
Abstract
Two major debates frame Arctic prehistory: the emergence of specialized maritime economies that support sedentism and growing socio-political complexity, and the role of these economies in the transition from Paleo-Inuit to Neo-Inuit traditions. This shift, originating in the Bering Strait, led to Thule Inuit dispersal across the Arctic, impacting Indigenous communities today. However, understanding these themes is still hampered by the challenges of building accurate site-based and regional-scale radiocarbon chronologies, influenced by driftwood use and marine reservoir effects. This paper presents a new high-resolution chronology for the key Ekven mortuary complex, located on the western shores of the Bering Strait, a site that defines the Old Bering Sea (OBS) phase, marking the earliest onset of the Neo-Inuit tradition. It is based on new direct radiocarbon dates of ancestral human remains and Bayesian modelling using OxCal, which considers available stratigraphic information, individual diets, previously dated faunal remains, and appropriate and species-specific ΔR values to account for marine reservoir effects. Our results suggest that the OBS emerged in this region at around 500 CE, which is later than some models predict, and indicate that the mortuary complex was used intensively until c. 1000 CE, after which few individuals were buried. The revised chronology also creates new opportunities to better integrate diverse cultural and palaeoecological proxies from across the Bering Strait, potentially clarifying the precise role of climatic and environmental factors at a key cultural juncture in Arctic prehistory.
Keywords
Introduction
Two major research themes are central to understanding Arctic Prehistory: (a) the emergence of specialized Arctic maritime economies; and (b) the role of Arctic maritime economies in the rise and expansion of the Neo-Inuit tradition out of the Bering Strait (Friesen and Mason, 2016). Specialized Arctic maritime adaptations involved intensified exploitation of walrus, and later, the coordinated hunting of bowhead whale (Fitzhugh, 2016). In turn, these new strategies were coupled with increasing sedentism, rising socio-political complexity, and compelling archaeological evidence for status competition, violent conflict, elaborate artwork and rich place-based mortuary traditions. Around the seasonally frozen waters of the North Pacific Arctic and Subarctic, these integrated socio-economic developments occur relatively late in the Holocene, with marked acceleration after around 2,500 years ago (Fitzhugh, 2016).
Despite their importance, the precise timing and sequence of developments leading to the rise of the Neo-Inuit tradition are understood only in general outline. The earliest phase of the Neo-Inuit tradition is defined by the Old Bering Sea culture (OBS), which then becomes ancestral to a series of daughter cultures including Birnirk and Punuk that eventually transition into Thule culture (Friesen and Mason, 2016; Gibbon and Ames, 1998; Gulløv and McGhee, 2006; Michael and Rudenko, 1961; Prentiss et al., 2023), at around 1000 CE (Ackerman, 1984). However, despite the pivotal cultural significance of the opening OBS phase, many aspects of its chronology, duration, and particular climatic and environmental context remain heavily debated.
The OBS culture refers to several settlements and burial sites on the coasts of the Chukchi peninsula, Alaska and Islands of the Bering Sea (Mason and Rasic, 2019). The study of the OBS culture has naturally focussed on their many coastal occupation sites, burial sites as well as the artefacts and tools recovered during excavation. One of the largest OBS sites, and one of the most important for the definition of the OBS culture, is the site of Ekven, located on the Chukchi Peninsula (Figure 1). The Ekven mortuary complex contains over 300 burials (Figure 2), with more burials likely unidentified. Although the nearby occupation site Uelen also had a small burial site, Ekven most likely served as a burial site for several other settlements along the coast. Excavated material points towards a culture with complex marine hunting technologies with a distinctive aesthetic style. Carved wooden masks, bone jewellery, and intricately decorated ‘winged objects’ (now commonly understood to be harpoon counterweights) have been excavated. The people of the OBS culture were, as would be expected of a coastal arctic community, marine hunters with their primary economic focus being large marine mammals such as walrus and seals.
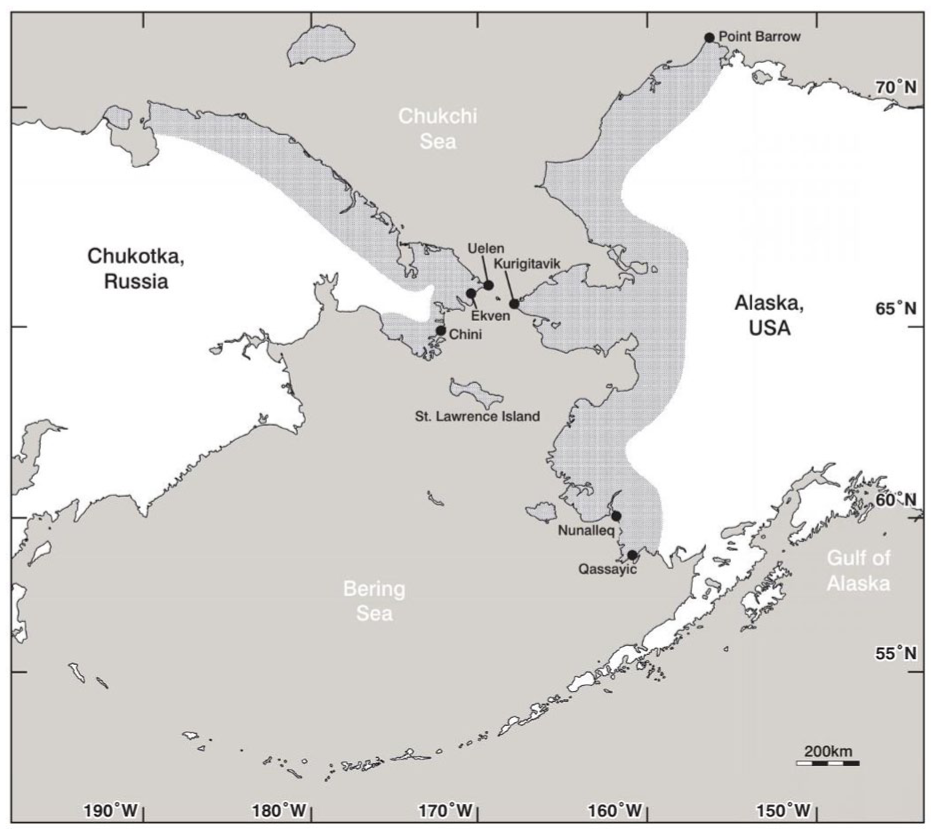
Location of Ekven mortuary site on the Chukchi peninsula, and the known distribution of OBS culture.

(a) Image of a polar bear carved from walrus ivory, 5.5 cm long, recovered from burial 308 (SMOA 778-Dr-IY); (b) slate ‘ulu’ knife, 11.5 cm long, with carved walrus ivory handle recovered from burial 313 (SMOA (State museum of Oriental Art) 784-Dr-IY; (c) burial 297 showing the adult male individual flanked by two whale bones; (d) view from the sea looking towards Ekven settlement (with material eroding out of the low banks onto the beach) and Ekven burial site (located on the hill above the settlement).
Although the chronology of the OBS has been studied in depth over many decades, several publications still conclude that the origins and precise chronology of the OBS are poorly understood. Some ‘high’ (or old) chronology estimates put the Okvik (early OBS) before 200 BCE with the entire OBS sequence predating 500 CE; more recent estimates place the height of the OBS Culture between 650 and 1250 CE, with its earliest phase Okvik and Ipiutak from 300 to 600 CE (Mason and Rasic, 2019). Archaeological excavations of Thule sites have confirmed the Bering Strait as the location of the emergence of the Thule (Mathiassen, 1927). The reconstruction of the origins of the OBS culture and cultural evolution of these societies in the Bering Strait region, however, remains a pressing question (Gusev et al., 1999). Reconstructions of the cultural history of coastal Chukotka are based on cultural artefact typologies from comparative studies of artefact assemblages in North America (Gusev et al., 1999). The development of the OBS culture is seen as uncertain and their temporal relationship with other archaeological traditions being somewhat vague (Mason, 2016). To date, the chronology of the OBS cultures has been based primarily on slight variations in their tools (harpoon points in particular), geographic distribution and housing styles. Given the well-documented uncertainties surrounding the chronology of the OBS culture, however, it is important to calculate absolute dates for the sites and artefacts being discussed.
The timing of cultural shifts has been made difficult by a wider lack of radiocarbon dating in the region, exacerbated by the old-wood problem and the marine reservoir effect. Further radiocarbon dating of OBS material is therefore necessary to shed light on the chronology of these eastern Siberian cultures. The Ekven burial site presents the perfect opportunity to do so. Care must be taken to account for the marine reservoir effect around the Bering Strait and to fully utilize any stratigraphic information which could help reduce calibration uncertainty. For the burial site of Ekven to be better understood, and by extension the entire OBS culture, we have radiocarbon dated 42 burials, conducted stable isotope analysis to quantify the amount of marine-derived carbon, and performed Bayesian radiocarbon modelling, which takes account of the relationships between samples, to date the site with a higher degree of accuracy.
Material and methods
Collagen extraction
In total, 5 bone samples and 37 teeth from 38 distinct humans from the site of Ekven were sampled for analysis. All dentine collagen extraction was performed at the University of Groningen using the laboratory’s standard extraction method (Dee et al., 2020). Tooth samples were cut into halves, using a dremel. Samples were subject to an ABA pretreatment: This involved the samples being decalcified over 24 h using weak acid (HCl, 2%–4% w/vol) at room temperature. If the material was not soft after 24 h, the acid solution was refreshed and soft portions removed and stored separately in demineralized water until further preparation. When the solution had stopped reacting and all the fragments had become soft and pliable, they were rinsed thoroughly with demineralized water. The samples were then exposed to NaOH (1%,~30 min) to eliminate humic acids, rinsed to neutrality, treated again with acid (HCl, 4% w/vol, 15 min) and then rinsed once more to neutrality. The raw collagen fraction was then denatured to gelatin in acidified demineralized water (pH3) at 80°C for 24 h. The dissolved gelatin was then filtered through a 50 μm mesh to eliminate any remaining foreign particulates and thoroughly dried. Finally, the crystalline product was scraped from the sides of the glass vessels. Between 3.5 and 5.5 mg of collagen was weighed out into tin capsules for analysis.
Bone collagen was extracted at Stockholm university. Samples of bone powder were obtained using a dentist’s drill. Surface layers were discarded to avoid contamination. Bone collagen was subsequently extracted according to the methods outlined by Brown et al. (1988). This included an ultrafiltration step (AmiconⓇ Ultra-15 Centrifugal UltracelⓇ Filters) to remove low molecular weight material (<30 kDa), in which the samples were centrifuged for 15 min at 3000 rpm and the remaining solution subsequently freeze-dried. Bone collagen was weighed into tin capsules (3.5–5.5 mg).
All IRMS and AMS analysis of both bone and dentine samples took place at the University of Groningen’s Centre for Isotope Research.
Shell sample pretreatments
In addition to the bone and dentine samples, two shell samples were prepared for analysis. According to Groningen AMS facility practices (Dee et al., 2020), the shell samples were etched with acid (HCl, 4%), rinsed with deionized water, dried and crushed to powder. An aliquot (~20 mg) of the crushed shell was placed in one arm of a glass reaction vessel. A saturated acid solution (H3PO4) was pipetted into the second arm, and the vessel was evacuated on a vacuum line. The acid was then tipped onto the shell fragments and the glass vessels placed in a water bath (25°C, 24 h). The CO2 evolved over this period was cryogenically transferred into a glass flask for stable isotope (δ18O and δ13C) analysis, and the subsequent graphitization and 14C measurement.
Stable isotope analysis
IRMS analysis at Groningen utilizes an Elementar Vario Isotope Cube™/Isoprime 100™ EA/IRMS combination. The isotopic content of materials is expressed in delta (δ) values, which are defined as the deviation (expressed in per mil) of the rare to abundant isotope ratio from that of a reference material:
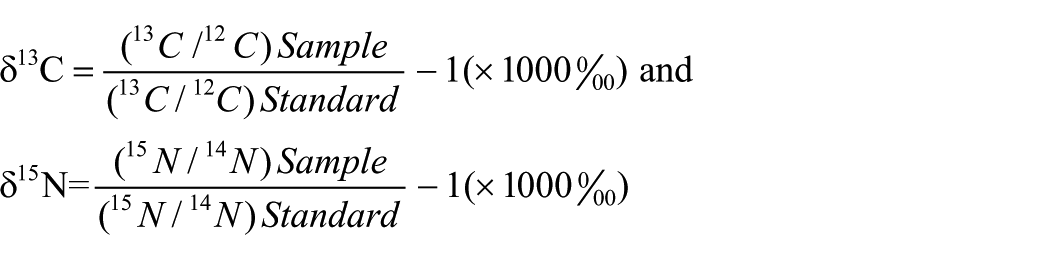
For carbon, the reference material is belemnite carbonate (V-PDB); for nitrogen, the reference is ambient air (Mook, 2005). The analytical error is typically 0.1‰ and 0.2‰ for δ13C and δ15N, respectively.
The atomic C/N ratio is a proxy for the integrity of the collagen. The widely accepted range of atomic C/N ratios for well-preserved bone and tooth collagen is 2.9–3.6 (DeNiro, 1985), which we have applied here.
AMS 14C dating
The extracted collagen was combusted to CO2 by an Elemental Analyser, connected to a Stable Isotope Mass Spectrometer (EA/IRMS, Elementar Vario Isotope Cube™/Isoprime 100™). Part of the CO2 is transferred into graphite, by a reaction with H2 gas at a temperature of about 600°C, using Fe powder as catalyst (Aerts-Bijma et al., 2001). The graphite was pressed into target holders for the ion source of the AMS. The AMS is a MICADAS-17 (IonPlus®) Mini Carbon Dating System (Synal et al., 2007). The present Groningen laboratory code is GrM.
The 14C dates are reported by convention in BP, that is, measured relative to the oxalic acid standard, corrected for isotopic fractionation using the stable isotope ratio δ13C = −25‰, and using a half-life value of 5568 years (Van der Plicht and Hogg, 2006).
Dietary modelling
Stable δ13C and δ15N isotopes measurements of the sampled human remains were modelled as dietary indicators, using the multi-isotope mixing programme FRUITS v. 3.1 (Fernandes et al., 2014) alongside measurements from published marine and terrestrial fauna (Britton et al., 2013; Chu, 1998; Dury et al., 2021; McManus-Fry et al., 2018). Average isotopic values for the previously analysed terrestrial fauna (δ13C = –20.5± 1.8, and δ15N = 4.1± 2.1‰) and marine fauna (δ13C = –13.4± 1.1, and δ15N = 16.7± 2.2‰) were used as endpoints. Because the OBS were marine mammal hunters, average pinniped δ13C and δ15N values represent the marine dietary endpoint. Consumer fractionation offsets of 1.5 ± 0.5‰ for δ13C and 4.5 ± 1.5‰ for δ15N were built into the non-weighted, concentration-independent, model. In addition to being important for the understanding of dietary behaviours, estimates of dietary protein sources are crucial for the modelling of radiocarbon dates.
Radiocarbon modelling
A comprehensive study of paired radiocarbon dates and δ13C and δ15N data of faunal material from the Ekven burials, including 13 marine and 25 terrestrial skeletal elements from 18 closed grave contexts, together with data from the wider region, forms the basis for estimation of the species-specific ΔR values used in the model (Dury et al., 2021). All 14C dates were calibrated in OxCal v.4.4.4 (Bronk Ramsey, 2009a). A radiocarbon model was constructed incorporating as much appropriate archaeological information as possible to reduce modelled date uncertainty. The model was built according to the following:
The model used a resolution of 1 year.
All samples and functions were modelled within a single unrestricted phase.
Within this phase, all samples were modelled within overlapping phases according to their burial contexts. Contemporaneous material from the same grave contexts was modelled together. This included previously radiocarbon-dated terrestrial and marine faunal samples (Dury et al., 2021).
Where more than one 14C measurement existed for a sample of a single skeletal element, an R_Combine function was applied.
Where more than one 14C measurement from an individual existed from two separate skeletal elements, a ‘Difference’ function was used to define the order of formation of these samples.
Terrestrial and marine samples were calibrated against the Intcal20 and Marine20 curves respectively (Heaton et al., 2020; Reimer et al., 2020).
Where possible, a measured ΔR value specific to each marine sample was applied; where a sample-specific ΔR value could not be calculated, a species-specific ΔR was applied (Dury et al., 2021).
For each human sample, a ΔR value of 289 ± 124 was applied. This reflects an average of the marine mammal ΔR values calculated from the Ekven burial site and chronologically relevant data from the wider Bering Strait region (Dury et al., 2021). The marine dietary protein contribution towards human bone collagen and dentine collagen was modelled and applied using a Mix_Curve function.
Any stratigraphic relationships between burial contexts were applied in the form of sequential, non-contiguous phases.
All 14C R_Dates (or R_Combine functions) were subject to an outlier analysis model allowing for 5% uncertainty.
For each burial context, the burial date was modelled. This was calculated by defining offsets between each R_Date and the Burial date. For each R_Date/R_Combine date from a human sample, the offset was defined as being the estimated ‘age-at-death’, minus the estimated ‘age-at-sample-formation’. For each R_Date from a non-human sample, the offset was defined as being between 0 and 50 years (this larger offset allowed for the possibility of slightly older terrestrial material having been incorporated into the burial context).
Results
AMS and IRMS data
The results of the stable isotope analysis and radiocarbon dating are reported in Table 1 and Figure 3. The atomic C/N ratios, all between 3.1 and 3.3, were used to assess collagen quality (DeNiro, 1985). Carbon and nitrogen concentrations were also considered (Ambrose, 1990; Sealy et al., 2014; van Klinken, 1999) to ensure C/N ratios reflected well-preserved collagen. All the samples were judged on these bases to have collagen of adequate quality. The standard deviations of the samples’ 14C dates are generally low (15–40 years), with the average error being ±20 14C years. The 14C dates range between 1375 ± 20 BP–2295 ± 20 BP, and the δ13C and δ15N values range from −12.6‰ to −10.9‰ and from 19.7‰ to 22.2‰ respectively. The error of the stable isotope measurements was between 0.1‰ and 0.3‰ for δ13C and 0.1‰ for δ15N.
δ13C, δ15N and 14C isotope data for human bone and dentine samples. Age at death estimates were made based on several osteological indicators. Sample formation periods of dental samples were based on estimates by AlQahtani et al. (2010).
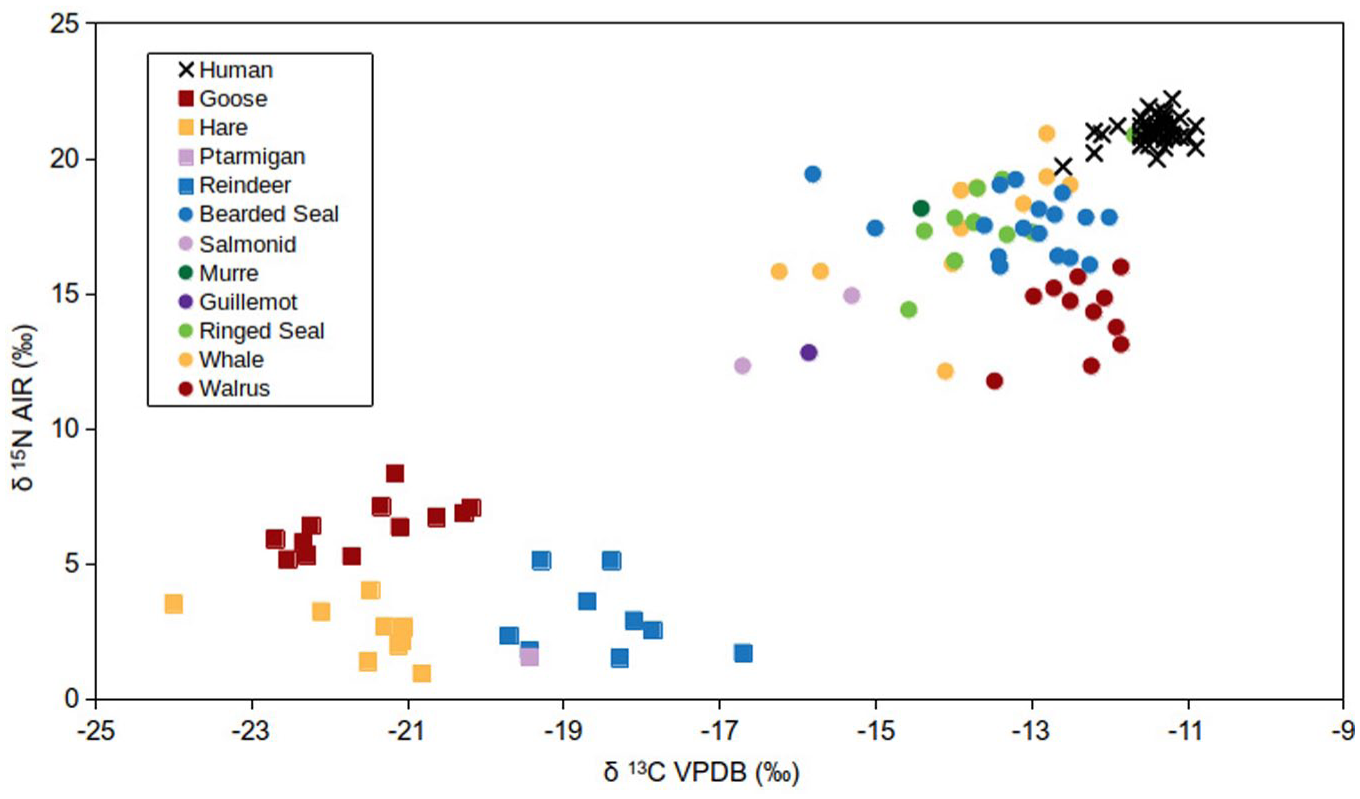
δ13C and δ15N data for the human samples from this study, as well as local terrestrial and marine fauna from Ekven and Nunalleq (Britton et al., 2013; Chu, 1998; Dury et al., 2021; McManus-Fry et al., 2018).
Human dietary modelling
The stable isotopic data of the humans and published fauna (Britton et al., 2013; Chu, 1998; Dury et al., 2021; McManus-Fry et al., 2018) makes it clear that those interred at the burial site, during their life, subsisted almost entirely on marine resources. Given the abundance of archaeological data pointing towards marine mammal hunting, this is not surprising. Although it appears clear, based on δ13C and δ15N values that the individuals buried at Ekven consumed a large amount of marine food, precise dietary estimates were achieved using the stable isotope mixing-model tool, FRUITS. The estimated contributions of marine protein to human bone and dentine collagen are presented in Figure 4. The data point towards a population whose main dietary input was marine; estimates range between 85% and 92% marine protein contribution. Here, pinniped stable isotope values have represented the marine dietary component. In subsequent radiocarbon modelling of the human 14C dates, the individual marine contribution estimate (±1SD), was applied to define the mixing of the Intcal20 and Marine20 curves for each human 14C date (Heaton et al., 2020; Reimer et al., 2020).
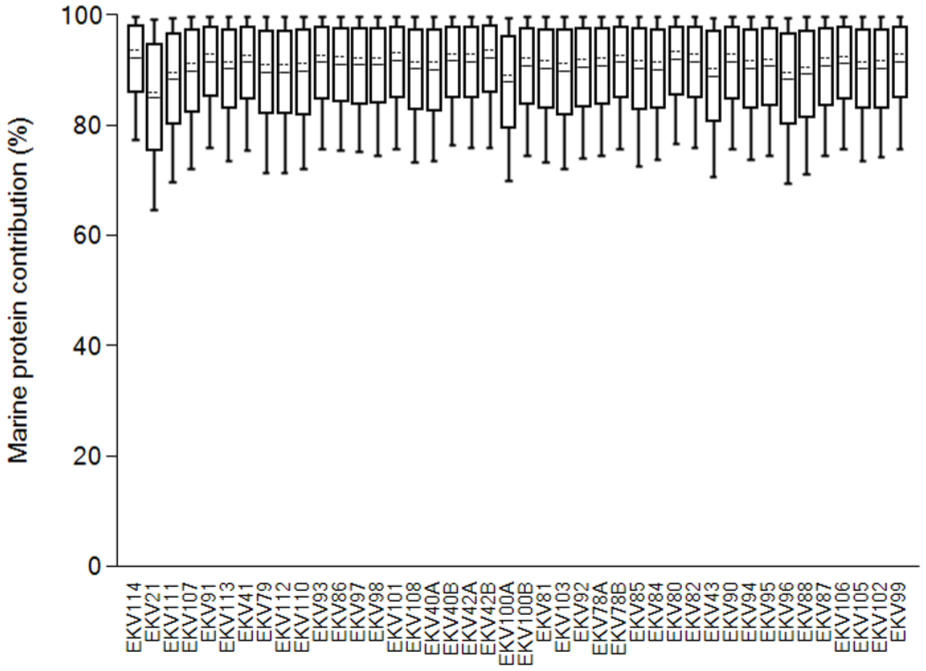
Marine protein contributions to diet for each human sample as calculated by FRUITS (Fernandes et al., 2014). The boxes provide a 68% confidence interval while the whiskers provide a 95% confidence interval. The solid and dashed lines represent the mean and median estimates respectively.
Burial date modelling
Table 2 and Figure 5 presents the modelled burial dates at the Ekven mortuary site (95.4% probability). All samples passed outlier analysis checks (with agreement equal to or above 95%, Bronk Ramsey, 2009b), other than EKV21 from burial 260, with an agreement of 93.4%. Moreover, all samples passed agreement index checks with values above 60% (Bronk Ramsey, 2009b). The majority of modelled burial dates plot between the years 500 and 1000 CE. The burial 280 is modelled as being the earliest, dating between 256 and 788 cal CE. Burial estimates have in many cases benefitted greatly from the inclusion of non-human faunal dates or stratigraphic information – both for narrowing down the estimated date range and for providing a more accurate date of the burial event (Supplemental Figure S1). For instance, in burial 318, with dates from one marine and two terrestrial faunal samples, together with one human sample, the modelled burial event has a very limited range of 634–798 cal CE. The burials 285-G and 304-B are known to be superimposed on, or cut into, the earlier burials 285-A and 304-A, respectively. When stratigraphic priors are included in the model, the human samples from these burials are more accurately calibrated. Of all the data considered in the model, only three estimated burial events fall mostly after 1000 CE, burials 324 (813–1345 cal CE), 322 (1013–1479 cal CE) and 260 (1022–1574 cal CE).
OxCal modelled burial date estimate (95.4% probability) along with the sampled material and stratigraphic priors modelled.
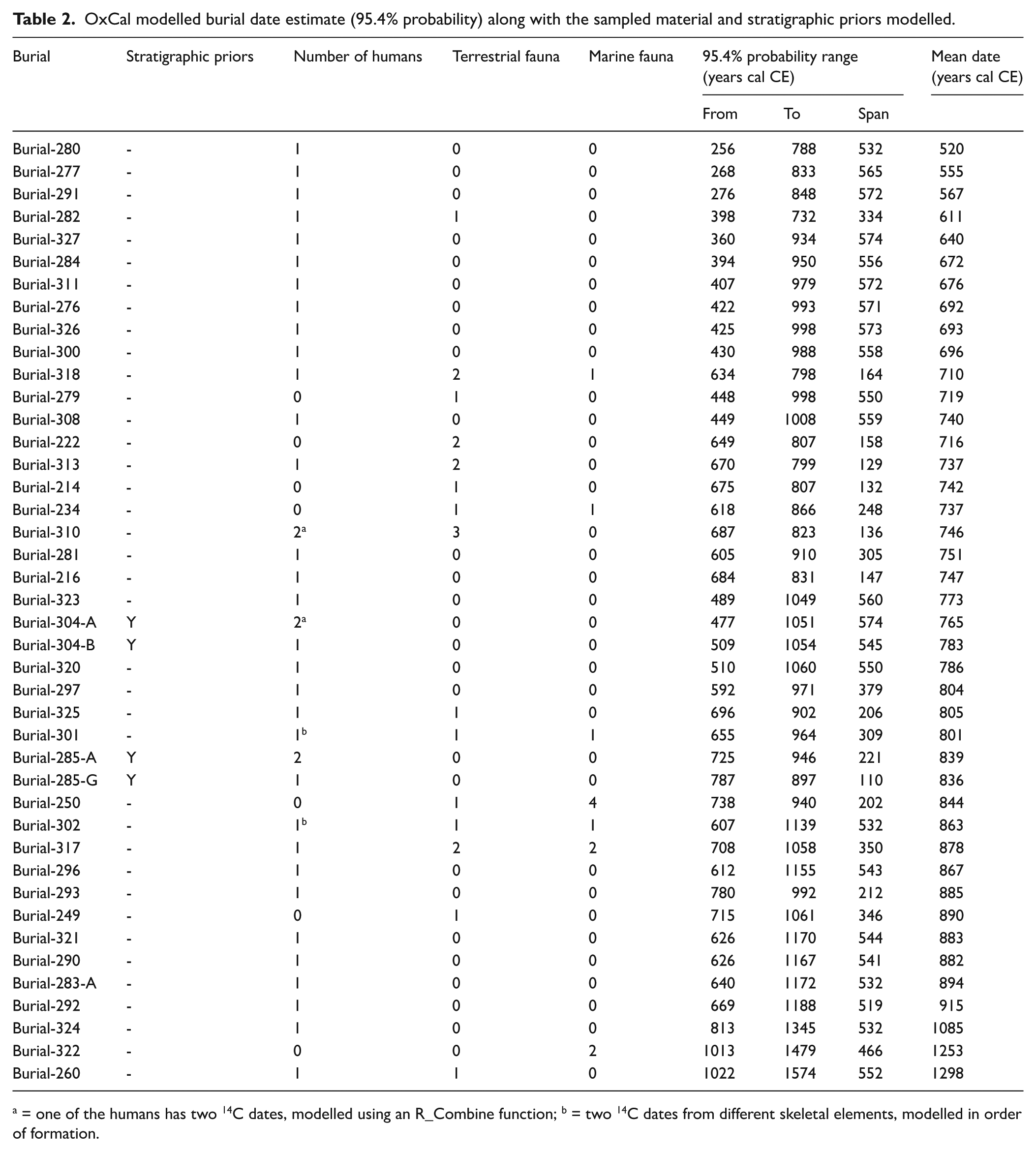
a = one of the humans has two 14C dates, modelled using an R_Combine function; b = two 14C dates from different skeletal elements, modelled in order of formation.
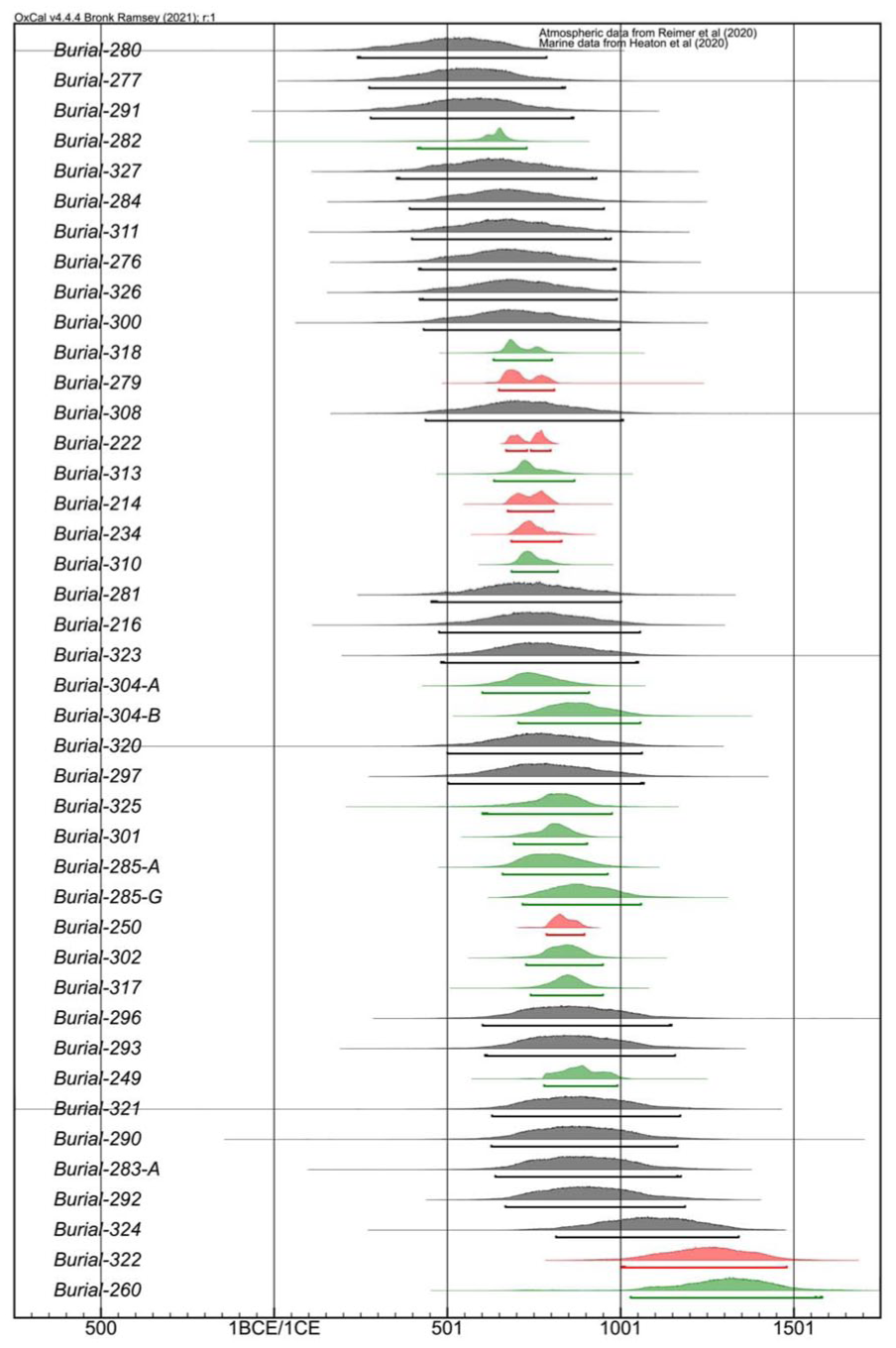
OxCal v.4.4.4 modelled Ekven burial dates. Burial estimates coloured red have been modelled based only on non-human faunal remains. Burial estimates coloured grey have been made based on a single 14C date from a human sample. Burial estimates coloured green have been modelled based on one or more human sample 14C dates, with additional stratigraphic information or non-human faunal dates.
Discussion
The archaeological record was examined to look at human diets in more detail. From their material culture (hunting equipment and refuse), it is known that OBS humans focussed their hunting efforts on large marine mammals, seals and walruses. Their diet was most likely supplemented by fish and marine birds, although they are often overlooked (Mason, 2016; Savinetsky, 2002). Shellfish have been described as somewhat of a starvation food, only consumed when necessary (Eidlitz, 1969). The OBS did not specifically target whales for hunting. Whale hunting cultures of the Bering Strait only emerged after the disappearance of the OBS culture. Opportunistic consumption of whales may have taken place by OBS groups, but their material culture does not point towards routine hunting or reliance on whales (Mason and Rasic, 2019). It must be remembered, however, that faunal assemblages are not the most reliable indicator of consumption. Differential processing of different species can lead to skewed proportions of consumption. It is more likely, for example, that large mammals were butchered close to the shore rather than dragged to the settlement, whereas fish were more easily transported and brought back to the settlement. Fish remains, however, suffer from poor survival in the archaeological record and are more likely to be missed during excavation. Large whale bones, on the other hand, will certainly be recorded during excavation, but during the site’s occupation, will most likely have been recycled as building materials (Habu and Savelle, 1994; McCartney and Savelle, 1993; Savelle and Habu, 2004), meaning that they accumulate beyond their expected numbers in cultural deposits. Terrestrial animal remains were also recovered from burial assemblages, however, most evidence (isotopic and artefactual) point towards large marine mammals being the primary food source of the OBS culture.
The data here are significant for the debate surrounding the emergence and evolution of the OBS culture in the Bering Strait. Though it is known that the OBS developed into the Punuk and Birnirk cultures and is the ancestral culture of Thule, there is currently no commonly accepted chronology for the cultures of the Bering Strait. The radiocarbon dating of OBS material has faced problems, including large and varied marine reservoir effects as well as the old-wood problem (Gerlach and Mason, 1992). The most recent and thorough reinterpretation of published 14C dates (Mason and Rasic, 2019) point clearly towards a more recent dating of the OBS culture.
The earliest dated burial (280) dates to 256–788 cal CE, and the most recent (Burial 260) to 1022–1574 cal CE. This span of the burial dates fits far better with the low (more recent) OBS chronology suggested by Mason and Rasic (2019), than with a high chronology. It is likely that the ‘high OBS chronology’ (old chronology) estimates are caused by the old-wood problem or not fully accounting for the marine reservoir effect. It is clear from an inspection of the data, however, that the vast majority of the sampled individuals calibrate very comfortably between the years 500–1000 CE, pointing towards a more recent OBS culture. Only three burials’ 95.4% modelled date distributions fall almost entirely after 1000 CE.
The data gathered here fits perfectly with the interpretation that the OBS culture developed within a period of climatic turbulence (Büntgen et al., 2016) that coincided with a major demographic surge between 500 and 1000 CE (Mason and Rasic, 2019). It may be that the intensification of storms during the fifth century CE and a greater upwelling of nutrients from the ocean floor led to an increase in the population of sea mammals (Mason and Gerlach, 1995) which in turn allowed for the full emergence of the OBS culture. By 1000 CE, the whale hunting Thule culture were able to supplant the OBS cultures in the Bering Strait (Mason, 2009),
The calibration and modelling of the 14C dates of the Ekven humans, appear to re-enforce the more recent OBS chronology. It must be remembered, however, that a cultural phenomenon may persist in certain locations whilst being supplanted in others, and the history of the OBS culture may have earlier roots at other locations. Moreover, with sea level rises and coastal erosion, the possibility that earlier, high-chronology OBS sites have been lost, should not be forgotten. This would lead to an absence of evidence of a high OBS chronology.
The burial-date estimates of burials 324 and 260 (represented by human samples EKV96 and EKV21 respectively) and burial 322 (represented by shell samples) stand out as more recent than other modelled burials, with mean burial estimates being post 1000 CE. It is therefore likely that the use of the burial site continues after the disappearance of the OBS culture. Given the total number of sampled individuals and the fact that the majority of samples calibrate so neatly between 500 and 1000 CE, it seems that the burial site was only in use intermittently after about 1000 CE. It may be the case that the burial site lost its wider regional importance and was used only by a smaller, more local population. This might explain why the proportion of burials that most likely date post-1000 CE is only three out of the 42. However, what these recent burials demonstrate is that the mortuary site was not forgotten by local people and that the site was not viewed as unusable, but rather as a site appropriate for contemporary burials.
Four modelled estimates for the burial site’s use are displayed in Figure 6. Burials 260, 322, 324 and 292 are the four most recent burials. Burial 280 is the oldest modelled burial. Assuming that the duration of the site’s use is measured based on modelled burials with mean dates falling before 1000 CE, (i.e. Burials 280–292), its use span is modelled to 395 ± 185 years. If the site’s use is based on burials with 95.4% date distributions falling at least partially before 1000 CE (Burials 280–324), the site’s span is 565 ± 192. If all burials are considered (Burials 280–260) the period of use spans 778 ± 196 years. The omission of only the most recent three graves almost halves the estimate of the site’s use.
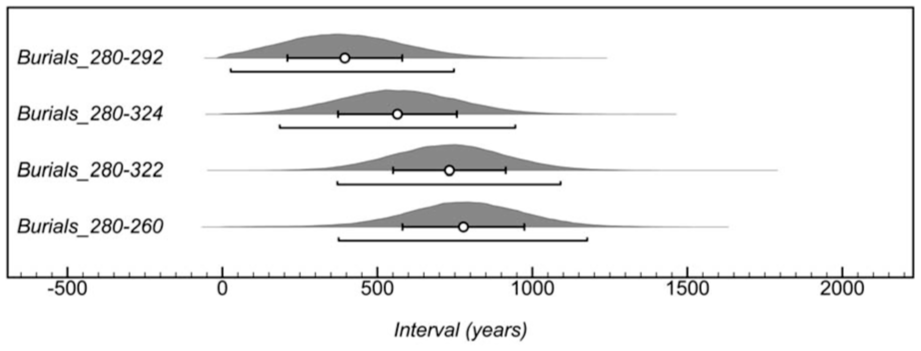
Four separate estimates of the duration of use of the Ekven mortuary complex, calculated by measuring the span between the earliest grave (280) and the four most recent graves (burials 292, 324, 322 and 260). The white point and error bars represent the mean estimate ± 1σ.
The model was rerun with a prior added to all individuals’ 14C dates, stating that the modelled dates should fall during the first millennium CE. This was designed to test if the burials 324, 322 and 260 (the only burials with mean dates post 1000 CE) could be reasonably modelled to within a more traditional OBS time frame, before 1000 CE. It should be remembered, however, that the date 1000 CE has been selected arbitrarily to mark the best current estimates of the emergence of the Thule culture, the date 1000 CE has not been generated or calculated by the model. With this dating prior applied, the human sample EKV96 in Burial 324 has an agreement index of 40%. Though it is possible that the burial can be modelled before 1000 CE, the modelled date distribution of sample EKV96 compared to its un-modelled distribution deviates too much for the 1000 CE constraint to be considered reasonable for this burial. The same conclusion should be reached for burial 322. Burial 260 is more complicated. The hare sample (EKV39) and human sample (EKV21) yielded very different modelled dates. Because the burial is defined by its human remains, and because there is no reason to believe either sample has been contaminated before analysis, it should be concluded that during the burial event, the hare bone was introduced into the burial as a historic relic or through other actions. Even though the human sample narrowly failed outlier checks (95%), considering its context, the modelled date for burial 260 should not be discounted. To any reasonable degree of certainty, the burial 260 cannot be considered to have been buried before 1000 CE. This model does demonstrate that the interpretation of Ekven as an OBS mortuary site should not be restricted to old chronology estimates of the OBS culture. Burials continued post-1000 CE, indicating a longer-lived OBS culture or continued use of the site by Thule groups. Ekven could be considered to have a more complex history, with a longer use span than previously thought, with an early, intensive burial phase, and after c. 1000 CE, a phase characterized by infrequent burials. Moreover, the possible conclusion that individuals (represented by samples EKV21 and EKV96 from burials 324 and 260, respectively) post-date the OBS culture, could be supported by their δ13C values. More routine whale hunting in the Bering Strait emerged with the Thule culture c. 1000 CE. Their low δ13C values (–12.6‰ and −12.2‰ respectively), relative to the mean of the rest of the population (δ13C = −11.4± 0.3‰), could be explained by a larger dietary input from whale species, which have a lower mean δ13C value.
Conclusions
A large marine reservoir effect has already been demonstrated in the Bering strait (Dumond and Gryphon, 2002; Dury et al., 2021; Khasanov et al., 2022; Khassanov and Savinetsky, 2006; Krus et al., 2019; Reuther et al., 2021). Recent radiocarbon dating of material specifically from Ekven, with a focus on the species that were being targeted for consumption, has resulted in an appropriate ΔR value to apply to human calibration (Dury et al., 2021). Stable isotopic modelling of human remains within the context of the ecology of the Bering Strait points towards a diet almost completely dominated by marine foods. Archaeological evidence points towards pinniped hunting being their economic focus. These data have been factored into a large OxCal model, focussed on understanding the use of the Ekven mortuary complex.
Assuming the sampled individuals are a broadly-representative sample population, the site can be tentatively considered to have two phases of use. An early and intensive burial phase, coinciding with demographic growth in the region, and a later phase characterized by infrequent burials after c. 1000 CE. The earliest and latest burials from this ‘intensive use period’ have mean dates of 520 and 915 cal CE, respectively, and provide data for a use period of 395 ± 185 years. The most recent burial has a mean calibrated date of 1298 cal CE (median 1311 cal CE). Considering the classification of the burial site as an OBS site, and the majority of samples falling between 500 and 1000 CE, this is supportive of the more recent ‘low OBS chronology’.
Further dating of Ekven assemblages would be useful for understanding the ‘internal’ chronology and use of this particular mortuary complex; this may result in other ‘post-OBS’ burials being identified, and allow for higher overall dating resolution from the OxCal model. Subtle cultural changes in terms of burial traditions and grave goods could also be explored with the refined chronology. The use of OxCal modelling and the application of further prior information (i.e. stratigraphy and accompanying faunal 14C dates) should help in achieving this aim. Finally, similar methods and approaches can also be applied across the Bering Strait region, improving the chronological resolution of other sites and occupations, supporting inter- regional correlations, as well as improved integration of archaeological sequences and diverse climatic and palaeoecological proxies that are needed to develop a fuller understanding of the deeper roots, causes, conditions and long-range consequences of this major Neo-Inuit cultural transition.
Supplemental Material
sj-docx-1-hol-10.1177_09596836251366194 – Supplemental material for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex
Supplemental material, sj-docx-1-hol-10.1177_09596836251366194 for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex by Gunilla Eriksson, Arkady Savinetsky, Maria Dobrovolskaya, Kirill Dneprovsky, Alison JT Harris, Johannes van der Plicht, Peter Jordan and Kerstin Lidén in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_09596836251366194 – Supplemental material for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex
Supplemental material, sj-pdf-2-hol-10.1177_09596836251366194 for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex by Gunilla Eriksson, Arkady Savinetsky, Maria Dobrovolskaya, Kirill Dneprovsky, Alison JT Harris, Johannes van der Plicht, Peter Jordan and Kerstin Lidén in The Holocene
Supplemental Material
sj-pdf-3-hol-10.1177_09596836251366194 – Supplemental material for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex
Supplemental material, sj-pdf-3-hol-10.1177_09596836251366194 for The emergence of cultural complexity in the Bering strait: A refined radiocarbon chronology for the Ekven (Old Bering Sea) mortuary complex by Gunilla Eriksson, Arkady Savinetsky, Maria Dobrovolskaya, Kirill Dneprovsky, Alison JT Harris, Johannes van der Plicht, Peter Jordan and Kerstin Lidén in The Holocene
Footnotes
Acknowledgements
Primary sampling and analysis for this paper was carried out with the ArchSci2020 MSCA ITN (2016-2021) by ESR 9. KD would like to thank the local communities in Chukotka for collaboration and logistical support during the excavations. We would like to acknowledge the assistance of Markus Fjellström (Stockholm University), Heike Siegmund (Stockholm University), Sanne Palstra (University of Groningen), Sven de Bruijn (University of Groningen) and Andrius Garbaras (Vilnius University) for aiding in laboratory work and analysis. We would like to acknowledge Mike Dee (University of Groningen) for many useful discussions.
Author contributions
Funding
The authors disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project has received funding from the European Union’s EU Framework Programme for Research and Innovation Horizon 2020 under Marie Curie Actions Grant Agreement No. 676154. This research has also benefitted from additional PhD funding awarded to AJTH from the Social Science and Humanities Research Council of Canada. GE would like to thank the Knut and Alice Wallenberg Foundation for financial support.
Ethical approval and informed consent statements
Sampling, analysis and publication follow the ethical requirements of all relevant institutions. Given that the research published in this article involves analysis of materials associated with descendent communities in Chukotka, a written confirmation of informed consent was signed by the relevant community. A copy of this document is available upon request.
Data availability statement
All data generated or analysed during this study are included in this published article and its supplementary information files.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.