
Abstract
A combined plant sociological and vegetation historical investigation of a semi-natural coastal heathland in the mid-north boreal transition zone of northern Norway is presented. The site is at the northern limit of the EU-protected anthropogenic Atlantic coastal heath complex. This study documents woodland clearing and grazing as the main regulating anthropogenic impact, forming the vegetation of the heathland and confirms the status as a semi-natural ecosystem. Detailed survey of flora and soil parameters in permanent plots was performed in 1993 and 1994, following a stratified random sampling design. Analysis of grazed and ungrazed parallels after enclosure from sheep grazing in 1991, did not render significant results related to grazing impact. The results from this study may thus be a base for future monitoring of the heathland. The heathland is classified as a Vaccinium myrtillus/Empetrum nigrum-heath with two main units; Alchemilla alpina/Anthoxanthum odoratum- and Calluna vulgaris/Cladonia arbuscula-type. A central minerotrophic mire under ombrotrophy development is classified in a Tricophorum- and a Carex rostrata-type with hummocks containing dwarf shrubs and shrubs. Pollen analysis of this mire reveals an incipient possibly anthropogenic, opening of the local birch woodland from c. 2500 cal. BP. Clearing of birch woodland and intensified grazing 1500–1400 cal. BP imposed erosion and a hydrological change in the mire. The heathland has remained open until modern time, with a period of regrowth c. 750–350 cal. BP, in the aftermath of the Black Death. The anthropogenic influence is compliant with other archaeological and palaeoecological studies from the region.
Keywords
Introduction
The processes behind secondary succession in traditionally managed farmland and the environmental consequences of abandonment have been a central point of cultural landscape research for several decades and has gained new actuality with increased political focus on national sustainability. For future landscape planning, including the restoring of former outfields, it is essential to attain detailed knowledge about the driving mechanisms and time scheme of vegetation succession in different types of ecosystems, climate conditions and farming cultures.
The open coastal landscape of northern Norway includes heathland persisting as a dynamic equilibrium regulated by grazing, formerly also mowing and to some degree burning and is as such regarded as semi-natural ecosystems by the European Habitat Directive (The Habitats Directive – European Commission). The EU-protected Atlantic coastal heathland, which is dependent on regular burning, mainly follows the southern and middle boreal climate zone in Norway. Northward from central Norway it is gradually replaced by heathland maintained primarily by grazing. As stated by the Norwegian Biodiversity Information Centre (biodiversity.no), it can be difficult to separate the types of heathlands only by differences in species composition and diversity, without knowledge about former and present management regime. Microscopic charcoal observations in palaeoecological records indicate that fire has a long history in being a part of the management practices of heathland, as compiled by Vandvik et al. (2014).
Few detailed studies are performed on plant sociology and succession of cultivated land in the oceanic part of the middle and northern boreal bioclimate transition in Scandinavia. Tveraabak (2004b) studied lowland Calluna heaths from central Norway to eastern Lofoten and found a relationship to boreonemoral and southern boreal anthropogenic Calluna heath but with a significant trend towards more Empetrum nigrum dominance to the north. Jensen et al. (2001) studied the effect of abandonment on traditionally managed grass meadows and found a significant successional trend towards heath birch woodland over 35 years. Still, little is known about the regenerative ability and long-term succession of coastal forest communities that are subdued to anthropogenic impact in this region, and consequently the time needed to develop a persistent cultural ecosystem. The aim of this investigation is to increase our knowledge about the processes leading to changes in vegetation composition in the coastal cultural landscape and to document the establishment and development of a local coastal heath in a time span covering the agricultural revolution.
Farming as part of a sustainable marine-oriented economy goes far back in northern Norwegian coastal history (Nilsen, 2017). Over the past few decades, several archaeological-scientific excavations have proven agricultural activity as far back as the Bronze Age (1800–500 BCE) and as far north as 69°N (Arntzen and Sommerseth, 2010; Cerbing, 2016; Jensen, 2020; Jensen and Ahlqvist, 2016; Jensen and Arntzen, 2017; Jensen and Westling, 2013; Sjögren and Arntzen, 2013). Direct evidences are the combination of archaeological structures and artefacts related to a farming economy and radiocarbon dated bones from sheep or goat, radiocarbon dated cereals, weeds, and corresponding local environmental changes. Indirect empirical evidence also testifies to this in terms of archaeological stray finds (Arntzen, 2013; Valen, 2007) and clear anthropogenic signals indicating animal grazing and/or cereal cultivation from the biostratigraphical records of bogs and lakes near ancient settlements (D’Anjou et al., 2012; Sjögren, 2009; Sjögren and Arntzen, 2013; Tveraabak and Vorren, 2001; Vorren, 1986; Vorren et al., 1990; Vorren, 2005). However, the number of investigated sites from northern Norway is still low and the empirical data base not solid enough to systematically investigate details and variations in management regime as well as impact on the natural environment on a regional base.
The present investigation site Haugheia is in the Lofoten region of Northern Norway (Figures 1 and 2), which is considered as within the northern boundary of the anthropogenic Atlantic Calluna heath communities (Fremstad, 1997; Kaland, 1986, 1999; Kvamme et al., 2004; Moen, 1999; Nilsen, 2004; Tveraabak, 2004a, 2004b). Management of Calluna heath by means of fire has been documented from eastern parts of the Lofoten archipelago and is recognized by increased abundance of Calluna and microscopic charcoal combined with deforestation and indication of grazing in the peat stratigraphy (Tveraabak, 2004a, 2004b). However, historical evidence of a management practice comparable to the one documented further south is lacking. The northern coastal heaths are generally characterized by their high cover of Empetrum nigrum ssp. hermaphroditum relative to Calluna vulgaris, and presence of alpine species in the lowland, like for example, Kalmia procumbens, Arctous alpina, Phyllodoce caerulea, Veronica alpina ssp. alpina (Fremstad, 1997; Haapasaari, 1988; Moen, 1999). The grazing impact varies along steep ecological and topographical gradients, and a distinctive character of the northern coastal pastures is the small-scale mosaic vegetation pattern. The effect of erosion from trampling animals may be large in the undulating terrain with a thin sod layer.
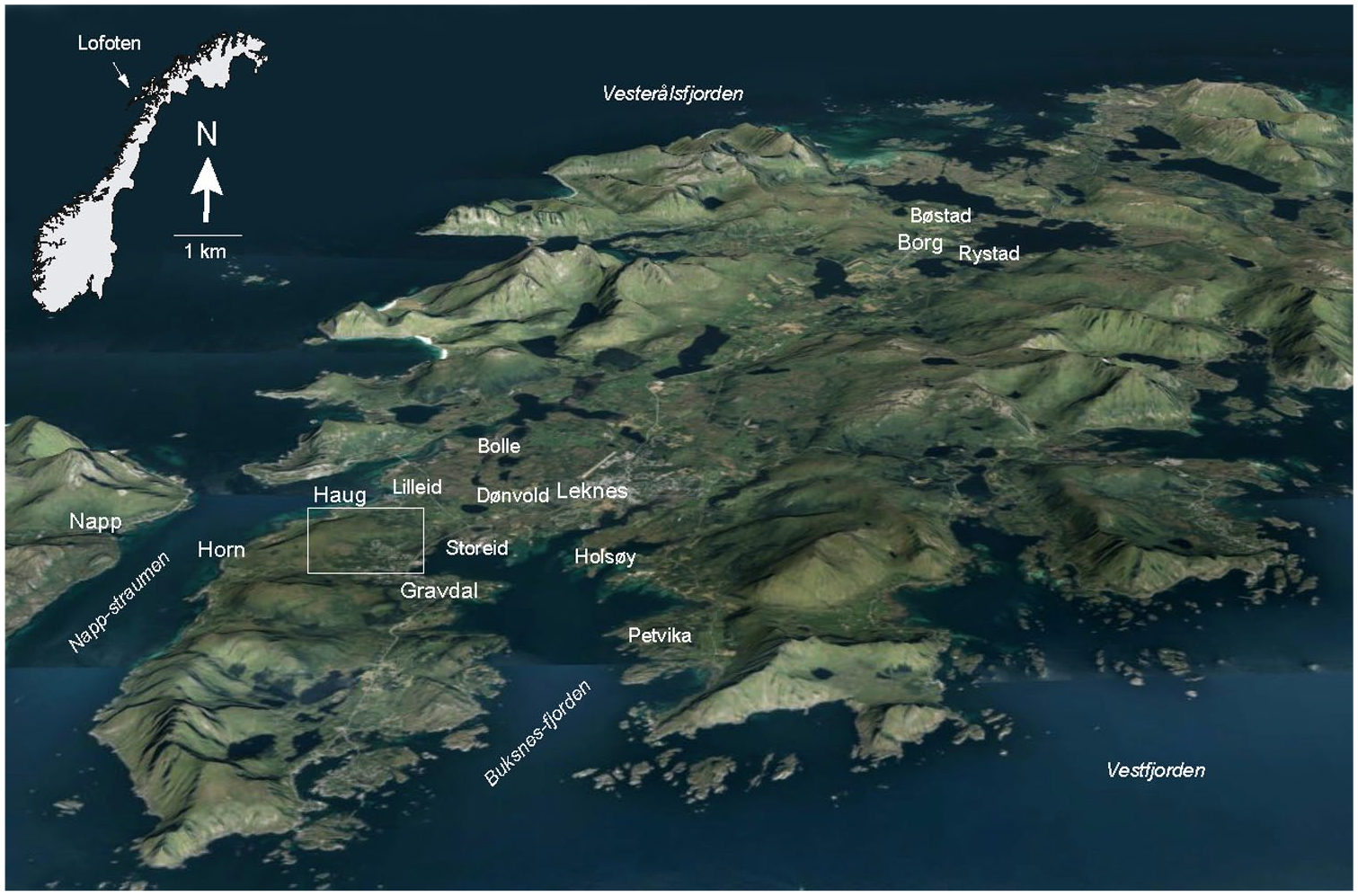
Vestvågøy island, Lofoten, Norway. Square indicates the Haugheia investigation site presented in Figure 2. Background 3D topographic map by Geodata.no.
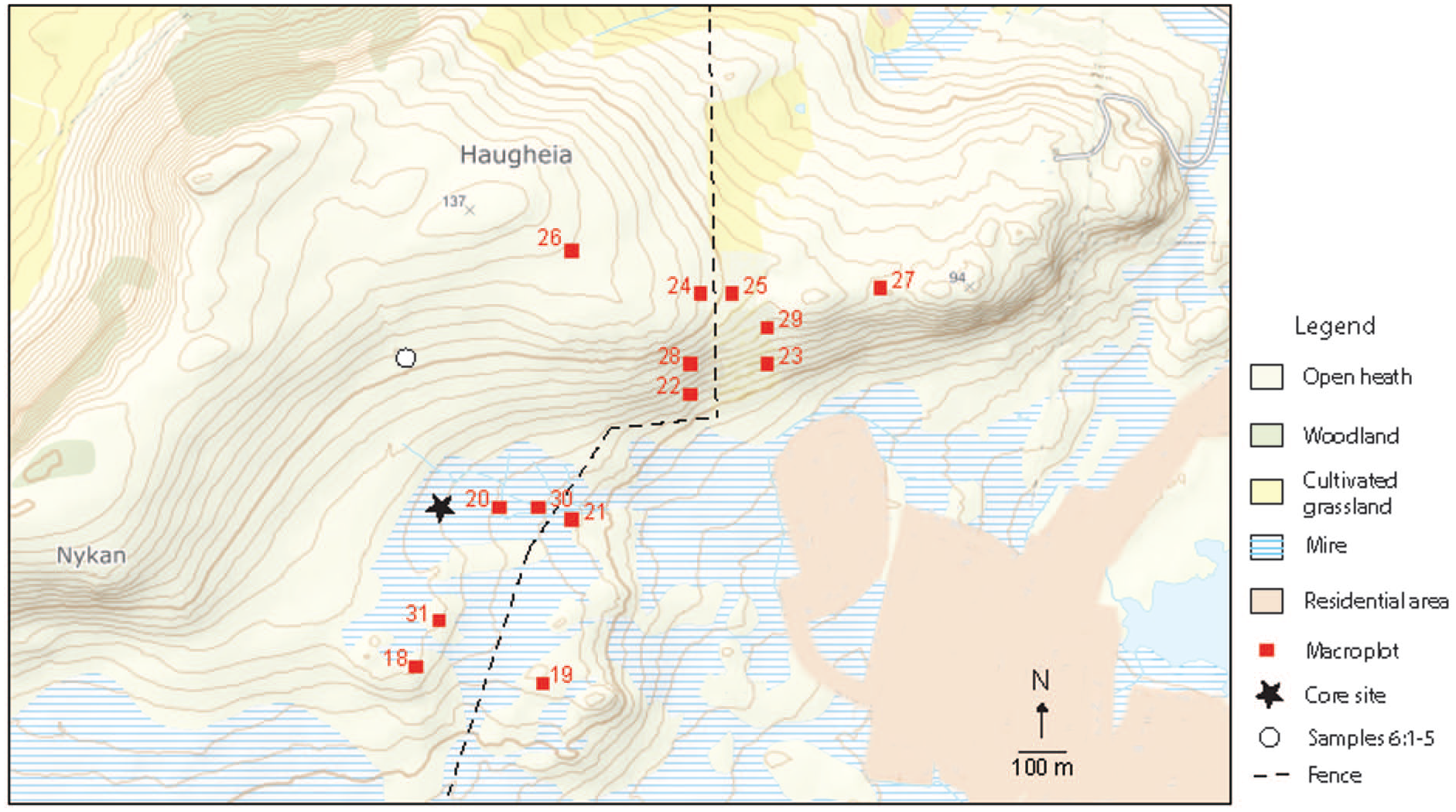
The investigation site Haugheia, Vestvågøy, Lofoten with macroplots for vegetation analysis and core site for peat profile.
A typical feature of the heathlands of the western Lofoten region is the zonation of vegetation relative to elevation and wind exposure. Dwarf shrub heaths are formed in the lowland, often closely connected with blanket bogs or sloping fens, while grass dominance increases with altitude and is mainly maintained by sheep grazing. Phytosociological and ecological studies of lowland mire complexes in the mid-north boreal coastal region includes the Bolle mire on Vestvågøy (Edvardsen, 1988), the Andmyra mire reserve (Buys, 1992) and a compilation of terrestrial mires from around 69°N (Vorren et al., 1999). The outcome of these investigations was detailed classifications of the mire morphology and vegetation along ecological gradients, discussed in relation to established European mire typology. The most comprehensive investigation of heathland vegetation in northern Norway and Finland, was carried out by Haapasaari (1988), followed up by Bråthen et al. (2024) in a re-investigation of his sites. These studies did, however, not include the Lofoten region.
The present project was initially designed as a monitoring project on vegetation succession and landscape planning financed by the Norwegian Research Council during 1993–1995. The project included three localities along the coast of northern Nordland and Troms counties, covering model sites on grazed and mowed heathland and meadow communities (Jensen et al., 2001). A grant from the University of Stavanger in 2010 made it possible to extend the approach to include long-term succession and development in studying the vegetation history of the heathland model site. The present publication will thus give an overall documentation and interpretation of the results from the heathland site Haugheia in the Lofoten region. The main research question is whether, and for how long, anthropogenic activity has played a dominant role in the development of a heathland complex of western Lofoten.
Site description
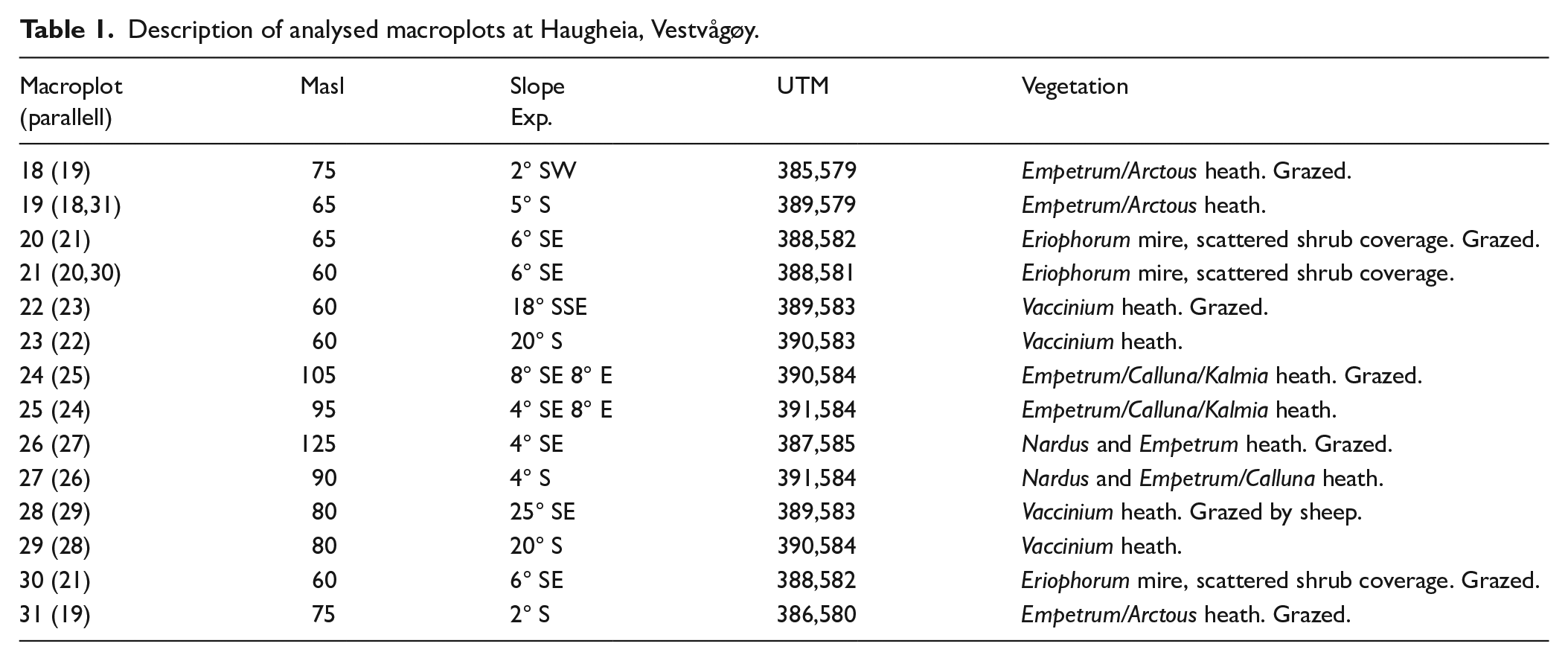
The investigation site covers c. 1 km2 centred on the uplands between the villages Gravdal and the farms Haug and Horn on the south-western part of Vestvågøy island, Lofoten at 68°N (Figures 1 and 2). The site is within a geotectonic contact zone between Precambrian mangerite and metamorphic sedimentary bedrock (Caledonian orogenetic ‘Leknes-group’) of quartzite, mica schist, amphibolite and marble (Tveten, 1978). The local vegetation ecological zone is middle boreal, markedly oceanic (Moen, 1999) which characterize much of the Norwegian coastal lowland between c 67 and 69°N. July mean temperature is 11–13°C and annual precipitation 1000–2500 mm based on data from weather stations (Moen, 1999, met.no/en). The higher parts of the uplands and the west-facing slopes consist of grass- and herb-dominated pasture, while the lower parts towards SE-NE consist of different formations of dwarf-shrub heath and mires (Figure 3a and b). The topography of Vestvågøy is influenced by the strong variations in relative sea level throughout the Holocene. The island is intersected by several valleys which used to be open straits. The lowland NE of the investigation site is such a broad valley, now consisting mainly of mires and shallow tarns.

(a) View towards north of the investigation site in 2010. The location of macroplot 18 is in the foreground. Star annotates the core site. Photo: C. Jensen. (b) Grass- and herb dominated Vaccinium heath, with a high grazing impact and erosion scars. Photo: C. Jensen.
The marine limit on Vestvågøy varies between 12 and 23 MSL and is measured at Haug and Gravdal as 12 and 15 MSL, respectively, adjusted for local conditions (Møller, 1984). Balascio et al. (2024) refined the timing of the Mid-Holocene transgression (Tapes-transgression) in western Lofoten by combining new sea-level index points to previously published data. The sea level increased from −4 MSL at 9000 cal BP, to +7 MSL at 6000 cal BP, followed by a decline to 4 MSL during the next millennium. A gradual decline in sea level is seen over the last c. 5000 years estimated to 0.8 m per 1000 years (op. cit.). Møller (1984) compiled studies on raised shorelines and terraces in the Nappstraumen area and found two major transgression phases during the Holocene; the Tapes transgression recognized at 9 MSL and a later one at 6 MSL The latter is probably seen at Petvika by marine sand and gravel overlying peat with birch wood dated at 4700 ± 110 BP (5650–5050 cal BP, 3700–3100 BCE; Moe, 1975). The discontinuity in settlement at the cave Storbåthallaren with a desertion phase between 4740 ± 140 BP (5800–5400 cal BP, 3900–3090 BCE) and 2460 ± 70 BP (2700–2350 cal BP, 770–400 BCE; Utne, 1973) may be a result of temporarily raised sea level.
Modern anthropogenic influence
The uplands have traditionally been outfields for the farms nearby. In 1959, 30 farms with totally c. 170 sheep and 10 horses used the area for grazing, while in 1993 three farms with c. 140 sheep, and later in the 2000s only one farm with c. 100 sheep. Some cattle grazed the area sporadically. The bogs were exploited for peat cutting until the 1960s. Haug- and Hornheia is a popular leisure area for the local people, but the trampling effect is mainly confined to a path up to the highest spot. The lower slopes are used for berry-picking in late summer and autumn. The eastern part of the area has been affected by the expansion of the village Gravdal and road constructions. The areal effect of farm abandonment and ‘urbanization’ is observed in the change of open fields and pastures from 95% in 1949 to 58% in 1991 of a mapped area of 430 hectares (Samuelsen et al., 1998). This development is mainly caused by regrowth of less productive land, like bogs formerly exploited for peat-cutting (an increase of 25%), and expropriation of land (mainly grazed and mown mire) for new settlements and road construction (an increase of 12%).
Prehistorical settlement
Humans probably arrived at Lofoten shortly after deglaciation, but any traces of settlements from the earliest times are below MSL or have been destroyed by the Tapes transgression (Glørstad et al., 2020). The oldest finds of possible agrarian origin are cattle bones from the rock shelter Storbåthallaren c. 6 km SW of Haug, dated to the Early Bronze Age (2740 ± 80 BP) and Pre-Roman Iron Age (2050 ± 60 BP; Utne, 1973). The region is rich in cultural heritage monuments, but few have undergone full archaeological excavation (The Directorate for Cultural Heritage, askeladden.no). In the immediate upland surroundings, the most relevant one is a N-S oriented 20 × 7 m house foundation of certain Iron Age typology, 54 MSL and c. 850 m N of the core site. Two Saami house foundations are found c. 800 mW of the core site. In addition to a mediaeval farm mound from the 1300s (Bakkan), the lowland below 15–20 MSL has several iron age grave mounds, where the islet Holsøya with 60 graves from the period 200–600 CE is central. Many prehistoric boathouse foundations are present as well, of which one near Leknes is radiocarbon dated to the Roman Iron Age/Migration Period (1685 ± 100 BP; Nilsen, 1997). Some of them bears evidence of multifunction purposes, such as the presence of culture layers with high charcoal content, fire-cracked rock, burnt bone and slag indicating residential use (Nilsen, 1997; Wickler and Nilsen, 2012).
Three courtyard sites were established on Vestvågøy during the Roman period and probably used until the Merovingian period, 200–600 CE. They are located at Bøstad near Borg, Gimsøy and Leknes (Figure 1), of which the Leknes courtyard c. 4 km NW of Haugheia was the largest one with 14 houses (Johansen and Søbstad, 1978). The marginal location relative to arable land and scarcity in positive finds related to agriculture resist an interpretation of the sites as farmsteads, but rather housings or gathering points for people connected with military defence, political or ceremonial activity (Johansen and Søbstad, 1978; Storli, 2000). The contextual and stratigraphic control of the charcoal which forms the basis of the four radiocarbon dates from the Leknes site is not good. However, all are on birch, and three of the dates overlap covering Late Roman Iron Age and early Migration period (1790 ± 90, 170 ± 70, and 1760 ± 90 BP), while the fourth is Late Merovingian until early Viking period (1270 ± 60 BP).
The investigation site is c. 16 km SW of the Late Iron Age chieftain centre at Borg (Figure 1). The surveys of Borg revealed settlements from 200 to 1000 CE, with particularly rich findings from the 8th to 10th century, indicating good contact with social elites in Europe (Munch et al., 2003). The first longhouse at the site was established in the 5th/6th century but demolished in the seventh century and a new and larger longhouse was then built at the same location. A thick lynchet exists close to the longhouse foundations. Finds of Hordeum vulgare var. vulgare (hulled barley), of which some unripe, and weeds within the Late Iron Age house foundation indicate local cereal cultivation (Hansson, 2003; Soltvedt, 1984). Pollen analysis from a bog at the nearby farm site Bøstad, shows a pronounced agricultural phase with Hordeum spp. (barley) starting in the Roman Iron Age and expanding during the Migration Period and Late Iron Age (Vorren, 1986; Vorren et al., 1990). This settlement pattern is consistent with what has been found in other vegetation history surveys near potentially large prehistoric farmsteads on Vestvågøy.
Material and methods
In connection with the establishment of a new suburb area, the eastern part of the heath outfields was divided by a fence in 1991 to keep the animals away. This event gave an opportunity to study the long-term effect of grazing systematically by establishing permanent research plots in corresponding vegetation types on either side of the fence in 1993. The study was designed as a monitoring project with 1993/1994 as year zero. The primary aim was to describe the status of the area in terms of flora and environmental factors, as well as to check whether the 2 years after the establishment of the fence have had a significant impact. The much later opportunity to carry out pollen analysis on a peat core from the local mire, added the ontological and historical aspects to the research focus.
Modern plant ecology
The sampling design is stratified random sampling following Økland (1990) and details are presented in Jensen and Eilertsen (1993) and Jensen et al. (2001). The sampling plots were deliberately laid out in relation to a rich – poor gradient to capture the greatest possible range of variation in the area – limited by the possibility of the grazed-ungrazed parallels. Based on a combination of physiognomic and major floristic characteristics of the local heathland, the following five subjectively selected formations of vegetation became objects of investigation: Empetrum/Arctous heath, Empetrum/Calluna/Kalmia heath, Vaccinium-heath, Nardus-heath and Eriophorum-mire. Within each formation, grazed and not grazed replicats of 100 m2 (10 × 10 m) macroplots were established (Figure 2). Five 1 m2 mesoplots for registration of species abundance were placed randomly within each macroplot, with one mesoplot for soil properties placed adjacent to it. This made a total of 14 macroplots with 70 mesoplots analysed for species abundance and soil properties (Table 1, S1 and S2). The following variables were collected: Presence/absence of species in macroplots, while in mesoplots percentage species cover, pH, P-AL, K-AL, K-HNO3, Mg-AL, Ca-AL at soil-depths of 0–5, 5–20 and 0–20 cm. In addition to the chemical parameters, slope (inclination measured as per mile of 9°) and aspect of exposure of each macro- and mesoplot were recorded by means of a clinometer and compass. In depth interpretation of the environmental results is however not prioritized in the present paper.
Description of analysed macroplots at Haugheia, Vestvågøy.
The species abundance was measured by a percentage scale of 17 cover classes (1, 2, 3, 5, 7, 10, 15, 20 and 30–100). Separate cover registrations were carried out for bottom layer (mosses/lichens), field layer (grasses, herbs, sedges, woody species < 30 cm tall), shrub layer (woody species 30–200 cm) and a tree layer (woody species > 200 cm). Cover and soil properties were recorded at approximately the same phaenological stage in 1993 and 1994, that is, during maximal floristic abundance in July and August. To balance for year-to-year variation, the mean cover of these 2 years is used for the numerical analyses. Standardization of floristic data with respect to identification level was carried out for some species prior to the numerical data treatment to compensate for possible errors in cover index. Sphagnum and Dicranum are hence, presented at genus level and the liverworths Ptilidium ciliare and Stereocaulon sp. are excluded to compensate for eventual misleading cover estimate due to their small size.
The phytosociological status of the heathland is investigated by means of hierarchical, divisive classification of all species data from the mesoplots, using Twinspan two-way species indicator analysis (Hill, 1979; Kent and Coker, 1992). This method combines and weights presence and abundance of a species and is hence suitable for classification according to the Nordic systems of plant sociology originally introduced by Nordhagen (1943), which due to the low number of species is mainly based on constancy, dominance and random occurrences rather than characteristic species. By using seven pseudospecies cut levels (0%, 5%, 10%, 20%, 40%, 60% and 80%) and six divisions the classes may be related to established phytosociological units. The results of the classification are used also for the inspection of homogeneity among replicates and whether grazing impact is a decisive parameter, testing the hypothesis that 1993/1994 may be treated as year zero. The investigation of coenoclines in the dataset was performed using detrended correspondence analysis (DCA) and by a passive canonical correspondence analysis (CCA) of the environmental variables. The chemical variables, except pH, and volume weight are log. transformed as cations are assumed to show a lognormal response curve. Logarithmic transformation makes parameters with different units of measurements comparable, which is why even volume weight is log.transformed. Grazing impact is measured as nominal variables (0–1) and ordinated separately.
Palaeoecology
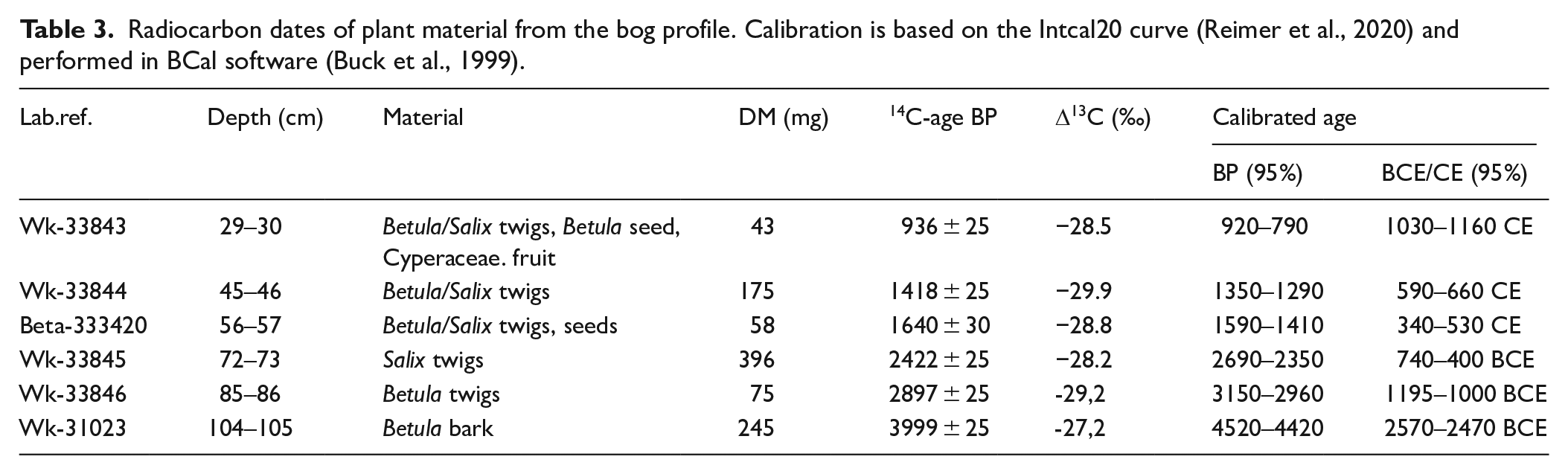
A 114 cm long peat core was sampled from the open grazed mire in 2010, 10–40 m from the mire macroplots by hammering a plastic tube of 10 cm diameter into the ground. The location of the sampling spot was selected to reflect the vegetation of the area as well as possible erosion debris from the grazed, sloping heathland. Generally, every second–third vertical cm was subsampled and a volume of 1 cm3 analysed for pollen, spores and charcoal fragments >20 µm after pretreatment with 10% KOH and acetolysis according to Fægri and Iversen (1989). Two tablets of Lycopodium spores (batch no. 483216, fabricated at Lund University, Sweden) were added prior to the acetolysis as a marker for indirect frequency calculations (Stockmarr, 1971). Loss-on-ignition (LOI) was measured contiguously every cm by drying 3 cm3 peat at 105°C for 24 h and then igniting at 600°C for 6 h. The LOI was calculated as percentages of dry weight. Plant macrofossils for radiocarbon dating (Accelerator Mass Spectrometry) from six selected stratigraphic levels were dried at 60°C for 1–2 days before shipping to Waikato Radiocarbon Dating Laboratory, New Zealand (five samples) and Beta Analytic Inc. (one sample). The 14C-ages were calibrated using BCal (Buck et al., 1999) against the Intcal20 curve (Reimer et al., 2020), and an age-depth model was constructed using linear interpolation in psimpoll 4.27 (Manual for psimpoll and pscomb).
Moss dominated surface samples from within the major vegetation formations were collected and pollen analysed following the same procedure as for the peat samples from the core. The upper 1 cm of the moss cover was sampled at a diameter of 5 cm, producing a volume of 19.6 cm3 per sample and a dry weight between 0.5 and 1.3 g. A total of 30 samples are analysed (Table 2). The pollen sum includes all identified pollen types.
Moss polsters from Haugheia Vestvågøy (HV) sampled during 2010 and analysed for modern pollen content.
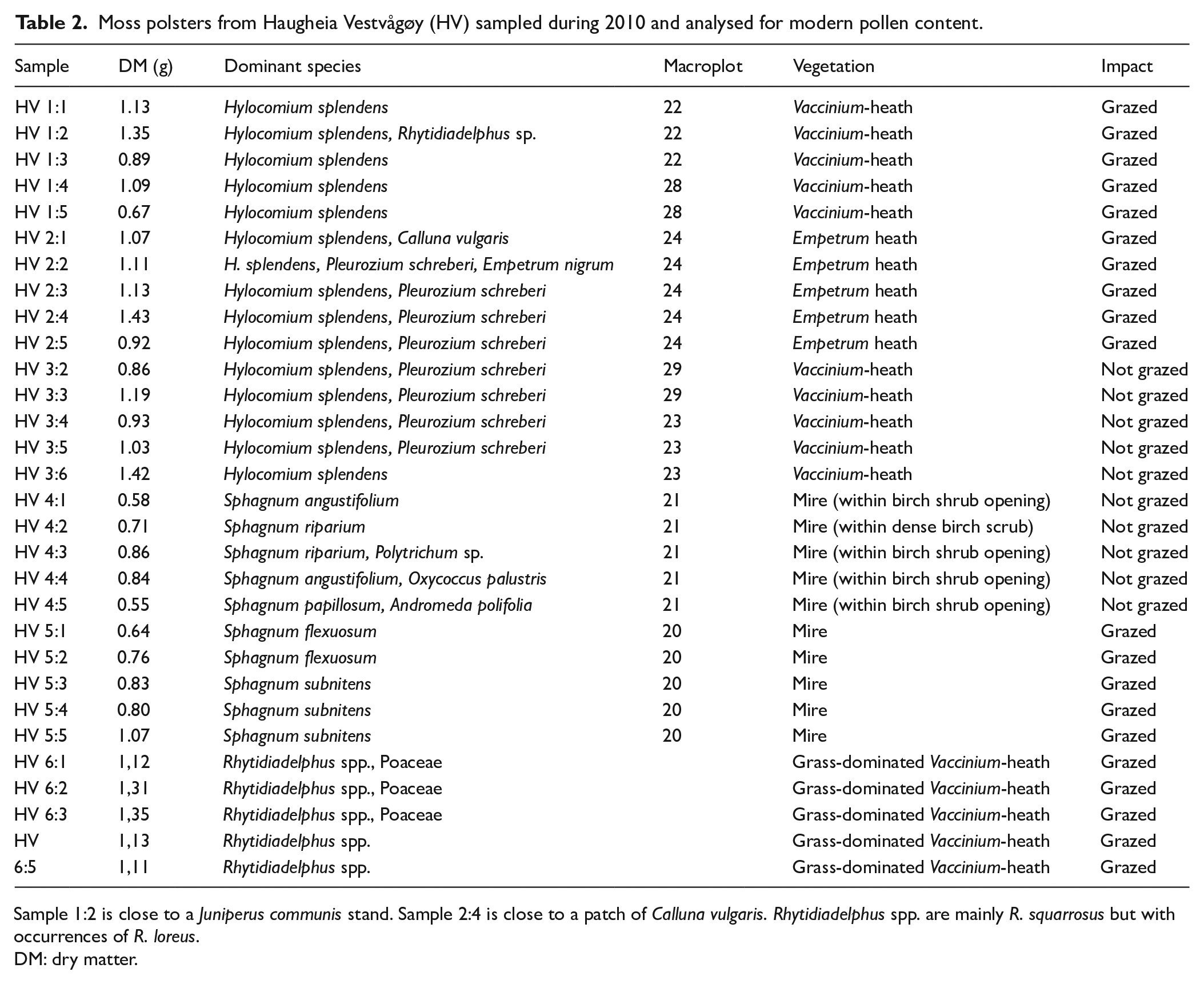
Sample 1:2 is close to a Juniperus communis stand. Sample 2:4 is close to a patch of Calluna vulgaris. Rhytidiadelphus spp. are mainly R. squarrosus but with occurrences of R. loreus.
DM: dry matter.
A DCA ordination of subfossil core samples and modern surface samples is carried out to investigate the major gradients in floristic change on a millennial scale.
Results
Modern plant ecology
The Twinspan classification (Figure S1 and Table S1) divides the dataset of all mesoplots in two major groups (TW 0 and TW 1) based on the presence of Sphagnum species. Group TW 1 consists of all the mesoplots from the mire, in addition to no 27048 belonging to the Nardus-heath formation. This plot contains a high cover (70%) of Sphagnum girgensohni, and as this is the only plot where this species is recorded, it is subsequently excluded from the ordinations because of the effect as an outlier. The heath group (TW 0) is divided based on Potentilla erecta and Anthoxanthum odoratum, and we get two main formations of the heath plots separating the herb-rich (TW 00) from the more exposed heath vegetation rich in dwarf shrubs, mosses and lichens (01). Group TW 00 is further divided in the TW 000 group with >20% cover of Vaccinium myrtillus and presence of Veronica officinalis and the TW 001 group with more than 20% cover of Nardus stricta and >5% Chamaepericlymenum suecicum, Festuca vivipara and Vaccinium uliginosum. Group TW 01 is divided into the TW 010 group with >10% Vaccinium myrtillus and group 011 with <5% Arctous alpina. The mire group (TW 1) is divided into group TW 10 based on >10% of Potentilla palustris and Vaccinium uliginosum and group TW 11 with >10% Polytricum commune. Further divisions with pseudospecies may be seen from the dendrogram in Figure S1. The Twinspan classification helps to sort out the main lines of the dataset as a base for ecological interpretation of the vegetation. According to the Norwegian Biodiversity Information Centre (biodiversity.no), species with a cover of >25% are regarded as dominant and species occurring in >12,5% of the plots as common. In the present context, only species with a frequency > 0.9 (>90%) are considered as common (Table S1). The following vegetation units are consequently outlined for the local heathland based on the selection of groups most representative for the local plant community:
A. Vaccinium myrtillus/Empetrum-heath (TW 0): Vaccinium myrtillus, Empetrum nigrum ssp. hermaphroditum and Hylocomium splendens appears with high frequency and relatively high mean cover. It is separated in two subunits.
A1. Alchemilla alpina/Anthoxanthum odoratum-type (TW 00). The two name-giving species are only present in plots belonging to this group and characterize prealpine coastal pastures of the region. Other species special for this subunit are for example, Campanula rotundifolia, Melampyrum sylvaticum, Veronica officinalis, Viola biflora, Carex pilulifera and Juniperus communis. It may be further separated in two subunits.
A1a. Rumex acetosa-type (TW 000). Rumex acetosa and Achillea millefolium separate this unit from A1b, while Rhytidiadelphus squarrosus and Vaccinium myrtillus are dominant and common species.
A1b. Melampyrum pratensis-type (TW 001). Melampyrum pratensis is the only divisive species towards other units. Empetrum nigrum ssp. hermaphroditum and Hylocomium splendens are dominant and common species.
A2. Calluna vulgaris/Cladonia arbuscula - type (TW 01). Apart from the name-giving species, other common species that are also divisive towards other units are Peltigera canina and Cetraria islandica. It is divided into two subunits.
A2a. Nardus stricta-type (TW 010). This unit has no divisive species towards other units, but the highest mean cover of the common species Nardus stricta and Pleurozium schreberi.
A2b. Arctous alpinus-type (TW 011). A. alpinus as well as Kalmia procumbens and Racomitrium lanuginosum are divisive towards other units. Empetrum nigrum ssp. hermaphroditum, Hylocomium splendens and Pleurozium schreberii are dominant and common species.
B. Sphagnum/Eriophorum mire (TW 1). The bottom layer is dominated by different Sphagnum species, with S. papillosum, S. flexuosum, S. angustofolium, S. subnitens and S. riparium as the most characteristic. Other common species are Lysimachia europaea and Chamaepericlymenum suecicum. It is divided in two subunits.
B1. Tricophorum-type (TW 10). Tricophorum cespitosum is selected as character species, but several other species are divisive towards other units, like Aulacomnium palustre, Carex pauciflora, Drosera rotundifolia, Neottia cordata, Pinguicula vulgaris, Comarum palustre, Oxycoccus palustris, Luzula sudetica and Rhinanthus minor. Empetrum nigrum ssp. hermaphroditum is the only dominant species besides Sphagnum sp., while Eriophorum angustifolium, E. vaginatum, Vaccinium myrtillus and Potentilla erecta are common species.
B2. Carex rostrata-type (TW 11). Apart from C. rostrata, other devisive species are Dactylorhiza maculata, Pyrola rotundifolia, Juncus filiformis, Betula nana and Montia fontana. Shrubs of Betula pubescens are dominant besides Sphagnum sp. while Carex echinata and Polytrichum commune are common species.
With respect to the effect of grazing impact among grazed and not grazed parallells, there is a tendency of separation at Twinspan division level 6. It is however too inconsistent to be interpreted as the primary regulating factor. Further division of the mire group (1) is probably caused by small-scale ecological variation rather than differing grazing impact. Four of the mesoplots belonging to the ungrazed macroplot 21, are grouped together at division level 3 (eigenvalue 0.443) but the species composition reflects homogeneity in humidity and nutritious status rather than grazing impact. Conclusions based on the Twinspan classification are that macroplots 18, 19 and 31 as well as 20, 21 and 30 are sufficiently homogenous to act as parallels for future research. The evident visual grazing effect observed in field on birch leaves and shoots is not confirmed by the registrations of understory species cover. Macroplots 22 and 23 are less homogenous but the splitting happens at a high division level and is most probably random. This is also the case for macroplots 24 and 25, where the deviation caused by a low abundance of Vaccinium myrtillus and a high abundance of Peltigera canina and Kalmia procumbens in mesoplot 25051 may be regarded as result of random variation. Macroplots 26 and 27 are however too heterogenous to act as parallels, as plot 26 shows a drier regime by the presence of lichens like Cladonia spp. and Cetraria spp. together with Calluna vulgaris, probably related to a higher and more exposed elevation. Macroplot 28 and 29 show an internal heterogeneity that may be related to grazing, and it is uncertain if they may act as parallels.
The DCA-ordination of heath-samples (Figure 4) rendered significant variation between axis 1 and 2 (p = 0.01). The results confirm the major vegetation gradient observed in the Twinspan-classification, and subsequently the subjectively selected poor-rich gradient planned by the field survey. Passive CCA of environmental variables shows that inclination (slope) and pH are the parameters that explain most of the variation in the dataset, separating the A1a Rumex acetosa subunit from A2b Arctous alpina, with A2a Nardus stricta and A1b Melampyrum (high Empetrum cover) in an intermediate position. The electrolyths Ca-AL, Mg-AL and P-Al are positively correlated with the A2b Arctous unit, while K-Al is positively correlated with the A2a Nardus unit. A Monte Carlo permutation test using forward selection of only the grazing/not grazing parameter rendered a not significant result (p = 0.19).
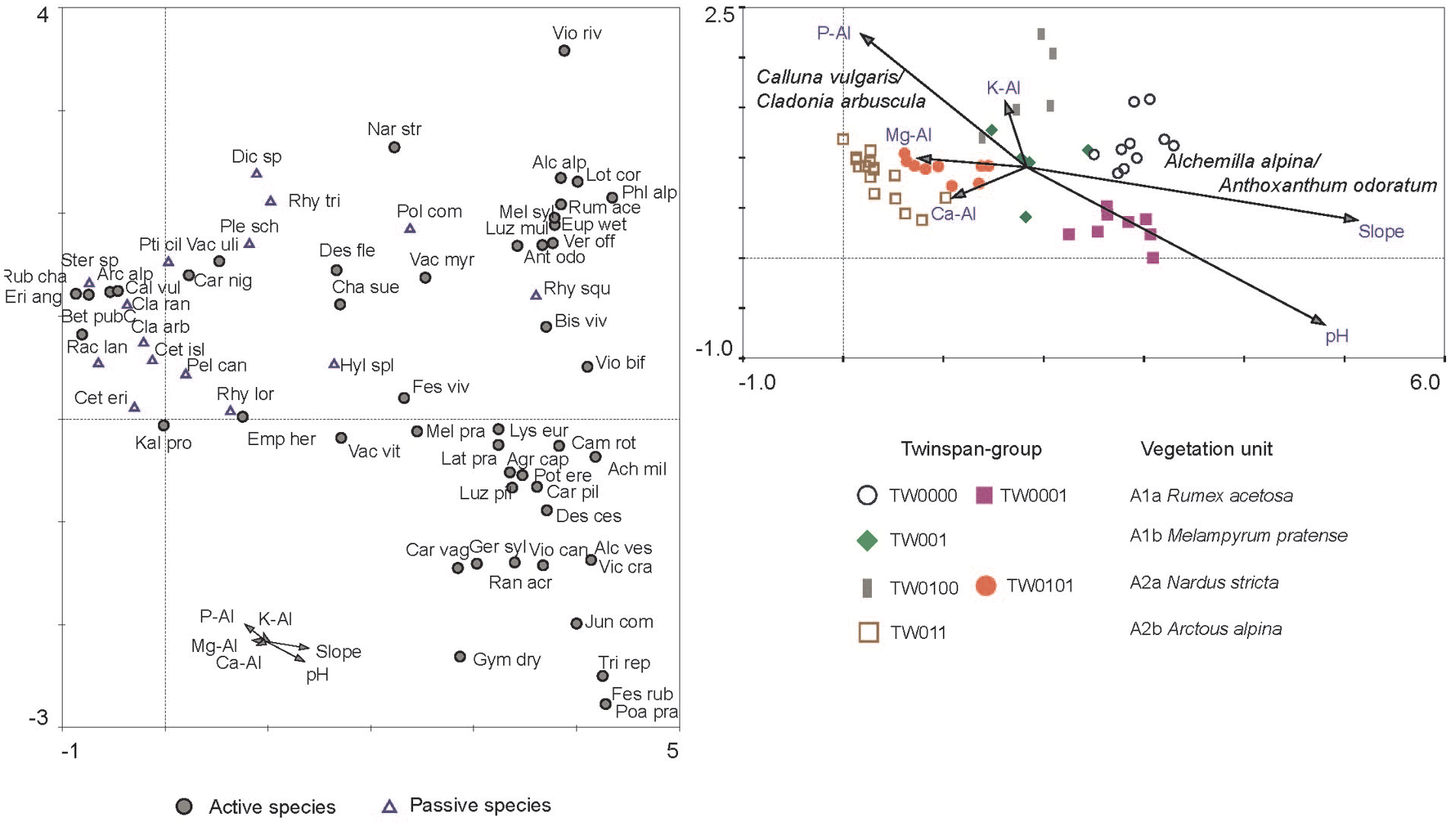
DCA of recent species from monitoring plots with passive CCA of environmental variables. Grazing parameter is not significant (p = 0.19) and not shown in the plot.
A DCA-ordination of samples from the mire plots (Twinspan groups 10 and 11) did not result in a significant variation between the axes as tested by a Monte Carlo permutation test and is not presented.
Palaeoecology
Pollen analysis of the surface moss polsters (Figure S2) shows distinguishing features between the Vaccinium heath, Empetrum heath and mire when it comes to dominant taxa and certain character pollen types. The grass-dominated Vaccinium heath with strong grazing impact (HV 6:1-5) has the highest abundance of pollen from Poaceae (around 60%), moderate values for most other taxa and the only occurrence of pollen from the grazing indicator Trifolium repens-type. The dwarf shrub dominated Vaccinium heath (HV 1: 1-5, HV 3: 6) has 5–25% Vaccinium-type, <10% Empetrum-type and <2% Calluna pollen. Juniperus pollen is best represented in this vegetation unit, as well as tall herbs and fern spores. The highest abundance of Empetrum and Calluna is found in the samples from the Empetrum heath (HV 2: 1-5), up to 30% and 50% respectively. Other taxa are moderately low. The mire samples (HV 4: 1-5, HV 5: 1-5) stand out with the highest abundance of Cyperaceae and Comarum-type pollen and some specific mire taxa like Thalictrum and Menyanthes. Generally, birch pollen values are low. Antropogenic indicators like apophytes and anthropochores are found in all vegetation units, including the mire. The upland vegetation and anthropogenic impact are thus well reflected in the pollen flora of the mire. Coprophilous fungal spores are mainly recorded in the grass dominated Vaccinium heath and Empetrum heath, perhaps as these habitats are frequently used as resting locations by the sheep. Most of the observed Assulina rhizopod are A. muscorum, which exists in dry habitats as well as bogs (Payne et al., 2012). Amphitrema sp. prefers wet habitats and are common in Sphagnum mires as observed in this study. Charcoal is lower than 10% although the site is close to residential areas.
The top 50 cm of the peat core consists of sedge peat of medium degree of humification, H 5–6 of the von Post Humification Scale (FAO, 2011), with scattered elements of small twigs with a diameter of 0.5–2 cm. A distinct presence of sand and gravel is observed within the Cyperaceae-peat at 50–54 cm. Remains of Equisetum and mosses are visible from 54 to 105 cm, as well as larger twigs and pieces of wood and bark. The degree of humification increases markedly from 89 cm (H 7–8) and from 105 cm onwards the peat takes almost full amorphous character (H 9) and contains lenses of sandy silt and dissolved shale rocks. The loss-on-ignition results largely confirm the overall high organic content but with a significant drop around 50 cm and increased mineral content at the bottom of the profile (Figure 5).
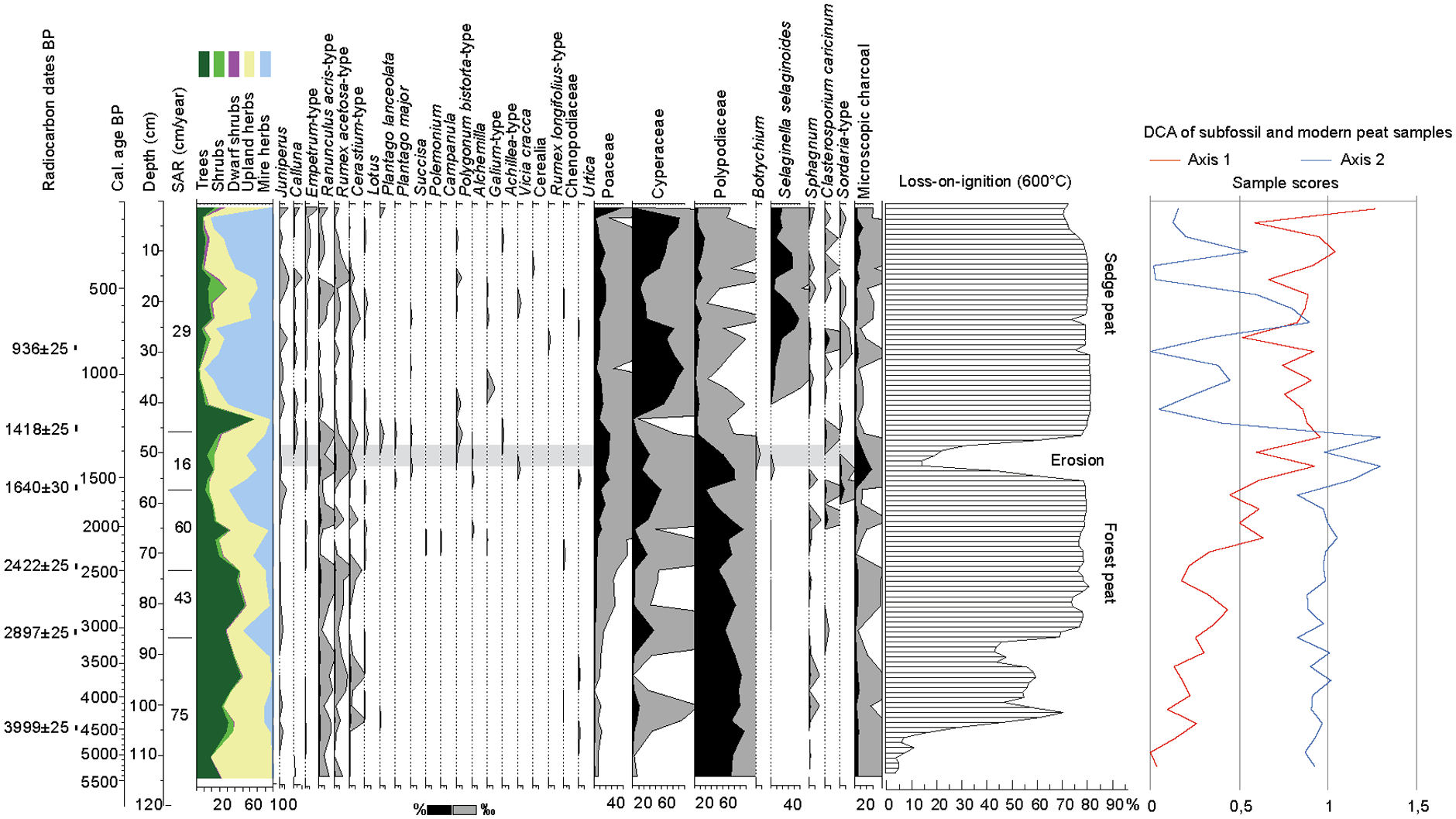
Pollen and sediment stratigraphy of the peat core from Haugheia. Selected pollen and spore taxa, sedimentation rate (SAR) and organic content (Loss-on-ignition) are presented. Sample scores of DCA ordination of fossil and modern pollen data, axes 1 and 2 in the right end of the diagram.
The chosen level of the six radiocarbon dates (Table 3) are based on marked changes in biostratigraphy, and a linear age depth model spanning 5200 years is constructed. The main trend in local vegetation development as displayed by the CONISS cluster analysis, and the change in local mire taxa, is largely synchronous (Figure S3). The following local pollen assemblage zones (LPAZ) are selected, primarily based on the cluster analysis:
HV I (114–72 cm, c. 5200–2500 cal BP, 3200–500 BCE): Betula pubescens-type – Polypodiaceae -Filipendula – Cichorioidae. Birch and ferns dominate this zone, while Filipendula and pollen from Composita Cichorioidae are subdominants. Pollen from several other tall herbs are present, like Valeriana, Geranium and species from the Caryophyllaceae family including Lychnis-type. Low-growing herbs are continuously represented by Melampyrum and Cerastium-type. A stable element of apophytes is seen in the presence of Ranunculus acris-type and Rumex acetosa-type. Salix, Equisetum and Caltha-type are present from c. 105 cm and throughout the zone.
HV II (72–40 cm, c. 2500–1150 cal BP, 500 BCE–800 CE): Poaceae – Polypodiaceae – Betula pubescens-type – Alchemilla. The birch dominance is reduced, and ferns are still well represented but decreasing throughout the zone. Poaceae is increasing, and Alchemilla is present from this zone. The diversity in herb pollen taxa is increased by pollen presumably coming from heath and meadow communities uphill, like for example, Polemonium, Campanula, Galium, Chamaepericlymenum suecica and Lysimachia europaea. A peak in microscopic charcoal in the middle of this zone is likely related to the erosion episode discovered in the sediment and dated to c. 1440–1390 cal BP (510–560 CE) indicated by low LOI-values. Calluna is observed from this level onwards, as well as an increase in pollen taxa related to grazing impact. Such taxa are Plantago major, Plantago lanceolata, Botrychium and fungal spores from the coprophilous genus Sordaria.
HV III (40–0 cm, c. 1150–50 cal BP, 800–50 CE): Cyperaceae – Selaginella selaginoides. This zone is characterized by low influx of pollen from birch and a change in the mire flora. Selaginella selaginoides gainsdominance together with Cyperaceae. Juniperus and Betula nana-type are recorded throughout this zone. Three subzones are outlined.
HV IIIa (40–26 cm, c. 1150–750 cal BP, 800–1200 CE): Cyperaceae – Calluna. Calluna pollen is present in low amounts. Similarly, the concentration of microscopic charcoal is low. Although Calluna is present, the pollen abundance is very low, and this goes also for the microscopic charcoal. Pollen from some apophytic taxa are present in low amounts, and some only sporadic, like Artemisia, Rumex longifolius-type and Brassicaceae.
HV IIIb (26–12 cm, c. 750–350 cal BP, 1200–1610 CE): Salix-Poaceae. An increase in pollen from Salix and tall herbs like Filipendula and Compositae Cichorioideae, together with a decrease in Cyperaceae and Selaginella selaginoides, indicates a regrowth of the mire. Increased input of dwarf shrubs and low herbs may come from hummocks on the mire and thus indicate a beginning ombrotrophy. A general increase in Poaceae and upland herbs suggests that the surroundings of the mire are still utilized as pasture.
HV IIIc (12–0 cm, c. 350–50 cal BP, 1610–50 CE): Cyperaceae – Selaginella selaginoides – Cerealia. The mire is again open and dominated by sedges and Selaginella selaginoides. The grazing impact is not strong as seen from the herb flora, but the presence of a cereal pollen grain and slight increase in microscopic charcoal point to farming activity not far from the site.
Radiocarbon dates of plant material from the bog profile. Calibration is based on the Intcal20 curve (Reimer et al., 2020) and performed in BCal software (Buck et al., 1999).
The DCA-ordination of fossil and modern samples highlights the two major gradients in the palaeoecological development of the site, that is, the change from a forested landscape to an open pasture, and the change in mire regime from Salix and birch charr to sedge mire as demonstrated by the sample scores of axes 1 and 2 (Figure 5). The minerogenic mire develops from 5000 to 4500 cal BP onwards and into a long-lasting phase of birch and willow charr. The surrounding upland consisted of, probably semi-open, birch woodland with a well-developed understory of herbs and dwarf shrubs (Figure S3). From c. 2500 cal BP, there is an incipient opening of the local landscape, which is seen by a reduction in birch pollen, increase in grass pollen and upland herbs. The mire develops into a more sedge dominated stage judged from the increase in Cyperaceae pollen and input of spores from the Carex-infesting fungus Clasterosporium caricinum. A beginning grazing impact may be seen already from around 1500 cal BP by the input of fungal spores from the coprophilous genus Sordaria, followed by a distinct erosion episode around 1400 cal BP, bringing abundant mineral grains, charcoal, and pollen from anthropochore plants into the mire. Calluna is present at low abundance thereafter. A further grazing impact on the mire is seen from c. 1150 cal BP by the increased and continuous presence of Selaginella selaginoides, which tends to benefit from grazing (Elven et al., 2022; Fremstad, 1997). A possible regrowth phase is observed by an increase in willow and tall herb pollen during 750–350 cal BP.
Discussion
This study shows that 2–3 years after cessation of grazing was not long enough time to provide a significant impact on species diversity and coverage at Haugheia. Although seedlings and young leaves of birch and willow on the mire were observed as being a favourite treat for the local sheep, it did not show up in the vegetation analyses. The prediction on year 1993–1994 being treated as year zero was thus supported, and the results from the present investigation may serve as a fundament for future research at this site.
The modern heathland
The sheep roam the entire heath during the day in search of food and shelter. Sheep droppings were found in all parts of the heath. During the fieldwork, it was observed that sheep naturally preferred the most nutritious grass- and forbs dominated uplands as well as the lush Vaccinium myrtillus heath and sedge mire with birch and willow shrub. The scantier parts of the heathland, often with boulders suitable for shelter, were much used as resting areas. A possible disadvantage of the experimental design is that the heathland along the uppermost part of Haugheia was not included, as this grass-dominated Vaccinium-heath with Alchemilla alpina probably has been most exposed to grazing. The reason for exclusion was the lack of ungrazed replicates for this vegetation type. It is thus interesting that the pollen deposition in the surface samples collected from this type of heath, stands out markedly by having a clear grass dominance and presence of coprophilous fungal spores (sample 6: 1–5 in Figure S2). The steep slope with erosion scars (Figure 3b) facilitates the input of palynomorphs to the mire below. Pollen from herbs related to grazing is present in all surface samples (Figure S2). This shows that the local grazing impact is possible to trace historically in pollen analysis of sediments, as demonstrated in this study (Figure 5 and S3).
The strong cultural influence makes it difficult to place the heath and mire communities of Haugheia in an established plant sociological system. The close vicinity to residential areas has for example, caused the establishment of Rhytidiadelphus squarrosus even to the mire, and the droppings from sheep roaming all over the heathland helps the spread of species. From a plant sociological point of view, it would be more appropriate to consider the fine scaled grouping of plants in the vegetation classification mainly as ephemeral elements. Heath and mire easily merge into each other in the strongly oceanic environment, and the extent and biological characteristics change over time with changes in climate as well as grazing impact. Some environmental factors are however stable over long time spans, like the edaphic conditions related to the bedrock and orographic aspects influencing light, temperature and water level together with the climate. Overall, the recorded flora of Haugheia (Table S3) is northern boreal with a few middle boreal species, like Anthoxanthum odoratum, Juncus effusus and Anemone nemorosa, although the latter may be an anthropochore this north (Elven et al., 2022).
The mire consists of a mixture of minerotrophic areas and low hummocks and lawns showing an incipient ombrotrophy and regrowth by shrubs. As seen from the peat core stratigraphy, this mire developed over mineral soil and parts of the vegetation has maintained contact with groundwater until present. The small-scale topography of the Haugheia mire consists of <50 cm tall hummocks with several species from the nearby heath vegetation, like Empetrum nigrum ssp. hermaphroditum, Vaccinium species, Betula nana, Chamaepericlymenum suecica and Lysimachia europaea as well as shrubs of Betula pubescens and Salix lapponum. These species are recognized in the numerical classification as common species for the heathland, while many of the species classified as the Tricophorum and Carex rostrata sub-groups are from the lawn and minerotrophic carpet or mud-bottom elements of the mire. The pH range of the mire plots is 4.1–4.9 and the acidic character is seen in species like Sphagnum papillosum, Tricophorum cespitosum, Eriophorum angustifolium, Carex rostrata and outside the plots also Narthecium ossifragum, a typical species of oligothrophic Atlantic mires. A few species, like Comarum palustre and Salix lapponum, indicate mesotrophy. This is a common type of mire in the lowlands of Vestvågøy. The species composition has many features in common with other local mires, such as Ålgreva at Rystad (Vorren et al., 2012) and Dønvold near Leknes (Edvardsen, 1988). Survey transects in the eastern part of the Dønvold mire show a transition from poor dwarf shrub dominated heath to oligotrophic-mesotrophic mire with several floristic attributes that are recognizable at Haugheia.
The Vaccinium/Empetrum heathland at Haugheia has some similarities with the units Empetrum-Calluna with Vaccinium and Empetrum-Calluna with grass and herbs found in a classification of lowland heath vegetation at four sites along the coast of Nord-Trøndelag and Nordland County (Tveraabak, 2004a, 2004b). These two units are dominant at the northernmost site Austvågøy in eastern Lofoten and explained as result of low winter temperatures influencing the growing season, snow cover and accumulation.
The long-term anthropogenic and climatic impact
There seems to be a correlation between the development in the mire and local anthropogenic activity at Haugheia. The reduction in birch and increase in grass from c. 2500 cal BP is followed by an input of species benefitting from grazing and fertilization by animals. The erosion event around 1400 cal BP may have been triggered by intensified exploitation of the upland and the mire. The synchronous peak in charcoal points to burning episodes in the surrounding upland of the mire. Calluna pollen is recorded from this event onwards but in low values. A temporary increase in birch pollen occurs soon after this event, followed by persistently low birch and fern values and increased Cyperaceae up to the present. The change in mire regime is probably linked to increased water level because of removal of trees. Selaginella selaginoides gains dominance, which may be an effect of increased grazing of the sedge mire. The heathland, including the mire, may have been utilized by a combination of chopping, grazing and possibly occasional burning, maintaining an open landscape from c. 1500 years ago until present. However, a period of regrowth is visualized in LPAZ HV IIIb and tentatively dated as between c. 750 and 350 cal BP (Figure 5 and S3) which overlaps with the time of destruction after the Black Death in the mid-1300s, when more than 80% of farms on Vestvågøy was abandoned (Nielsen, 1977).
The nearby Dønvold mire experienced a reduction in the local birch forest with a concomitant increase in grasses and apophytes and the first charcoal observations c. 3500 cal BP (Nilssen, 1983). This development is later markedly enhanced at a stratigraphic level dated as 2760 ± 110 BP (3210–2700 cal BP) when the first Hordeum-type pollen is recorded. A clear anthropogenic signal in the pollen record is thus observed earlier at Dønvold than at Haugheia, where a comparable development is seen around 2500 cal BP. An increase in Ericaceae and decrease in Filipendula dated as 1170 ± 60 BP (1200–1100 cal BP, 680–990 CE) is interpreted as an accelerated development towards heathland at Dønvold, with a later marked and lasting increase also in Calluna and Rubus chamaemorus, while the mire has undergone ombrotrophy. The change towards Cyperaceae dominance around 1150 cal BP in the Haugheia mire, is accompanied by a low but steady presence of Betula nana-type and other dwarf shrubs, which may be interpreted as a further development towards heathland. Empetrum-type pollen is observed already at c. 4500 cal BP but more frequently from c. 2500 cal BP, implying that Empetrum nigrum has been a natural part of the local flora in the long run. A possible human triggering of heath expansion by pastoral farming is seen in lake sediments of eastern Lofoten from 200 CE, followed by an increase in Calluna pollen and microscopic charcoal interpreted as a possible increased human impact (Tveraabak, 2004a, 2004b). The representation of Calluna in the pollen diagram from Haugheia is however low, and a causal relation between Calluna and human activity is not obvious. Calluna vulgaris is commonly found as patches of local dominance within the Empetrum heaths in Lofoten and Vesterålen and may be the dominant vascular species of ombrotrophic bogs in the region. Calluna vulgaris is at its northwestern biogeographical limit in this area, and sensitive for local climatic variations, particularly summer temperature, and grazing impact. Distinct charcoal layers were not recorded at Haugheia, but the area was not systematically investigated for this. Regeneration rates of Calluna vulgaris as well as towards the pre-fire community are found to be faster in south, benefitting from a higher productivity rate and larger pools of pioneer species (Vandvik et al., 2014; Velle and Vandvik, 2014). Empetrum nigrum does not benefit from burning as a regeneration trigger like Calluna vulgaris and is kept at low cover during repeated high intensity burning. When burning ceases, Empetrum nigrum may gain dominance over Calluna vulgaris, also helped by a low preference from herbivores and the allelopathic influence on other plant species and soil biota, by releasing phenolic compounds from the leaves (Bell and Tallis, 1973; Edvardsen et al., 1988; Nilsson et al., 1998; Odén et al., 1992). The ongoing increase of Empetrum nigrum in boreal heathland is observed as a regional phenomenon. Bråthen et al. (2024) did in 2020 a resurvey of 139 coastal heathland sites north of 69.6°N, originally surveyed by Matti Haapasaari during 1965–1975 (Haapasaari, 1988), and showed a significant change in species dominance pattern. Haapasaari defined six heathland types, each with >25% cover of their characteristic dwarf shrubs Calluna vulgaris, Kalmia procumbens, Betula nana, Vaccinium myrtillus and Empetrum nigrum. Bråthen et al. (2024) found that except for the Betula nana heathland type, all had an increase in Empetrum nigrum cover, and the species richness was stagnant or reduced.
The development observed at Haugheia is probably reflecting an expansion in agricultural activity all over the Vestvågøy region, with increased exploitation of outfields. It has not been possible to allocate the development to a special farm because of the lack of archaeological excavations. Heathland development with Calluna vulgaris and grass dominance is documented from 1500±70 BP (1530-1300 cal BP, 420–650 CE) in the Bøstad peat core (Vorren, 1979, 1986) near Borg. This coincides well with the timing of the first house foundation at Borg (Munch et al., 2003). The first farm at Bøstad was however established sometime between 2770 and 2300 cal BP (800–350 BCE, 2430 ± 130 BP) when clearing of birch, development of grassland and a contiguous presence of Hordeum-type is evident in the pollen diagram (Vorren et al., 1990). At Rystad c. 1.5 km SW of Bøstad, two major agricultural phases are visible in the pollen and microscopic charcoal records; one commencing in the Pre-Roman Iron Age (2340 ± 70 BP, 2360 ± 60 BP) and lasting for c. 400 years and the next during Late Roman Iron Age with apparently contiguous agricultural impact of varying intensity until present (Vorren et al., 1990, 2012). Biochemical analyses of sediment cores from the lake adjacent to Rystad have revealed an increase in faecal sterols and other biochemical compounds from human and animal faeces at c. 2250 cal BP, and a further increase after 1900 cal BP, reaching a maximum c. 1450 cal BP (500 CE), which is interpreted as the result of maximum Iron Age populations of humans and grazing animals (D’Anjou et al., 2012). Periods of very low input and consequently low impact from humans and livestock happened from c. 1300 cal BP (650 CE) and accelerated at 1100 cal BP (850 CE; op.cit.). Topness et al. (2023) investigated the content of charcoal and PAHs (incomplete combustion products) in three lakes around the Borg site. They found a general increase in local fire activity over the past 5000 years, with much higher values during the period c. 2400–1500 cal BP (Pre-Roman Iron Age until Migration period) and from 900 cal BP (Early Mediaeval Age) until present time.
Altogether, the human impact on sites at Vestvågøy as observed from botanical, sedimentary and chemical records match well, and the results from Haugheia fits well into this picture. A possible coeffect between regional climate deterioration and increased local grazing by livestock is discussed at a regional scale by Jørgensen (2020) and by Vorren et al. (2012), the latter based on distinct wet-shifts (abrupt changes from high to low peat decomposition) in the Rystad cores c. 2600 cal BP, 2300–2200 cal BP and 1600–1500 cal BP. These events do not match precisely with the dated biostratigraphy of Haugheia. Although it may be explained by methodological bias, the most reasonable explanation is that the Haugheia record mainly reflects local anthropogenic events.
Conclusions
A detailed, combined phytosociological and palaeobotanical investigation of the semi-natural Haugheia heathland in Lofoten resulted in novel information about the floristic composition, origin and development of coastal, anthropogenic heathland at its northern boundary.
The status of the heathland as a semi-natural ecosystem developing from long-term anthropogenic influence is confirmed by the vegetation history. Clearing of pristine woodland followed by grazing through c. 1500 years has maintained the open landscape. Regular burning has probably not been a part of the maintenance judged from the low abundance of Calluna pollen and microscopic charcoal, but sporadic burning especially in the initial phase, may have been the case. Empetrum nigrum has been a steady and increasing component of the heathland, possibly because of the grazing impact and lack of high intensity burning of the heathland.
Two years after cessation of grazing, no significant impact on species diversity and coverage was observed in the survey plots. The floristic results from this study thus make a robust base for future investigations on grazing impact at the site.
The location in a bioclimatical transition zone is reflected in the composition of the modern flora, which is northern boreal with distinct middle boreal elements. The Vaccinium myrtillus/ Empetrum nigrum-heath is classified in two main units; an Alchemilla alpina/Anthoxanthum odoratum-type and a Calluna vulgaris/ Cladonia arbuscula-type. The Sphagnum/Eriophorum-mire, which is in close connection with the heath, is classified in a Tricophorum-type and a Carex rostrata-type reflecting minerotrophy, but with ombrotrophic hummocks containing dwarf shrubs and shrubs. The anthropogenic influence, primarily grazing by sheep, is observed in the flora of the entire heathland, inclusive the mire. This is also evident in the pollen flora, as observed from analysis of surface moss polsters.
An incipient possibly anthropogenic, opening of the local birch woodland happened from c. 2500 cal BP. Intensified exploitation of the heathland by grazing is observed from 1500 to 1400 cal BP when clearing of trees on and around the mire probably lead to increase in groundwater level and an erosion event. The mire develops into an open sedge mire shortly after. The open character of the heathland has remained until modern time, apart from a period of regrowth c. 750–350 cal BP, possibly because of abandonment following the Black Death. The anthropogenic influence on the local heathland is compliant with archaeological and palaeoecological studies from the region, stating high levels of Iron Age population density and farming activity. It is evident that woodland clearing, and a persistent grazing impact has contributed significantly to the formation and maintenance of the heathland as a semi-natural ecosystem.
Supplemental Material
sj-docx-4-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-docx-4-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Supplemental Material
sj-docx-5-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-docx-5-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Supplemental Material
sj-docx-6-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-docx-6-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Supplemental Material
sj-pdf-1-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-pdf-1-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Supplemental Material
sj-pdf-2-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-pdf-2-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Supplemental Material
sj-pdf-3-hol-10.1177_09596836251366188 – Supplemental material for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway
Supplemental material, sj-pdf-3-hol-10.1177_09596836251366188 for Phytosociology and long-term succession of a 1500 year old coastal heathland pasture in Lofoten, northern Norway by Christin Eldegard Jensen in The Holocene
Footnotes
Acknowledgements
I would like to thank Karl-Dag Vorren, Ragnar T. Samuelsen and Svein Morten Eilertsen for a rewarding collaboration in the NRC-project Cultural landscape and re-growth in coastal northern Norway, and Karl-Dag Vorren also for kindly assisting during peat coring. The valuable comments from an anonymous reviewer helped improve the manuscript, for which I am very grateful.
Correction (October 2025):
Article updated online to correct “A central minerotrophic mire under ombrotrophy development is classified in a Tricophorum- and a Carex rostrata-type with hummocks containing dwarf shrubs and shrubs. Pollen analysis of this mire reveals an incipient possibly anthropogenic, opening of the local birch woodland from c. 250 cal. BP” to “A central minerotrophic mire under ombrotrophy development is classified in a Tricophorum- and a Carex rostrata-type with hummocks containing dwarf shrubs and shrubs. Pollen analysis of this mire reveals an incipient possibly anthropogenic, opening of the local birch woodland from c. 2500 cal. BP” in the Abstract section.
Author contributions
Christin Eldegard Jensen: Conceptualization, Data curation, Formal analysis, Methodology, Resources, Writing – original draft, Writing – review & editing.
Funding
The author disclosed reciept of the following financial support for the research, authorship, and/or publication of this article: The vegetation survey in 1993-1995 was financed by The Norwegian Research Council, on project no. 266.729 103270/720 ‘Cultural landscape and re-growth in coastal northern Norway’. It was a co-operation between Holt Research Centre, The Norwegian Research Institute (now NIBIO Norwegian Institute of Bioeconomy) and the University of Tromsø, Department of Biology and Geology (now UiT The Arctic University of Tromsø). Holt Research Centre was scientifically responsible for the sampling and analyses of soil parameters, while the University of Tromsø was responsible for the sampling and analysis of floristic data. The peat coring and palaeoecological study in 2010 was financed by Museum of Archaeology, University of Stavanger.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.