Russell and Johnson, 2007; Tierney et al., 2015; Verschuren et al., 2000). In recent decades, a decrease in monsoonal rainfall is recorded (Guhathakurta and Rajeevan, 2008; Zhou et al., 2008), and precipitation decreased over the Arabian Peninsula (Kotwicki and Al Sulaimani, 2009). Although many studies have already addressed the effects of these fluctuations in mid- and low-latitude regions, temperature and moisture changes over the last millenium are regionally variable and are highly heterogenous in time and space, which makes further investigations necessary.
This study presents (1) the first subfossil chironomid record from the Arabian Peninsula, (2) relates shifts in the chironomid assemblages to precipitation changes over the last ~1100 years, and (3) compares the chironomid-inferred hydrological reconstruction from Lake Gayal el Bazal with existing records from the monsoonal region aiming to produce a complete and more detailed image of hydrological variability for the past 1100 years.
Site description
Lake Gayal el Bazal (14°46′15.7″N, 49°22′26.5″E, 85 m above sea level (a.s.l.)) today is an almost desiccated water body situated in Hadhramaut province, southern Yemen, located in the center of the Ghayl Ba Wazir town (Figure 1b). Its catchment area (24 km2) consists of Precambrian metamorphic rocks and Eocene limestone plateaus with elevations of up to 50 m a.s.l. Quaternary fluvial sediments were deposited in the wadis and surrounding plains (Parth et al., 2021). The lake fills a karstic sinkhole (Hehmeyer et al., 2002), and its surface area has greatly decreased reaching 0.1 km2 due to extensive anthropogenic groundwater abstraction (Figure 1c), which in turn led to a change in pH from 8 to 9. The remaining pond is mainly fed by short intermittent streams or local wadis and has a single small outlet. The regional ecotone comprises desert alluvial shrubs and grasses.
For the time period 1991–2020 CE, the annual precipitation in the region averages <100 mm/year (29.94 mm/year, available from worldclim.org, Supp. 1, available online), with summer temperatures reaching 38°C, and winter temperatures averaging 20°C (Al-Alimi et al., 2013, Climate Research Unit (CRU)). Modern hydrological conditions of the southern part of the Arabian Peninsula are mainly driven by seasonal migration of the ITCZ, linked to strengthening and weakening of the monsoons (Fleitmann and Matter, 2009). During summer, the interplay between the Red Sea trough (Baseer et al., 2019) as well as the Indian Summer Monsoon (ISM; Neugebauer et al., 2022) leads to two rainy seasons, one lasting from March to April and the second one from June to August (Baseer et al., 2019; Fleitmann and Matter, 2009).
Materials and methods
Sampling, chronology and sedimentology of sediment core GWB01
Three sediment cores were retrieved in December 2001 from paleo-lake Gayal el Bazal using a piston corer (Parth et al., 2021). A composite core GBW (326 cm) was generated based on sedimentological investigations. The chronological framework of the GBW composite core was established using five AMS radiocarbon dates obtained from terrestrial plant remains (wood fragments, and twigs) recovered at depths of 62, 112, 157, 242, and 325 cm (Supp. 2, available online). The uppermost section of the core was assumed to represent the modern age (or coring year), and this assumption was incorporated into a regression model to constrain the age of the upper layers (Stager et al., 2009; Yuan et al., 2006; Zhang and Mischke, 2009). All radiocarbon dates were calibrated using the most recent IntCal20 calibration curve (Reimer et al., 2020). Sediments of the GBW core mostly consist of calcareous silt with minor occurrences of sand and clay. Details of the sedimentology and chronology are given in Parth et al. (2021).
Chironomidae analysis
Sediment samples were prepared according to Walker (1987) with KOH solution (10%) and heated for 20 min at 80–90°C. The prepared sediment was passed through a 100 μm-mesh nylon sieve. Head capsules were handpicked under a stereo-zoom microscope with 32- and 16-fold magnification. Identification of Chironomidae and Ceratopogonidae taxa was done using a Zeiss Axio Imager.A2 microscope following Andersen et al. (2013), Brooks et al. (2007), Eggermont and Verschuren (2004), and Walker (2001). Head capsules were identified to morphotype and generic levels. Some head capsules could only be identified down to subfamily/tribe level (e.g. Chironomini indet. /Tanytarsini indet.). Chironomidae and Ceratopogonidae head capsule concentrations are presented as number of HC per dry mass sediment (HC g−1).
Statistical analysis
Graphical representation of Chironomidae and Ceratopogonidae abundances was done with the C2 program (version 1.7.7, Juggins, 2014). Rarefaction analysis was used to check the appropriateness of the number of head capsules per sample (Engels et al., 2020). For multivariate analyses, only samples with more than 32 head capsules were used. For this purpose, nearby samples were also combined. Chironomid morphotype abundances <2 % were eliminated from multivariate statistics. Prior to ordination and cluster analyses, chironomid abundances were square-root transformed. Stratigraphic zones were identified using constrained hierarchical cluster analysis with Bray–Curtis distance and the CONISS linkage method (Grimm, 1987). A broken-stick model (Bennett, 1996) was used to identify the statistical significance of the clusters. A Detrended correspondence analysis (DCA) revealed an DC axis 1 length of 1.3 standard deviations, therefore a Principal component analysis (PCA) was used to present changes in species composition (Ter Braak, 1987). We identified the influence of the environmental variables on the chironomid morphotype distribution with a Canonical Correspondence Analysis (CCA; Ter Braak, 1987). Variance inflation factors (VIF) were calculated to identify environmental variables with high correlation (Miles, 2014). We used variance analysis (ANOVA; 999 permutations) to test the significance of each environmental variable. All analyses were performed using vegan-package and rioja-package from R software (R Core Team, 2021).
Results
Chironomidae assemblages of sediment core GBW from Gayal el Bazal
In total, 85 samples were analyzed for Chironomidae and Ceratopogonidae head capsules (Figure 2, Supp. 3, available online). On average, 47 chironomid head capsules were counted per sample (range: 0–258 HC). A minimum number of head capsules was set at 32 HC per sample for the environmental reconstruction based on results of the rarefaction analysis (Supp. 4, available online). This shows that most of the samples reached their asymptote, suggesting that the potential number of species was found in an individual sample. A maximum number of 10 chironomid morphotypes was identified (Supp. 5, available online), out of which four were considered as non-rare taxa (Hill’s N2>5) and six as rare taxa (Hill’s N2<5). Paratendipes nudisquama-type was also considered as a principle chironomid morphotype because of its abundances of up to 12% of the total assemblage. This leads to a number of five principle chironomid morphotypes, which were included in the statistical analysis (Supp. 6, available online). Three significant biostratigraphical zones were identified using Constrained Clustering (Supp. 7, available online) and changes of CCA of chironomid assemblages. For Ceratopogonidae head capsules two different morphotypes, Bezzia-type and Dasyhelea-type, could be distinguished (Supp. 6, available online).
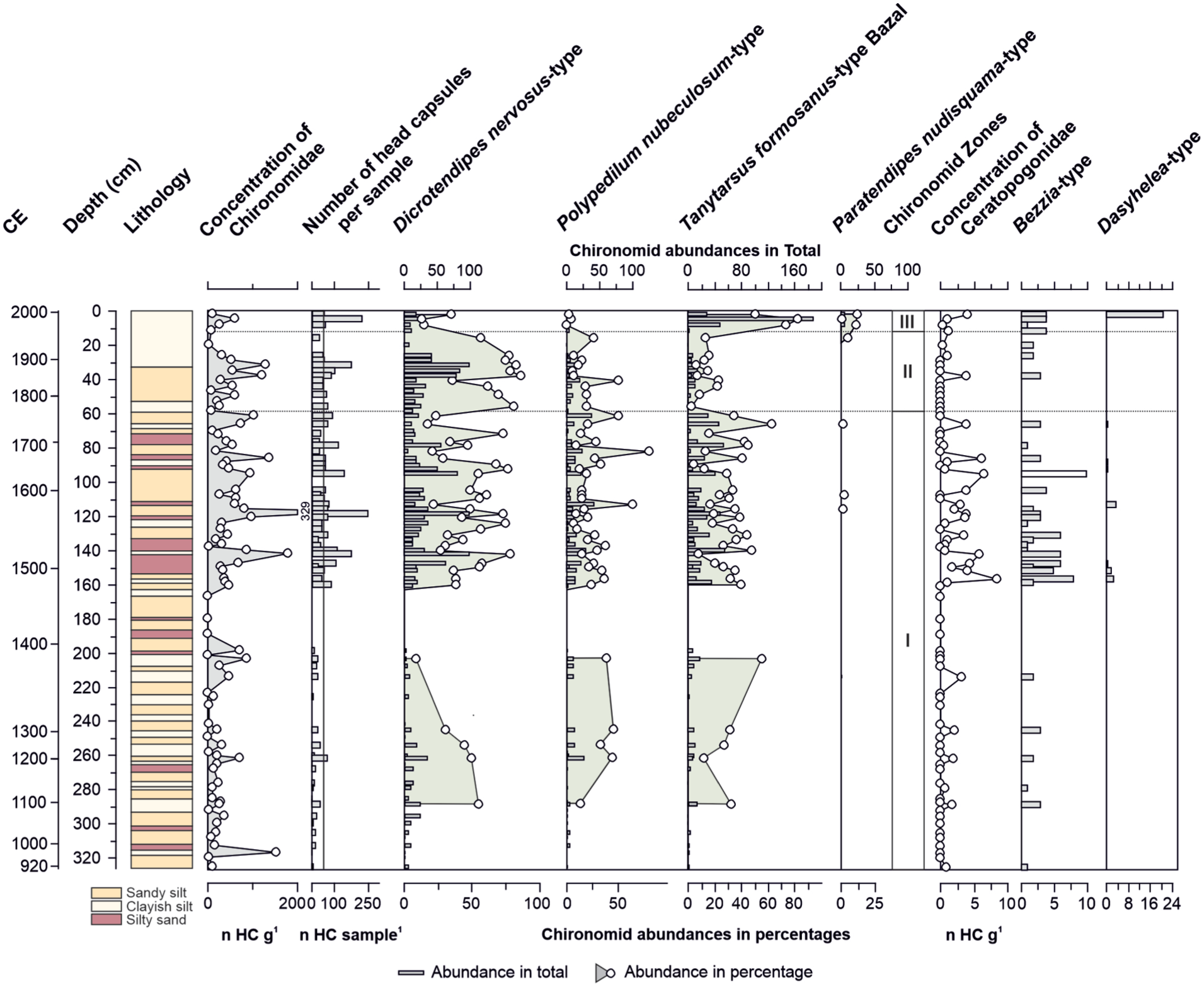
Lithology of sediment core GBW, concentration and number of chironomid head capsules per sample, total (horizontal bars) and relative abundances (circles) of principle chironomid morphotypes, as well as concentration and number of head capsules of Ceratopogonidae morphotypes Bezzia and Dasyhelea. Chironomid zones I–III are based on results of cluster analysis.
Zone I (326 – 61.5 cm, 914 – 1750 CE)
The beginning of this zone (326–167 cm) is characterized by a low head capsule concentration of 14 HC g−1 on average. The minimum number of 32 head capsules was reached in five samples (289, 262, 254, 245, and 204 cm) only. Dicrotendipes nervosus-type is dominating the bottom part of the zone until 254 cm, with abundances of up to 56%. Polypedilum nubeculosum-type and Tanytarsus cf. formosanus-type Bazal reach abundances of 11–37% and 13–33%, respectively. Until 204 cm, the abundance of Tanytarsus cf. formosanus-type Bazal increases up to 56%, while Dicrotendipes nervosus-type decreases down to 9%. Polypedilum nubeculosum-type reaches abundances up to 38%. From 189 to 167 cm, chironomid head capsules were absent. From 160 cm upwards, the chironomid concentration increases (10–174 HC g−1). The assemblages are dominated by Dicrotendipes nervosus-type (17–80%), together with Polypedilum nubeculosum-type (6–66%) and Tanytarsus cf. formosanus-type Bazal (5–63%). Paratendipes nervosus-type reaches abundances up to 3%. Only a few (<5 HC) Ceratopogonidae head capsules were found below 160 cm, belonging to the Bezzia-type. Concentration of Ceratopogonidae increases above 160 cm (>5 HC g−1), the Bezzia-type is the dominant morphotype, and a few Dasyhelea-type appear.
Zone II (56–16 cm, 1750–1930 CE)
Head capsule concentration fluctuates between 3 and 92 HC g−1. The dominance of Dicrotendipes nervosus-type (35–87%) and Polypedilum nubeculosum-type (5–52%) increases, while the abundances of Tanytarsus cf. formosanus-type Bazal (3–35%) decrease in Zone II. Other chironomid morphotypes, such as Chironomus anthracinus-type and Chironomus plumosus-type, reach abundances of up to 5%. The concentration of Ceratopogonidae decreases (<5 HC g−1), and are mainly represented by the Bezzia-type.
Zone III ((8–2 cm, 1930–1989 CE)
Head capsule concentration decreases (9–52 HC g−1). Abundances of Dicrotendipes nervosus-type (13–35%) and Polypedilum nubeculosum-type (0–4%) decline. The abundances of Tanytarsus cf. formosanus-type Bazal increase and reach up to 82%. Paratendipes nudisquama-type appears in this zone as well and shows increasing abundances of up to 12%. The concentration of Ceratopogonidae remains low (<5 HC g−1), the dominance of the Bezzia-type is replaced by Dasyhelea-type in the uppermost sample (2 cm/1989 CE).
Relationship between chironomid assemblages and environmental variables
The principal component analysis demonstrates that axes 1 and 2 explain 73% and 26% of the variation in the chironomid assemblages, respectively (Supp. 8, available online). CCA (Figure 3) reveals that the included environmental variables (TOC, Ca/(Ti + Al+Fe), Sr/Rb, Ti/Al, and quartz (%), Supp. 9, available online) explain 21% of the variation of the chironomid assemblages (CCA1: 19 %, CCA2: 2 %, Supp. 10, available online). Samples belonging to Zones I and II are predominantly associated with the positive side of CCA1, while samples of Zone III are associated with the negative side of CCA1. Verification inflation factors suggest that TOC, Ca/(Ti + Al + Fe), Sr/Rb, Ti/Al, and quartz (ViF < 5) are non-correlated environmental variables (Supp. 11, available online), which are therefore independent predictors. ANOVA identified Ti/Al (8 variance) as a significant variable (Supp. 11, available online). Dicrotendipes nervosus-type and Polypedilum nubeculosum-type are associated with the positive side of CCA1, and are predominantly influenced by the increase in TOC, Sr/Rb, and Ca/(Ti + Al + Fe). On the other hand, Paratendipes nudisquama-type and Tanytarsus cf. formosanus-type Bazal are related to the negative side of CCA1, mainly explained by the increases in Ti/Al ratios and quartz.
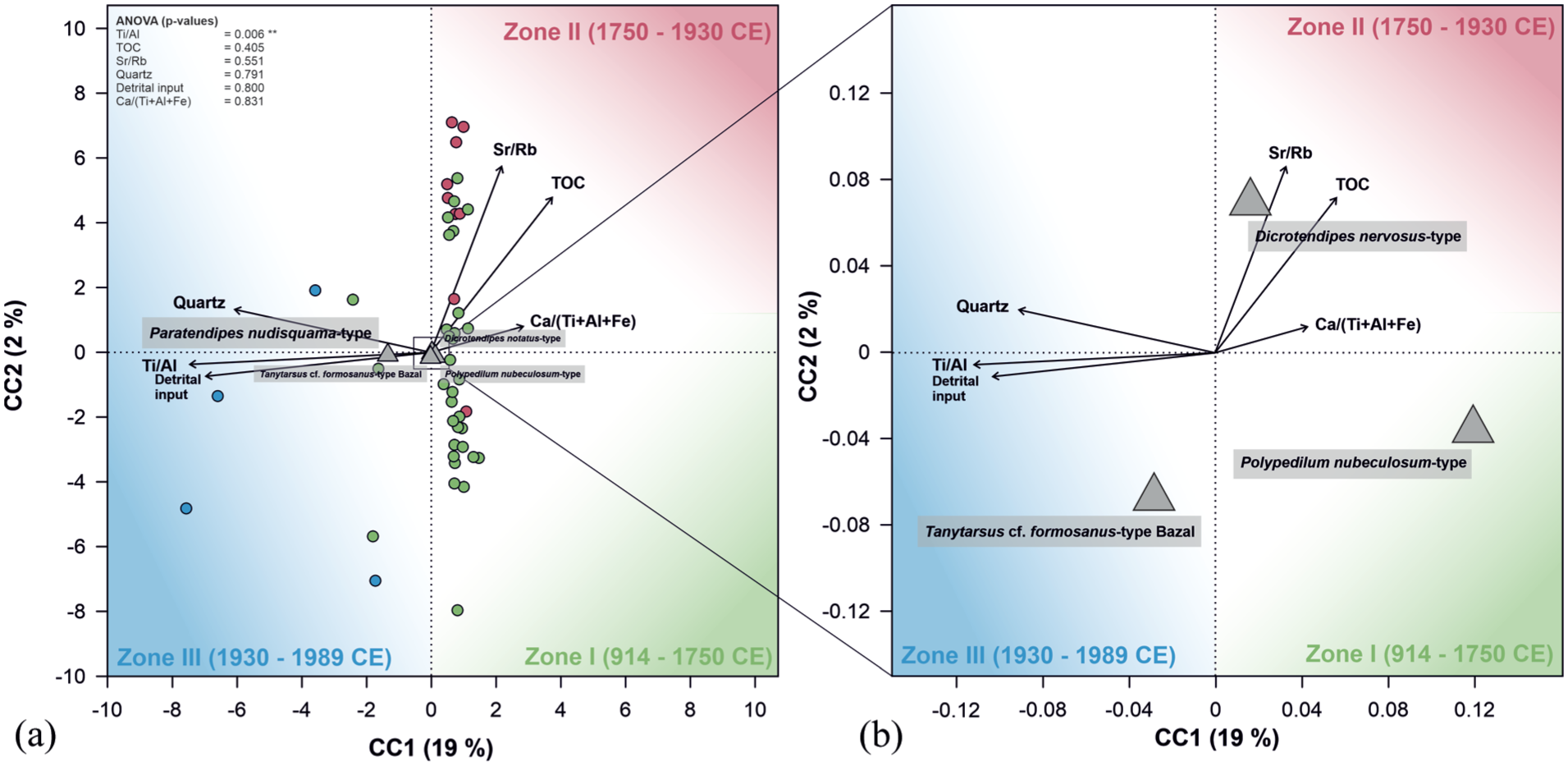
(a) Relationships between chironomid morphotypes and independent environmental variables (TOC, Ca/(Ti + Al + Fe), Sr/Rb, Ti/Al, and quartz abundance) displayed with Canonical Correlation Analysis (CCA). Environmental parameters derived from Parth et al. (2021). Dominant chironomid morphotypes are indicated by gray triangles and samples are represented by colored points (green = Zone I, 914–1750 CE; red = Zone II, 1750–1930 CE; blue = Zone III, 1930–1989 CE; colored shapes represent direction of sample scores). Zone II is characterized by a dominance of Dicrotendipes nervosus-type and increased TOC, Ca/(Ti + Al + Fe) and Sr/Rb ratio, while Tanytarsus formosanus-type Bazal dominates Zone III, characterized by increased Ti/Al values and detrital input. Note that (b) is an enlarged and more detailed version of (a) providing a better overview of the dominant taxa.
Discussion
As far as we know, the subfossil chironomid record from Lake Gayal el Bazal is the first available from Yemen. Considering the extreme aridity, only very few paleolimnological studies are available from the Arabian Peninsula, and the fewest use chironomids (Reeves and Epler, 2016). Some descriptions of adult, winged chironomidae taxa exist, for example, from the United Arab Emirates, Kuwait, Oman, and Saudi Arabia (Giłka, 2009). To our knowledge, identification guides for subfossil chironomid head capsules from the Arabian Peninsula are not yet available. The geographically closest descriptions of subfossil chironomid morphotypes are from East Africa (Eggermont et al., 2006) and were thus considered in this study. Middle Eastern and East African records of subfossil chironomidae are available from, for example, Iran (Aubert et al., 2017, 2019), Kenya (Verschuren et al., 2000), and Chad (Eggermont et al., 2008), mainly using the well-documented identification literature and ecological descriptions from Europe and Africa (Brooks et al., 2007; Verschuren, 1997). The morphology of the Gayal el Bazal chironomid morphotypes are closest to the morphotypes described by Brooks et al. (2007). Dicrotendipes nervosus-type, Polypedilum nubeculosum-type and Paratendipes nudisquama-type are well known morphotypes from Europe (Brooks et al., 2007). Morphological characteristics, such as mandibles, mentum, and antennal pedestal, were easy to assign to the known morphotypes. Only Tanytarsus, found in the sediments from Gayal el Bazal, could not be assigned to the Tanytarsus morphotypes described by Brooks et al. (2007), because of differences in shape and size of the mentum and antennal pedestals as well as the number of teeth of the mandibles. Nevertheless, Tanytarsus specimens from Gayal el Bazal resemble Tanytarsus formosanus Kieffer (Eggermont and Verschuren, 2004), whereby the mandible of the morphotype from Gayal el Bazal is missing the additional tooth at the base of the dorsal tooth. For this reason, the Tanytarsus formosanus morphotype from Gayal el Bazal is named Tanytarsus cf. formosanus-type Bazal hereafter. The identification of the new morphotype Tanytarsus cf. formosanus-type Bazal underscores the potential of chironomid research to expand regional taxonomic knowledge and ecological understanding. Although we use these studies to guide our paleoenvironmental interpretation of the Gayal el Bazal chironomid morphotypes, it cannot be ruled out that the ecological requirements differ between these highly divers ecosystems in Europe, Africa, and the Arabian Peninsula.
Due to the high aridity and limited seasonal precipitation in the study area, this extreme habitat may be one reason for the relatively low chironomid diversity as shown by a Shannon index of <1, which is consistent with findings from similar arid to semi-arid regions worldwide (Lin et al., 2017; Rigterink et al., 2022; Zhang et al., 2007). The chironomid taxa from Gayal el Bazal are defined as affiliated with shallow, meso-eutrophic, temperate waters, and inhabiting mainly lentic systems. In arid regions, water is the decisive limiting factor for the growth and survival of plants and animals. For this reason, lake level and salinity might be the most important environmental factors shaping chironomid communities of lakes from arid and semi-arid regions (Eggermont et al., 2006, 2008; Laug et al., 2020; Rigterink et al., 2022; Verschuren et al., 2000; Zhang et al., 2007), such as the Arabian Peninsula, and can give in turn assumptions about past moisture availability and precipitation, especially in closed lake systems (Chen et al., 2009; Fritz, 2008). Other influencing factors, such as temperature or lake bottom oxygen availability, play a subordinate role in Lake Gayal el Bazal, because the lake is shallow, and lacking stratification (Holgerson et al., 2022; Scheffer, 1997). Results of the performed CCA (Figure 3) associate shifts in the main chironomid morphotypes to changes in moisture and productivity-driven environmental parameters, such as precipitation, salinity, and nutrient loads. Therefore, shifts in chironomid morphotypes were used to identify changes in productivity and moisture availability during the last ~1100 years, more specifically shifts between drier and wetter intervals.
Environmental reconstruction of Lake Gayal el Bazal based on chironomids
Due to limited sediment sample size available, abundance of head capsules at the bottom of the core was low, restricting ecological interpretations for the period before 1475 CE. The Little Ice Age is mainly dominated by littoral taxa, Dicrotendipes nervosus-type and Polypedilum nubeculosum-type, suggesting a shallow lake environment and well-established macrophyte vegetation (Brooks et al., 2007; Moller Pillot, 2009). Occurrences of Ceratopogonidae, mostly Bezzia-type, indicate a shallow, warm water environment (Ilyashuk et al., 2005), which is supported by a higher availability of organic matter indicated by increased TOC content. Furthermore, increased Sr/Rb and Ca/(Al + Ti + Fe) ratios (Parth et al., 2021) suggest negative precipitation/evaporation ratio and high productivity within the lake (Kasper et al., 2021; Li et al., 2008; Zhisheng et al., 2001) underlining drier conditions. As a deposit-feeder, Dicrotendipes is associated with fine sediments that may be associated with soil erosion, which also can affect the trophic status of the lake (Zhang et al., 2013). We suggest that during the periods dominated by the Dicrotendipes, lake levels decreased and Lake Gayal el Bazal received more eroded materials. While Dicrotendipes nervosus-type is associated with macrophyte vegetation (Langdon et al., 2010), Polypedilum nubeculosum-type rather represents clear waters. For this reason, periods dominated by Dicrotendipes nervosus-type, for example around 1520 CE, during 1555–1560 CE, around 1660 CE, and especially from 1770 to 1900 CE, point to years with higher productivity, as confirmed by increased TOC values, for which we therefore postulate the lowest lake levels of the record and maximum drought.
Major trends suggested by the statistical analysis (PCA, CCA) are driven by Tanytarsus cf. formosanus-type Bazal. For Eurasia, chironomid morphotypes belonging to the genus Tanytarsus are mostly cold-stenothermic and are known to inhabit the profundal parts of standing waters with low productivity (Brooks et al., 2007; Langdon et al., 2010). Tanytarsus formosanus is common in lentic and lotic water bodies from Europe and across Africa, Australia to South- and East Asia (Giłka, 2009) and tends to be tolerant to higher salinities but seems to be intolerant to daily oxygen fluctuations (Ekrem, 2001a, 2001b). In African lakes, Tanytarsus formosanus is adapted to lower salinities (<1000 µS cm−1; Eggermont et al., 2006), and coarser grain sizes (Eggermont et al., 2007), underlining its use as an indicator of a higher lake level and low productivity. In the Gayal el Bazal record, the chironomid morphotype Tanytarsus cf. formosanus-type Bazal is associated with high runoff (Ti/Al ratios) and elevated Quartz content, as explained by the CCA (Figure 3). Wetter episodes, characterized by a rise of Tanytarsus cf. formosanus-type Bazal, are evident during the LIA, especially from 1523 to 1542 CE and 1681–1750 CE, and after 1900 CE, respectively. These periods are marked by an increase in Ti/Al ratios and quartz content, indicating enhanced precipitation, causing stronger runoff from the catchment (Neugebauer et al., 2022).
Paratendipes nudisquama-type, which is known from the Near East and Northern Africa (Płóciennik et al., 2010), tends to be abundant in peaty moorland pools and bogs, and also appears in streams (Khazenzi et al., 2022; Moller Pillot, 2009). CCA reveals that P. nudisquama-type is associated with higher quartz content in Lake Gayal el Bazal (Figure 3). We suggest that increasing abundances of head capsules belonging to Paratendipes nudisquama-type were introduced to the lake via enhanced runoff by intermittent streams or flash floods, which play an important role in the hydrological conditions of desert areas. In mid-low latitude drylands, their recurrence rates tend to increase with increased monsoonal activity during warm periods.
After Hadramawt province had become independent from the United Kingdom in 1967 CE, the region experienced socioeconomic growth. This is well expressed by the population density in the region that had doubled from ca. 14,000 in 1970 CE to almost 30,000 by 1990 CE (Supp. 11, available online, Goldewijk et al., 2017). The agricultural sector was modernized, which increased groundwater abstraction from Ghayl Ba Wazir (Hehmeyer et al., 2002). In addition, since the 1980s, the wells of Ghayl Ba Wazir have been used to supply drinking water to the provincial capital, al-Mukalla. Groundwater abstraction was further intensified after 1998 CE (Hehmeyer et al., 2002). This led to a drop of the water table (Rybakov et al., 2009), which is likely to have lowered the level of Lake Gayal el Bazal.
Larvae of Ceratopogonidae Dasyhelea-type are found in aquatic and semi-aquatic habitats such as mud, wet soil, ponds, streams, salt marshes etc. (Szadziewski et al., 2011; Waugh and Wirth, 1976). As non-swimmers (Mullen and Hribar, 1988), they are dependent on the presence of blanket algae between which they live. Increasing abundances of Ceratopogonidae Dasyhelea-type after 1989 CE could therefore be interpreted to have resulted from extended wetlands being established after lake level draw-down. Because there is no increase in TOC and Sr/Rb ratio in the uppermost sample, we assume that productivity plays no role in this interpretation. An increase in the Ca/(Al + Ti + Fe) ratio supports the interpretation of lower lake levels, suggesting a decrease in precipitation in the uppermost samples.
Climate forcing and regional teleconnections
The record from Gayal el Bazal spans across several distinct climate fluctuations, including the end of the Medieval Climate Anomaly, the Little Ice Age, and the post-industrial era (Parth et al., 2021, 2023). As revealed by the environmental reconstruction of the paleolake, the chironomid assemblages from Gayal el Bazal correspond closely to wet and dry intervals. For this reason, we used the chironomid PC1 (Figure 4) to identify key hydrological events such as droughts or pluvial phases.
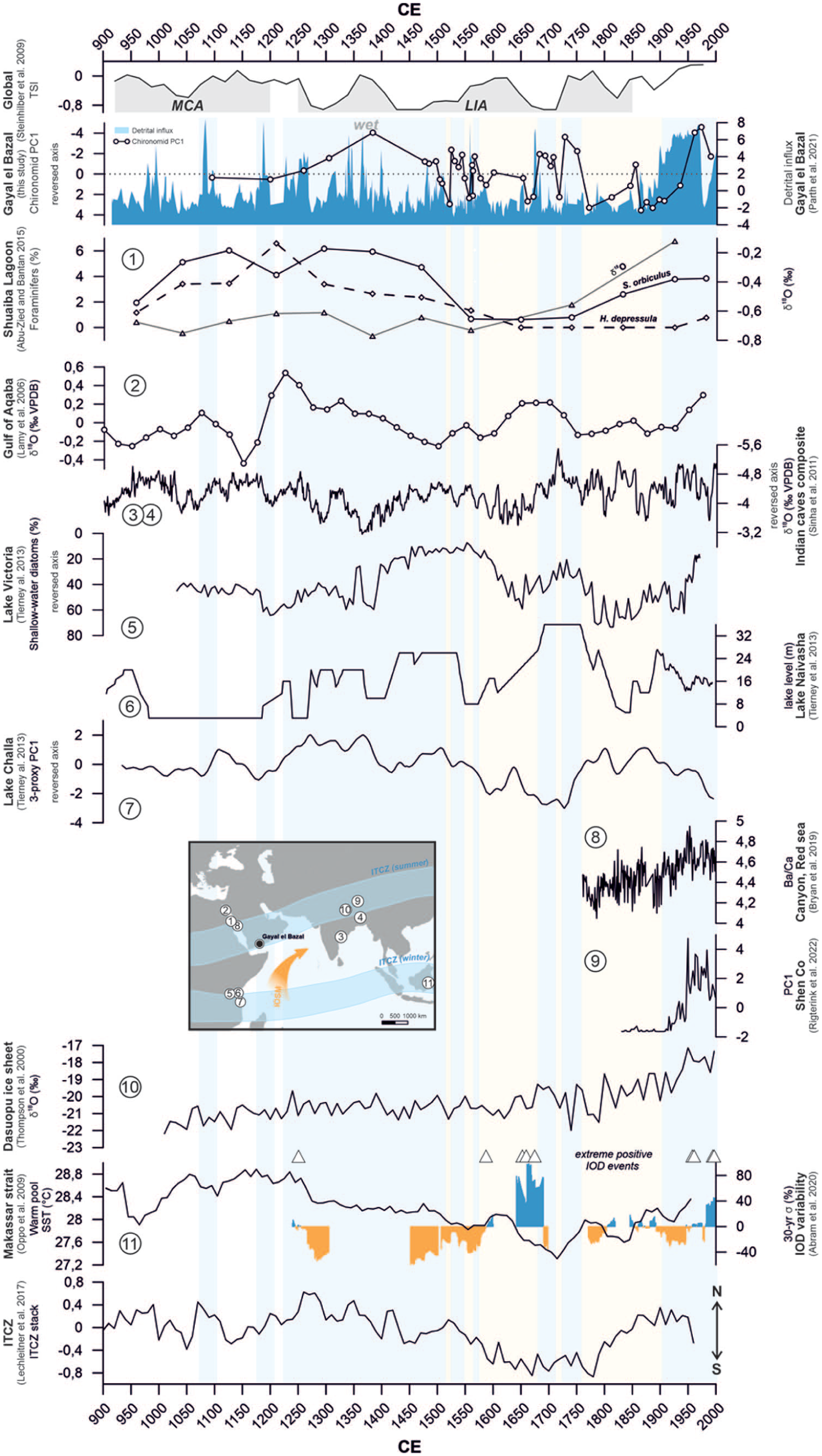
Total solar irradiance (TSI, Steinhilber et al. (2009) marking increased solar activity during the MCA (1000–1200 CE) and decreasing during the LIA (1250–1860 CE). Comparison of PC1 (chironomid assemblages with dominant chironomid morphotypes), head capsule concentration (HC g-1) and detrital influx to Lake Gayal el Bazal as indicators of wet and dry episodes (Parth et al., 2021) during the last millenium compared to paleoclimate records from the summer ITCZ region. Note that positive PC1 values denote a decrease in lake level (dry phases) and negative PC1 values mark lake level increase (wet phases). The selection of paleoclimate records (circled numbers refer to Figure 1a) is explained in the discussion. Warm pool sea surface temperatures from the Makassar strait (Oppo et al., 2009), Indian Ocean Dipole (IOD) variability (vertical blue/orange bars denote positive/negative IOD events, Abram et al., 2020) as well as extreme positive Indian Ocean Dipole events (white triangles, Abram et al., 2020), and the ITZC variability (Lechleitner et al. 2017) were added to put our results into a global context. Our results show dry periods, especially between ca. 1575 and 1670 CE and ca. 1770–1900 CE, respectively, during the Little Ice Age (LIA). A pluvial event was recorded between 1681 and 1749 CE. An increase in precipitation has been detected since ca. 1930 CE. Paleoclimate datasets available from the National Oceanic and Atmospheric Administration (https://www.ncdc.noaa.gov/cdo-web/).
During the LIA, changes in the thermohaline circulation in the North Atlantic resulted in a southward shift of the westerlies (Wanner et al., 2011), and colder temperatures, which are best expressed in records from the Northern Hemisphere (Bradley et al., 2003; Briffa and Osborn, 2002; Trouet et al., 2009; Wassenburg et al., 2013). In the Lake Gayal el Bazal record, the Little Ice Age is archived as a dry period. This is consistent with several other hydro-climate records from the summer ITCZ region (Figure 4), spanning from the western Arabian Peninsula to India, a vast region also directly affected by the monsoonal system (Abu-Zied and Bantan, 2015; Bryan et al., 2019; Rigterink et al., 2022; Sinha et al., 2011; Thompson et al., 2000). A dry phase from ca. 1575 to 1670 CE is also shown by abundances and δ18O -enrichment of foraminifers from Shuaiba Lagoon, Red Sea, suggesting consistent drought during the 16th century (Abu-Zied and Bantan, 2015). A second dry phase (1770–1930 CE) is also found in 18O-enrichment displayed by a record from the Gulf of Aqaba (Lamy et al., 2006). Drought was also recorded by low-latitude archives elsewhere, such as in Indian cave records (Sinha et al., 2011) and lakes in central equatorial Africa (Russell and Johnson, 2007). During warm phases such as the MCA, the monsoon-season was characterized by increased rainfall, which accompanied a northward shift of the ITCZ on the Arabian Peninsula (Fleitmann et al., 2004). The most plausible explanation for a dry LIA in our record is a multi-centennial weakening of the Indian summer monsoon (Schneider et al., 2014) caused by a southward shift of the ITCZ (Lechleitner et al., 2017; Sachs et al., 2009).
A wet period detected in our record between 1681 and 1749 CE coincides with a pluvial event from 1680 to 1765 CE, archived in proxy records from coastal lakes in East Africa (Tierney et al., 2013). This indicates that the region was not continuously exposed to dry conditions during the LIA, but that potentially local climate drivers have an influence on the climatic conditions of the region. The Indian Ocean Dipole, a natural climate phenomenon of the Indian Ocean, is characterized by a sea surface temperature (SST) gradient in the Indian Ocean (Saji et al., 1999). Positive IOD events, characterized by cool/warm SSTs in the eastern/western Indian Ocean, initiated by enhanced ocean upwelling in the east, and strengthening of the winds along the equator, lead to an increase in atmospheric convection over the western Indian Ocean associated with a weakening of the Walker cell (Webster et al., 1999). This leads to increased rainfall in East Africa (Webster et al., 1999). Abram et al. (2020) reconstructed the IOD variability during the last millenium using coral δ18O records from the Mentawai Island (Figure 4). They detected different extreme positive IOD events, especially during the 17th century (Tejavath et al., 2024) and the pre-industrial era. The pluvial phase, which we detected during the 17th century, directly follows an extreme positive IOD event in 1675 CE (Abram et al., 2020). This supports an impact of changing Indian-Ocean sea surface temperatures on the hydrological conditions of the Arabian Peninsula.
The current warm period (ca. 1850–today) is marked by the modern, human-induced climate warming, where temperatures are rapidly approaching levels that exceed any other warm interval since the Pliocene (IPCC, 2021). Since ca. 1850 CE, human-induced global warming has been leading to ecosystem changes worldwide (IPCC, 2021). Deforestation and intensified farming have resulted in a decline in biodiversity, increased surface erosion, and degradation of freshwater supplies. Although the dating of this section of the Gayal el Bazal record is somewhat imprecise (Parth et al., 2021), our record is in line with other archives that indicate wetter conditions, for example the increase in Ba/Ca values of dust measured in corals since the 1950s in the Red Sea (Bryan et al., 2019), and chironomid records from the Tibetan Plateau (Rigterink et al., 2022). Moreover, δ18O data from the Dasuopu ice sheet in the Himalaya (Thompson et al., 2000) show an increase in local wetness, which corresponds to our interpretations from southern Yemen. This, however, contrasts with modern-day precipitation data measurements from the region (Almazroui et al., 2012; Almazroui and Saeed, 2020). Almazroui et al. (2012) detected increasing wetness on the Arabian Peninsula from 1979 to 1993 CE and a decreasing trend from 1994 to 2009 CE. The contribution of extreme weather events to annual precipitation increased between 1970 and 1999 CE (Almazroui and Saeed, 2020). As our record only goes back to 1989 CE, it is unlikely that this decreasing trend can be captured by our record. Climate data for the Republic of Yemen (CRU, Supp. 1, available online) show an increase in annual precipitation during the 1950s, with a peak around 1963 CE and a decreasing trend during the last decades. However, during early summer (May–Jun) and autumn (Sep–Nov), precipitation increased from the interval 1960–1990 CE to the interval 1991–2020 CE. Abundances of chironomid taxa are controlled by various factors such as hatching rate, survivability of eggs, proportions of eggs that successfully hatch, development rates, timing of emergence, swarming, and more. These factors are in turn dependent on variables that vary seasonally, for example, temperature, light intensity, or pH (Tokeshi, 1995). This suggests that our data rather reflects a seasonal increase in wetness than an overall trend. The recent increase in humidity is probably associated with the strengthening of the ISM during the last decades as a consequence of anthropogenic warming (Bryan et al., 2019).
Conclusions
Chironomidae analysis was carried out for the first time in Yemen, using a sedimentary record from Lake Gayal el Bazal that documents hydrological variability during the last 1100 years. Dry phases (ca. 1575–1670 CE and 1770–1930 CE) are characterized by a dominance of littoral chironomid morphotypes like Dicrotendipes nervosus-type and Polypedilum nubeculosum-type, in combination with increasing TOC contents and Sr/Rb ratios, indicating a productive environment with lower moisture availability. Moister phases (ca. 1520–1565, 1680–1750 CE, and ca. 1930 CE–today) favored chironomid morphotypes such as Tanytarsus cf. formosanus-type Bazal, which are related to lower productivity and higher runoff. This is supported by an increase in Ti/Al ratios and quartz content, suggesting higher runoff caused by enhanced precipitation. Higher abundances of the Ceratopogonidae Dasyhelea-type suggest that anthropogenic influence started in 1989 CE, when groundwater abstraction caused semi-terrestrial habitats to expand. The chironomid results from Gayal el Bazal are consistent with results from several other paleoclimate records from surrounding regions, spanning from East Africa over western Saudi Arabia to the Tibetan Plateau, indicating severe droughts during the Little Ice Age, associated with orbitally-paced latitudinal southward shifts of the ITCZ. A pluvial phase from 1681 to 1749 CE could be connected to a positive Indian Ocean Dipole event, causing an upward lift of the Walker circulation over the western Indian Ocean and enhanced rainfall. Contradicting results regarding the pre-industrial era suggest that our chironomid record primarily reflects seasonal wetness increases rather than an annual trend. Our work highlights the importance of using aquatic bioindicators in arid regions, given their sensitivity to various influences, including climate-driven changes and human impact.
Supplemental Material
sj-docx-1-hol-10.1177_09596836251366210 – Supplemental material for Hydroclimate variability and human influence in southern Yemen over the past 1100 years: Insights from subfossil midge (Diptera: Chironomidae, Ceratopogonidae) assemblages from paleolake Gayal el Bazal
Supplemental material, sj-docx-1-hol-10.1177_09596836251366210 for Hydroclimate variability and human influence in southern Yemen over the past 1100 years: Insights from subfossil midge (Diptera: Chironomidae, Ceratopogonidae) assemblages from paleolake Gayal el Bazal by Sonja Rigterink, Shah Parth, Nicolas Waldmann, James Russell, Julia Schmidt and Antje Schwalb in The Holocene
Footnotes
Acknowledgements
We thank the Continental Scientific Drilling (CSD) Facility, University of Minnesota, for providing sediment samples. We thank Douglas Schnurrenberger and Juris Zarins, who performed fieldwork in Yemen in 2001. The authors would like to acknowledge Dr. Nimer Taha for his assistance with chironomid analysis. We would like to thank the anonymous reviewers for their helpful comments.
Author contributions
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: We thank the University of Haifa Bloom School of Graduate Studies for granting a research stay to Sonja Rigterink under the Sandwich Scholarship umbrella.
Data availability statement
The data supporting the findings of this study are available in the Supplementary material and upon reasonable request from the corresponding author.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.