
Abstract
Plant macrofossil and pollen records from the partially varved sediments of Lake Odensjön in southernmost Sweden provide detailed information on local forest development and land-use history within Söderåsen National Park during the past 600 years. A relatively dense forest dominated by Fagus (European beech) covered the steep talus-dominated slopes surrounding the lake during at least a century prior to c. 1550 CE, when a change towards more open conditions occurred in response to intensified grazing. The Fagus population on the slopes was at its minimum at c. 1700–1900 CE, when high Juniperus pollen frequencies indicate the presence of grazed heathlands on the horst plateau above. Reduced grazing pressure caused renewed and rapid forest expansion after c. 1900 CE, which resulted in development of the Fagus-dominated forest with a homogenous age structure that characterises the slopes near the lake and large parts of the park today. This dynamic forest history is supported by dendrochronological data from the local Fagus population and information from historical maps, documents and aerial photographs. It is also consistent with palaeoecological evidence from Fagus-dominated forests in other agriculturally marginal areas in southernmost Sweden, suggesting that these were shaped by a similar land-use history. Therefore, the recent and rapid forest development revealed in this study has implications for management strategies aiming at conservation of Fagus-dominated forest in similar settings in this part of Sweden and neighbouring countries.
Keywords
Introduction
The estimated proportion of Fagus sylvatica (European beech) trees in South Swedish forests is on average only 2.5% (Swedish University of Agricultural Sciences, 2024). This region, which forms part of the northern limit of the natural range of Fagus, is characterised by coniferous forests (dominated by Picea abies and Pinus sylvestris) in the northern and inland areas, while coastal and lowland areas are characterised by agricultural land and broadleaved forests (dominated by Betula pubescens, Betula pendula, Quercus robur, Quercus petraea, Alnus glutinosa, Populus tremula, Fagus sylvatica, Salix caprea and Fraxinus excelsior). However, forests dominated by Fagus occur locally, and their sometimes almost monospecific composition is largely explained by the high shade-tolerance of Fagus and its ability to shade out other tree taxa (Houston Durrant et al., 2016). The resulting limited shrub-, field- and ground-layer vegetation gives these forests a distinctive open character that attracts many visitors. Together with a unique flora and fauna, this explains why Fagus-dominated forests are relatively well-represented in South Swedish nature reserves and national parks.
As revealed by pollen data, Fagus reached southern Sweden around 4000–3500 cal BP after migrating northward from glacial refugia in southern Europe, and its maximum northern distribution in Sweden was attained around 1000 cal BP (Björkman, 1999). Subsequently, the distribution of Fagus has become more fragmentary. This is usually attributed to a combination of human exploitation of Fagus trees and competition from Picea, which was spreading southwards into southern Sweden during the Late-Holocene (Björkman, 1996). In the beginning of the 20th century, when the influence of forestry on tree distribution was still limited, Picea had reached as far south as the north-easternmost part of the province of Scania (Hesselman and Schotte, 1906; Figure 1). Like Fagus, Picea is a shade-tolerant and dominant species, and Fagus appears to have been outcompeted by Picea in most areas. However, Fagus-dominated forests exist locally today within the region of conifer forest dominated by Picea. As suggested by palaeoecological evidence, the persistence of Fagus populations in these isolated areas depended on recurring disturbance from fire events (natural or human-induced) and land-use activities (e.g. selective cutting, forest grazing, and agriculture) (Björkman, 1999; Lindbladh et al., 2008).
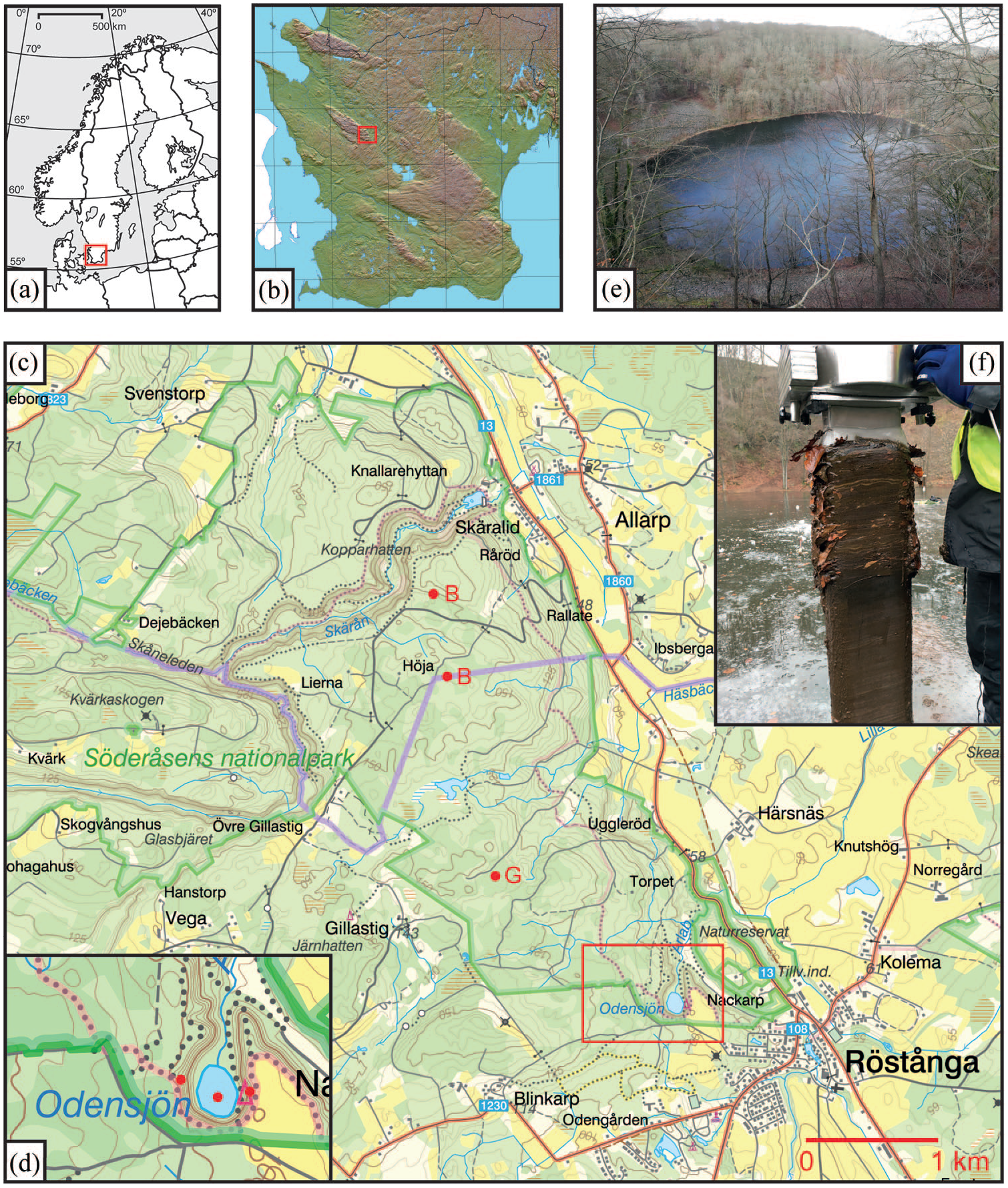
The setting of Lake Odensjön. (a) Overview map of Scandinavia and neighbouring countries with the area covered by panel (b) marked by the red rectangle. (b) Topographic map of the province of Scania showing the NW-SE-trending horst ridges characteristic of this region, with the area covered by panel (c) marked by the red rectangle in the south-eastern part of the Söderåsen horst. The northern border of Scania is marked by the black line. © Lantmäteriet. (c) Topographic map of the north-eastern margin of the Söderåsen horst NW of Röstånga with the position of Odensjön in the south-eastern corner of Söderåsen National Park. The positions of the peatland sites studied by Göransson (1999) and Bergman (2000) are marked by the letters B and G in red, respectively, and the area covered by panel (d) is marked by the red rectangle. © Lantmäteriet. (d) Enlargement of the area around Odensjön with the lake and tree coring sites marked by red dots. © Lantmäteriet. (e) View across Odensjön from the southern plateau edge in February 2020 showing talus without tree cover on the lower western slope to the left, and the outlet area in the north. (f) The upper part of the analysed freeze core from Odensjön with the conspicuous increase in Fagus leaves visible at c. 0.26 m sediment depth.
The presence of Fagus-dominated forests outside the natural range of Picea in the southernmost part and coastal areas of southern Sweden is also closely related to historical land use (Björkman, 1997, 2001; Hannon et al., 2018, 2022; Hultberg et al., 2010; Lindbladh et al., 2008). Some of these forests are found in agriculturally marginal areas used as pasture until the mid-19th century, where subsequently decreased grazing pressure allowed Fagus to rapidly expand and become locally dominant. For example, this process of forest development is historically documented from the horst ridges dominated by Proterozoic gneiss bedrock that constitute a characteristic feature of Scania, the southernmost province of Sweden (Persson, 1971; Sjöbeck, 1927; Figure 1). This is explained by their rugged topography and less fertile soils with frequent stones and boulders, making them relatively unsuitable for crop cultivation. However, the distribution of Fagus-dominated forest is much restricted in these areas today because of replacement by Picea plantations during the last c. 150 years.
Here we explore the history of Fagus-dominated forest within Söderåsen National Park, which covers an area of c. 16 km2 of the Söderåsen horst ridge in NW Scania, characterised by a spectacular landscape with deep fissure valleys and Fagus-dominated forests that in many areas are mature and have a homogenous age structure (Figure 1). Other types of broadleaved forest are also found, mainly Alnus forest (along streams and in wetlands) and Betula forest, and Picea plantations covered relatively large areas when the park was established in 2001 (Naturvårdsverket, 2001). According to the management plan, natural vegetation is to be left largely untouched, while areas of cultural importance are to be preserved through active management. Areas with alien vegetation or previously used for forestry (Picea plantations) are to be restored, primarily as broad-leaved forest (Naturvårdsverket, 2001). Although information about past land use from historical records was considered when management strategies and methods were developed, palaeoecological records providing a long-term perspective on forest development were not available until around the time of establishment of the park (Bergman, 2000; Göransson, 1999). New palaeoecological records with high temporal resolution are therefore important to inform management of the Fagus-dominated forests within the national park, and for understanding the development of their unique flora and fauna.
In this study, we analyse plant macrofossils and pollen preserved in the uppermost (younger than c. 1450 CE) and partially varved sediments of Lake Odensjön (Figure 1). Because of high accumulation rates and a well-constrained chronology, these sediments offer a unique opportunity to determine in detail when the Fagus-dominated forest characterising large parts of the park were formed and identify the conditions and processes responsible for their development. Our interpretations of the palaeobotanical records are compared with independent tree establishment data obtained from dendrochronological analyses of Fagus trees growing around the lake, and the resulting reconstruction of local forest development is validated against information from historical maps, documents and aerial photographs. We also compare our reconstruction with palaeoecological evidence from sites in similar settings within southernmost Sweden to identify any common patterns in forest development. The results of our study have implications for forest management, not only within the park but also in other Fagus-dominated forests in southernmost Sweden and neighbouring countries with comparable topographic, geological and climatic setting.
Material and methods
Site description
Lake Odensjön (56°0′14″N, 13°16′32″E, 60 m.a.s.l.) is situated at the head of a narrow valley incised into the north-eastern margin of the Söderåsen horst ridge approximately 1 km WNW of Röstånga in the north-western part of the province of Scania, southernmost Sweden (Figure 1). The lake, which is almost circular in shape and has a diameter of c. 150 m, is bordered by steep talus-dominated slopes except in the north, where a small stream leaves the c. 20 m deep lake and continues northwards down the valley. The lake is fed mainly by groundwater, but water is also seasonally supplied through a few very small streams draining <0.1 km2 of the neighbouring horst plateau situated 30–40 m above the lake. Mean annual values (1991–2020) of temperature and precipitation at Ljungbyhed, located on the plain below Söderåsen, 8 km NNW of Odensjön, are 8.2°C and 839 mm, respectively (Swedish Meteorological and Hydrological Institute, 2025). Corresponding mean values are 0.2°C and 71 mm for January, and 17.3°C and 82 mm for July.
As for most parts of the Söderåsen horst, the bedrock in the immediate surroundings of Odensjön is dominated by Proterozoic gneiss, but Cambrian sandstone and alum shale also occur within a 1 km distance to the south (Ringberg, 1984). Gneiss bedrock is exposed at the highest levels around the lake. The steep talus slopes below are, especially in their lower parts, characterised by angular stones and boulders formed through frost weathering (Figure 1). Sandy till dominates the plateau, but clayey till is found in areas with sedimentary rocks. Fen peat occurs in the outlet area (Ringberg, 1984).
Due to its spectacular setting, rounded shape and great water depth, there are several myths connected to Odensjön and different theories about its origin. According to Rapp (1984), the lake basin was shaped through recurrent erosion by local cirque glaciers during the Weichselian glaciation. This erosion is believed to primarily have occurred during periods of tundra conditions before the main advance of the Weichselian ice sheet across this part of Sweden, culminating around 20,000 cal BP. This ice advance is assumed to have left the topography of the lake basin and adjacent valley unaltered, but cirque glaciers may have reappeared and continued to shape the lake basin during cold intervals of the Late Weichselian (Rapp, 1984). A pollen-stratigraphic record from Odensjön was presented by Berglund and Rapp (1988), but this only covers the Late Weichselian part of the sediment sequence and therefore provides little information relevant to the development of the present Fagus-dominated forest.
The slopes surrounding the lake are in their upper parts characterised by closed forest dominated by Fagus sylvatica. The forest on the lower slopes is more open and diverse, with Tilia cordata, Corylus avellana, Carpinus betulus, Betula pendula, Betula pubescens and Quercus petraea, but trees are absent from some of the lower talus areas (Figure 1). Closed Fagus-dominated forest dominates the immediate surroundings on the plateau above the slopes, but Picea plantations are found as close as c. 100 m W of the lake, and a cultivated field is located next to the plateau edge on the opposite side. The fen in the outlet area is covered by Alnus glutinosa, Betula pendula and Betula pubescens.
Field and laboratory methods
The uppermost 0.91 m of sediments were collected with a freeze corer (Renberg and Hansson, 2010) from lake ice in the deepest part (19.9 m) of Odensjön in February 2017. A conspicuous increase in Fagus leaves at c. 0.26 m sediment depth was noted already in the field (Figure 1), suggesting a recent and major expansion of Fagus close to the lake. Moreover, the core displayed potentially annual laminations (varves). By combining counting of these laminations with 210Pb dating of the uppermost 0.33 m of the sediments and 14C dating of leaves (Fagus and Betula) from three levels between 0.68 and 0.88 m, Hertzman (2021) confirmed that the sediment sequence is partially varved. The chronology used here, which is constrained by these 210Pb and 14C data, was developed and described in detail in a recent study focussing on the development of the diatom community of the lake (Silvester et al., 2025).
A 2 cm wide segment spanning the uppermost 0.86 m of one side of the freeze core was cut out using a band saw and subsequently divided with a hand saw in a walk-in freezer into contiguous samples for pollen analysis measuring 2 × 2 × 2 cm. The sample containing the conspicuous increase in Fagus leaves at c. 0.26 m was split into two 1-cm thick subsamples, which resulted in a total of 44 samples. Of these, 28 were selected for analysis and prepared by cutting away the innermost parts showing evidence of varve distortion caused by downward penetration of the corer. The remaining parts were left to melt and thoroughly mixed before transferring 2 cm3 of representative material with a syringe to test tubes. Following standard chemical treatment for pollen analysis (Faegri et al., 1989), microscope slides were then prepared and screened at 400× magnification. Identification was supported by literature (e.g. Faegri et al., 1989), online databases (PalDat, 2020; The Global Pollen Project, 2020) and a pollen reference collection (Department of Geology, Lund University). Grass pollen were classified based on size criteria as either Poaceae (<40 µm) or Cerealia (>40 µm). Approximately 500 pollen grains were counted in each sample. Percentage calculations were based on the sum of terrestrial pollen. For taxa belonging to other groups (ferns, clubmosses, aquatic plants, and mosses), and for charcoal fragments, percentages were based on the combined sum of terrestrial pollen and the total count of the respective group.
The remaining part of the freeze-core side was cut into samples for plant macrofossil analysis, spanning the same depth intervals as the pollen samples, and their volumes were estimated by water displacement. After careful washing through a 0.50 mm sieve, the residue of each of the 44 samples was inspected in a petri dish under a stereo microscope at 75× to 625× magnification. Identification was supported by literature (Birks, 2007; Cappers et al., 2012; Mauquoy and Van Geel, 2007), a reference collection of seeds and fruits (Department of Geology, Lund University), and a reference collection of tree leaves and buds constructed from material collected on the ground at Odensjön in February 2020. Leaf fragments smaller than c. 2 mm were counted as unidentified, and pieces of Fagus leaves >⅓ of a whole leaf were counted separately. The analysis focused on tree macrofossils, but twigs, charcoal particles and Nymphaea alba seed fragments were also counted. Stems and leaves of Sphagnum mosses were very abundant in some samples, which made quantification difficult. These remains were therefore categorised as either present (⩽20) or abundant (>20), while concentration values were calculated for all other taxa.
To estimate the age of the local Fagus population, wood cores were collected with an increment borer in May 2020 from trees considered to be among the earliest established based on size criteria. Because the permission granted only allowed coring of ten trees, five trees within two small areas on the upper slopes on opposite sides (W and E) of the lake (Figure 1) were selected to provide as representative data as possible. To obtain cores spanning the full lifetime of trees, coring was made at breast height (c. 1.4 m) on the uphill side of trunks lacking evidence of growth disturbances. The surfaces of the wood cores were prepared by cutting with an industrial razor blade, after which talcum powder was applied to the cell cavities to enhance the contrast between the ring boundaries. Subsequently, the annual rings were counted and analysed using standard dendrochronological procedures and equipment (Bräker, 2002). Tree-ring widths were measured with a precision of 0.01 mm, using a digital positioning table connected to a stereomicroscope with 10× to 60× magnification, and analysed with the TSAPWin Scientific software (Rinn, 2003). Conventional cross-dating techniques (Cook and Kairiukstis, 2013) were used for the development of a tree-ring chronology, identification of potential measurement errors or missing rings and for dating individual trees. The quality of the cross-dating, the measurements and the tree-ring chronology was evaluated using the COFECHA software (Holmes, 1983). When a core did not contain the pith, an estimated number of missing rings was added based on fitting of transparent concentric circles to the top of the core (Bollschweiler et al., 2008). Thereafter, 10 more years were added to account for the estimated time for the trees to grow up to the level of coring. To enable for studies of climate- and environment-related growth variations in the trees, the tree-ring data were standardised to minimise influences related to the age trend of the trees (Bunn, 2008).
Results
Plant macrofossil data
The macrofossil record displays a clear tripartite pattern, with relatively rich occurrence of tree remains (mainly Fagus and Betula) in the lower and upper parts of the sequence and an intermediate section with very few tree macrofossils (Figure 2). Sediments deposited between c. 1450 and 1700 CE contain relatively large amounts of Fagus leaf fragments (large and small) and buds, as well as unidentified leaf fragments, but concentrations are generally lower after c. 1550 CE. Betula fruits were also recorded in the lower part of the sequence, but only continuously up to c. 1550 CE, and single macrofossils of Alnus (fruits) and Carpinus (leaf fragments and fruits) were found in sediments deposited before c. 1550 CE. No Fagus remains were identified in sediments dated to c. 1700-1900 CE, but some unidentified leaf fragments were found. Betula fruits are almost continuously present in low numbers within this intermediate section, and a few female catkin scales were also recorded. Fagus leaf fragments (large and small) and buds reappear at c. 1900 CE and are relatively abundant in the upper part of the sequence, with concentrations comparable with those before c. 1550 CE. The record of unidentified leaf fragments displays a similar pattern. Betula fruits are common in sediments deposited after c. 1900 CE, showing the highest concentrations recorded within the sequence, and female catkin scales occur in most samples. In addition, leaf fragments of Betula, Alnus, Carpinus and Quercus are present, but only in some samples and in low numbers. Alnus fruits and cones also occur at some levels, and one Carpinus fruit was found in the upper part of the sequence.
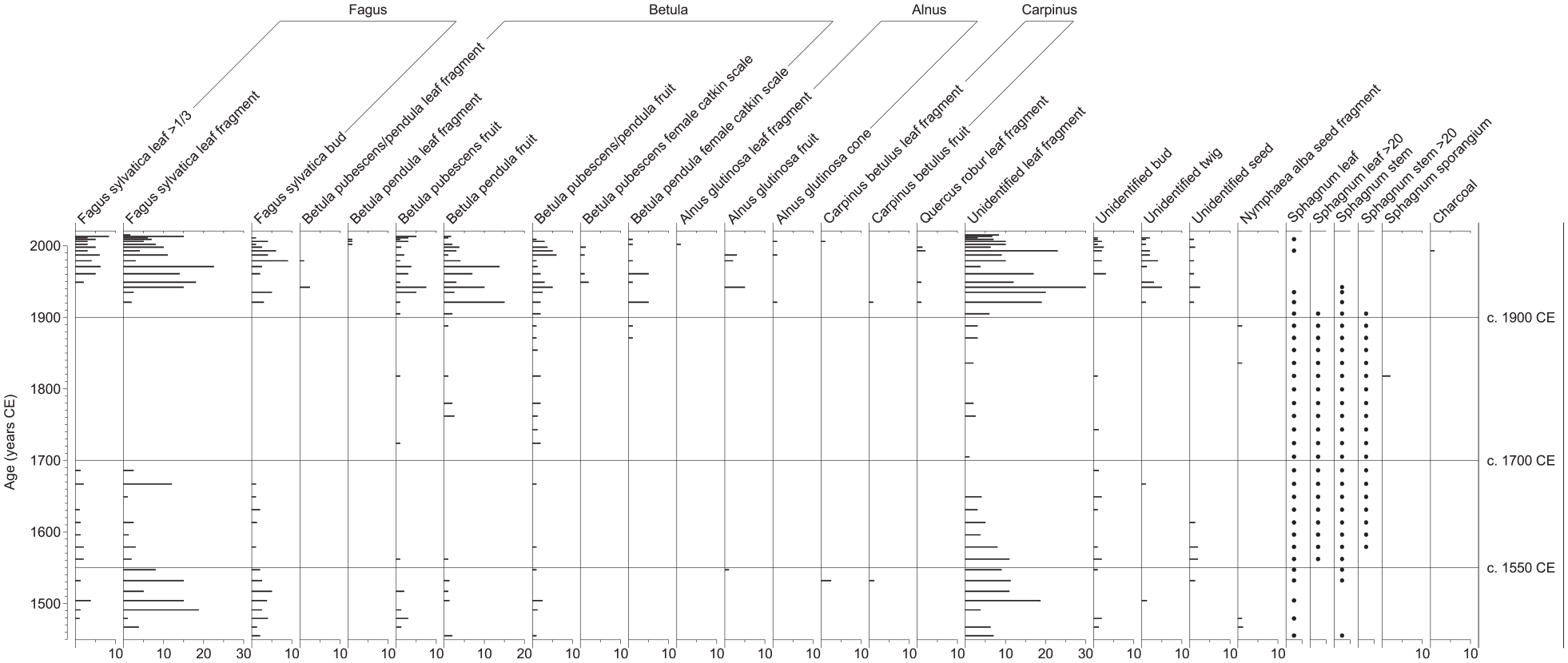
Plant macrofossil concentration diagram for the analysed sediment sequence from Lake Odensjön showing the number of finds per 50 cm3 of sediment, except for stems and leaves of Sphagnum mosses, which are indicated by dots as either present (≤20) or abundant (>20).
Single unidentified twigs were found in the oldest part of the sequence, none in the intermediate section deposited between c. 1700 and 1900 CE and relatively large amounts in the youngest part. This is consistent with the tree macrofossil records described above, and unidentified buds and seeds display similar patterns (Figure 2). Sediments deposited between c. 1550 and 1900 CE are rich in Sphagnum leaves and stems, but remains of peat mosses were also found in low numbers outside this interval. Single seed fragments of Nymphaea alba were recorded at a few levels in the lowermost and middle parts of the sequence.
Pollen data
The pollen record displays a tripartite pattern resembling that described for the macrofossil record, with a minimum in total tree pollen frequencies between c. 1700 and 1900 CE mainly reflecting low Fagus frequencies (Figure 3). The oldest part of the sequence (c. 1450–1700 CE) is dominated by Fagus pollen, with maximum values around 50–60% recorded before c. 1550 CE, but Betula and Alnus frequencies are also relatively high (c. 10–20%). Quercus and Pinus are continuously present with values reaching c. 5%, and Carpinus was recorded in almost all samples, while Tilia, Fraxinus and Ulmus show generally lower frequencies and more scattered occurrences. Single Picea pollen were found in the lowermost part of the sequence. The onset of a marked Fagus decrease occurs around 1550 CE. Alnus and Quercus are the only other tree taxa showing clear increases at the same time. Juniperus frequencies also start to increase, but a more marked increase to c. 20% begins later, after 1600 CE. Among the other shrubs recorded, only Corylus is continuously present between c. 1450 and 1700 CE, but values are consistently low (<5%). Calluna is present in almost all samples at similar frequencies. Empetrum is the only other dwarf-shrub taxon recorded but appears first around 1550 CE and shows even lower values. Poaceae pollen (<40 µm) were continuously recorded with c. 5–10% from the base of the sequence, but values are generally higher after c. 1550 CE, and Cerealia occurs with very low frequencies in most samples. Cyperaceae is almost continuously present but never reaches 5%. Filipendula, Plantago and Brassicaceae are the most frequently recorded herb pollen types between c. 1450 and 1700 CE, but their values are well below 5%. Polypodiaceae undiff. spores show relatively stable frequencies around 5%. Sphagnum spores are also almost continuously present, although values are generally lower but increase at c. 1550 CE. Pollen of several aquatic taxa were recorded at low frequencies in sediments deposited up to c. 1700 CE.
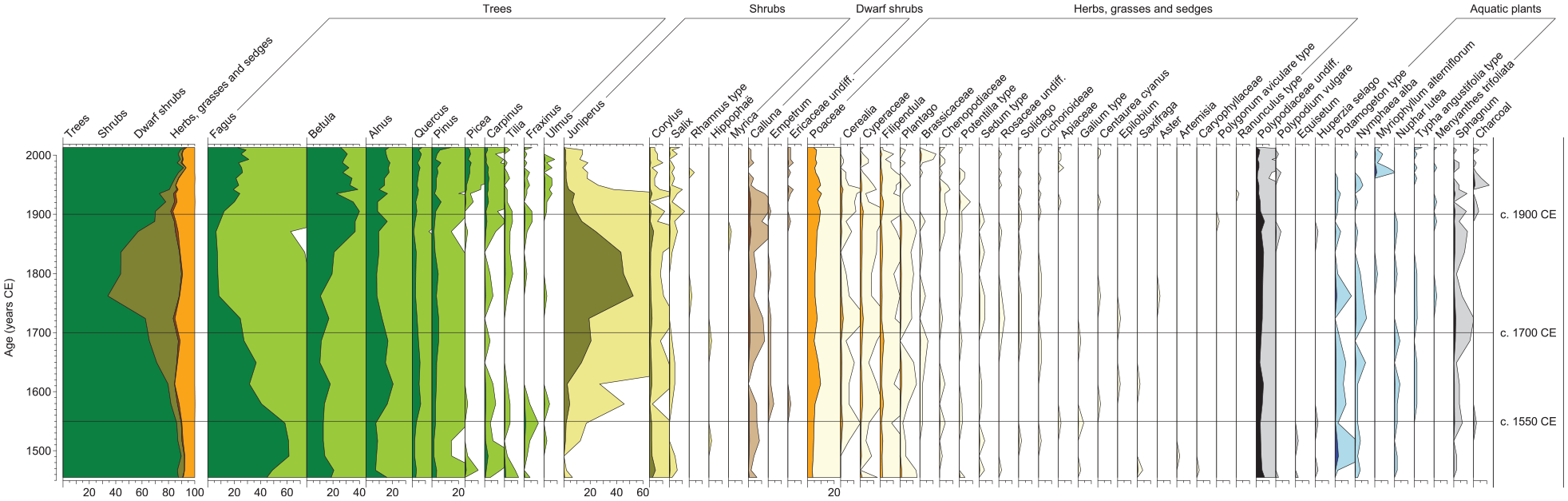
Pollen percentage diagram for the analysed sediment sequence from Lake Odensjön with silhouette curves in light colours representing 10 x exaggeration of the data to enhance visibility of changes in rare pollen types. Percentage calculations were based on the sum of terrestrial pollen. For taxa belonging to other groups (ferns, clubmosses, aquatic plants, and mosses), and for charcoal fragments, percentages were based on the combined sum of terrestrial pollen and the total count of the relevant group.
A second and major step in the decrease of Fagus pollen begins around 1700 CE, culminating in minimum frequencies of 5–10% recorded until c. 1900 CE (Figure 3). Alnus and Quercus decrease together with Fagus, while Betula frequencies initially remain between 10 and 20% but later start to gradually increase to c. 40%. Pinus continues to be present at similar frequencies as before c. 1700 CE. The occurrences of Carpinus, Fraxinus and Ulmus are more scattered than before, and values are generally lower, while there is a slight increase in Tilia. Only one Picea pollen was found. Juniperus displays a prominent peak in the period of c. 1700–1900 CE at maximum frequencies of 40–50%, and Corylus continues to be present at low frequencies. The same applies to Calluna, but values are slightly higher than before, while the occurrence of Empetrum is more scattered. Poaceae (<40 µm), Cerealia and Cyperaceae frequencies remain similar, and there are no clear differences in the herb pollen records compared with the period before c. 1700 CE. Frequencies of Polypodiaceae undiff. and Sphagnum spores remain similar after c. 1700 CE, as do the records of aquatic pollen taxa.
Fagus frequencies are relatively stable at c. 20–30% after c. 1900 CE (Figure 3). Following initial maximum values around 40%, Betula frequencies gradually decrease to c. 25%. Alnus and Quercus show slightly higher values than before, while Pinus frequencies remain at a similar level except for a recent increase to c. 15%. Carpinus was continuously recorded with slightly higher values after c. 1900 CE, and Fraxinus and Ulmus occur more regularly, while the presence of Tilia is more scattered. Picea is continuously present and shows frequencies around 5% after c. 1950 CE. At the same time, the Juniperus decrease initiated before c. 1900 CE reaches values below 5%. Corylus frequencies are generally lower after c. 1900 CE and even lower after c. 1950 CE. Calluna values also drop around 1950 CE, while no Empetrum pollen were found after c. 1900 CE. On the other hand, Ericaceae undiff. was recorded in most samples after c. 1900 CE but at very low frequencies. Poaceae (<40 µm) frequencies remain similar. Initially, the same applies to Cerealia, but very few pollen were recorded after c. 1950 CE. The occurrence of Cyperaceae is more scattered after c. 1900 CE, and frequencies are lower. The herb pollen records do not display any major differences, while Polypodiaceae undiff. and Sphagnum spores are less frequent. Aquatic pollen taxa continue to be present at low frequencies but change in relative importance.
Tree-ring data
The estimated germination period of the selected Fagus trees ranges from 1825–1835 to 1925–1935 CE (Table 1, Figure 4a). The limited number of trees in the dataset makes interpretation of establishment patterns speculative, but a potential phase of more extensive establishment is identified between c. 1860 and 1890 CE, and establishment appears to have followed a similar pattern in the two groups of sampled trees (Figure 4a). The data also reveal that the Fagus population has experienced notable growth variations, in particular a period of relatively strong growth during the 1980s and a period of restricted growth from 1995 to 1998 CE (Figure 4b).
Dendrochronological data for the ten Fagus trees (named 1-10) cored at Lake Odensjön: laboratory identification number, number of measured rings, the year of the youngest (outermost) and oldest (innermost) measured ring, the estimated number of missing rings toward the pith, the estimated germination period, the mean radial growth (mean TRW) in millimetres, and relevant remarks. Trees 1-5 belong to the group sampled on the western side of the lake, and trees 6-10 to the group sampled on the eastern side of the lake. From the ten analysed trees a 176-year tree-ring width (TRW) chronology was constructed spanning the period 1844-2019 CE (Figure 4), with an average tree series length of 114 years, and a mean series intercorrelation of 0.458.
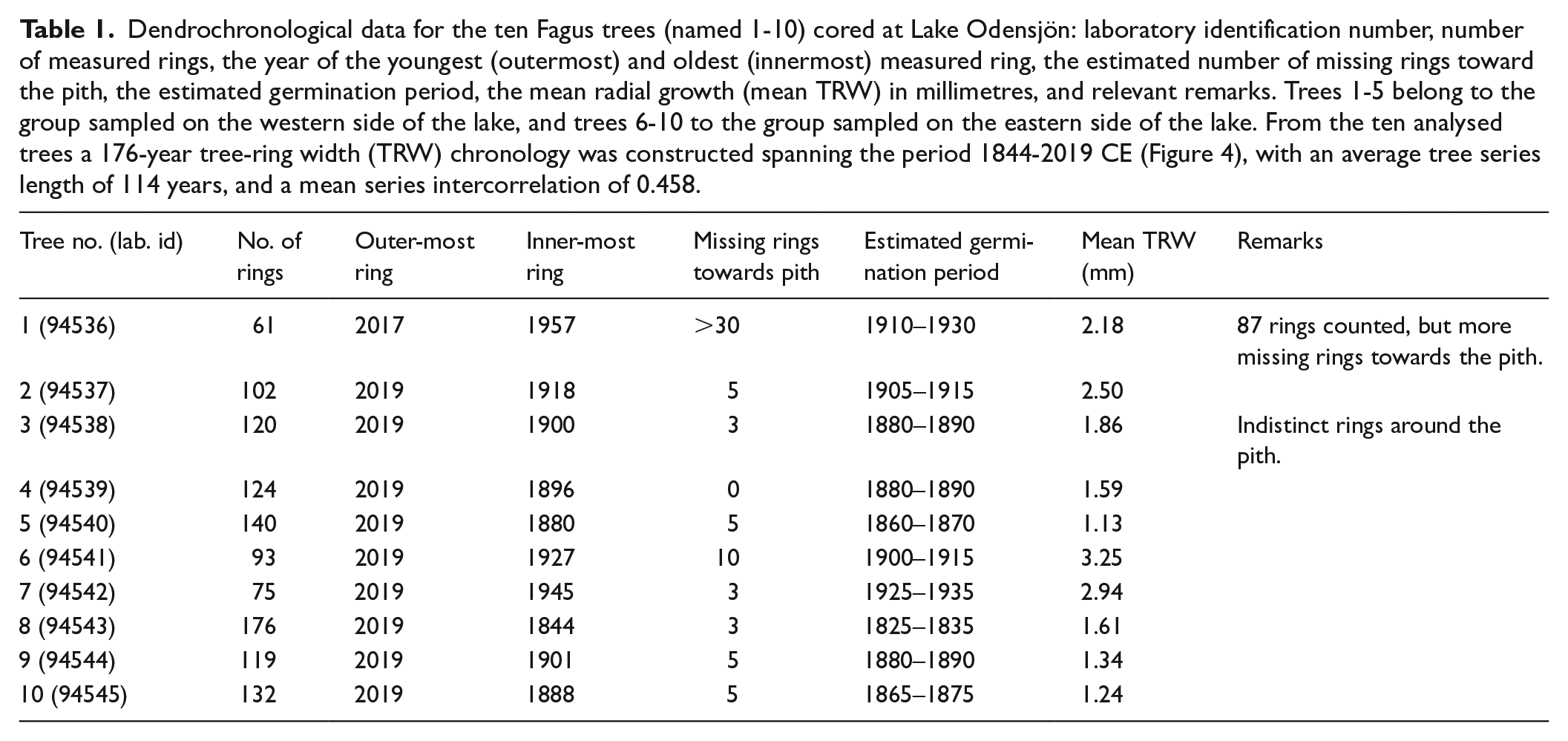
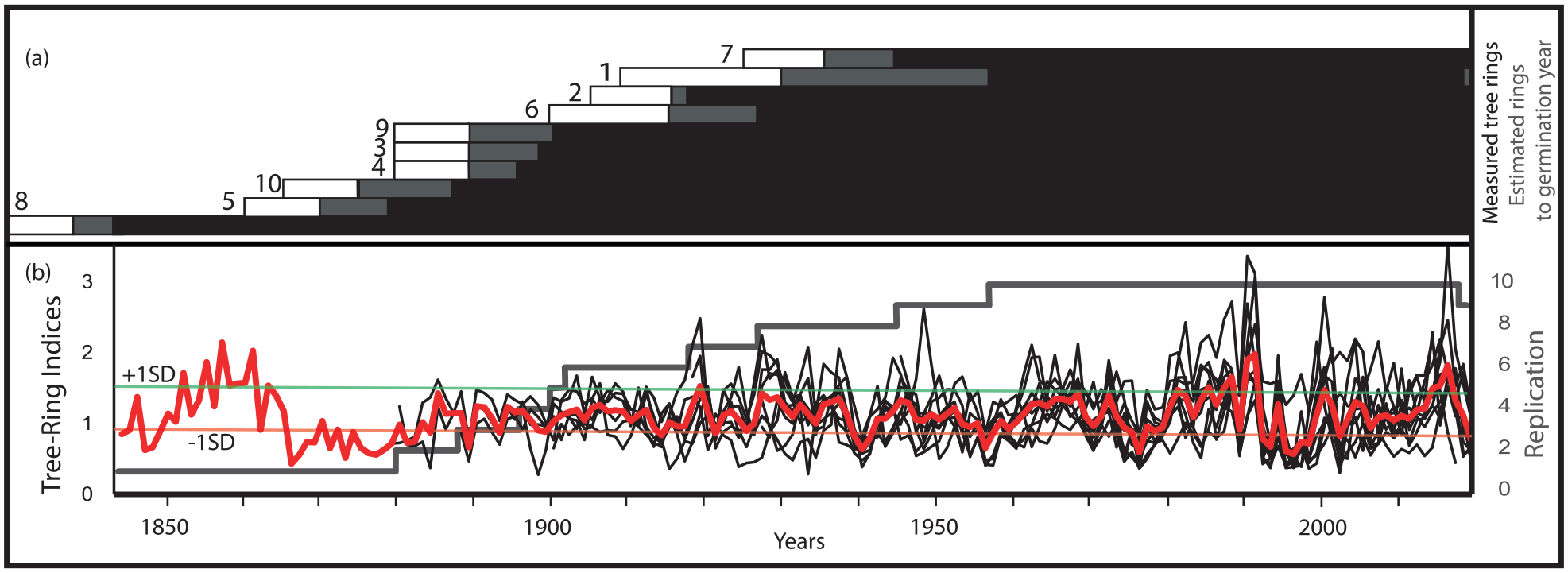
Results of the dendrochronological analysis of the ten Fagus trees cored at Lake Odensjön. (a) Overlapping tree sequences with the period covered by measured annual rings shown in black, estimated missing rings shown in grey, and the estimated germination period shown in white. The numbers (1–10) correspond to the tree numbers in Table 1, but here the samples are sorted by estimated germination period. (b) Tree-ring indices from individual trees (black) and the averaged tree-ring chronology (red). The grey curve shows the tree-replication record, and the thin red and green horizontal lines indicate the thresholds for ring-width values above and below one standard deviation (SD), respectively.
Discussion
Changes in forest composition and tree cover
The relatively rich occurrence of Fagus macrofossils indicates the presence of a substantial Fagus population on the slopes around Odensjön between c. 1450 and 1550 CE, and single macrofossil finds show that Alnus and Carpinus also were present close to the lake (Figure 2). Relatively high concentrations of Betula fruits demonstrate its local presence, but the lack of leaves suggests that Betula was mainly growing on the upper slopes rather than at the lake margin. Considering that Odensjön is very small and surrounded by steep slopes, the pollen record is interpreted as mainly reflecting local vegetation, especially at times of dense tree cover when local pollen production is higher and wind transport of pollen to the lake basin from neighbouring areas on the plateau above and beyond is likely to be less efficient (Faegri et al., 1989). Accordingly, high frequencies of Fagus pollen support the macrofossil evidence that Fagus was widespread on the slopes in the period of c. 1450–1550 CE, and the pollen records of Betula, Quercus, Carpinus, Tilia, Fraxinus and Ulmus indicate that these tree taxa also were present there (Figure 3). The Alnus record probably reflects trees growing at the lake margin and in the fen area around the outlet. However, the low Pinus pollen frequencies are interpreted as reflecting presence of pine forest regionally, because Pinus rarely occurs as a minor component in broad-leaved forests in southern Sweden. The high proportion of tree pollen (>80%) indicates a dense tree cover at the lake margin and on the surrounding slopes during the initial century of the record.
Decreased tree cover and a reduced Fagus population on the slopes around Odensjön after c. 1550 CE are indicated by lower concentrations of Fagus macrofossils and unidentified leaf fragments (Figure 2) and lower Fagus pollen frequencies (Figure 3). Fewer finds of Betula fruits indicate that also the local Betula population was reduced. However, Betula pollen frequencies remain at a similar level, suggesting that the decrease in Betula mainly occurred close to the lake. The increase in Quercus pollen may be interpreted as an expansion of Quercus as the forest became more open but could also reflect improved conditions for pollen production and dispersal in a more open landscape. Quercus, Carpinus, Tilia, Fraxinus and Ulmus were probably still present on the slopes surrounding the lake, but the pollen records suggest that their populations were reduced. Elevated Juniperus and Poaceae (<40 µm) pollen frequencies after c. 1550 CE may partly reflect increased presence of these taxa on the slopes in response to improved light conditions associated with a reduced tree cover. However, they may also reflect stronger representation, and possibly an expansion, of heathland vegetation on the plateau above, because a more open vegetation on and immediately above the slopes would facilitate wind transport of pollen from vegetation in the surrounding, higher areas. Although heathland expansion is not clearly reflected by the Calluna record, slightly higher Calluna frequencies were recorded following the subsequent marked Juniperus increase around 1600 CE. However, heathland expansion on the plateau after c. 1550 CE is supported by the appearance of Empetrum in the pollen record. The increase in Alnus pollen frequencies at c. 1550 CE could partly be a proportionality effect of lower input of Fagus pollen from the slopes but is likely to also reflect an Alnus expansion in the fen area around the lake outlet, which may explain the lower abundance of Betula macrofossils. Expansion of Alnus could reflect wetter conditions along the lake margin caused by increased surface run-off on the slopes as tree cover decreased. Wetter condition in the outlet area may also be inferred from the observed increases in Sphagnum macrofossils and spores, but increased shading from an associated Alnus expansion would be unfavourable to peat mosses growing there. Therefore, the Sphagnum increase is instead interpreted as reflecting an expansion of aquatic Sphagnum species in response to improved light conditions in the near-shore environment as shading from trees on the surrounding slopes decreased. This scenario, which is supported by diatom data from the same sediment sequence (Silvester et al., 2025) also better explains the rich occurrence of well-preserved Sphagnum macrofossils in the sediments.
An interval with only low concentrations of unidentified leaf fragments in a few samples indicates that the decrease in tree cover around Odensjön starting at c. 1550 CE culminated in very open conditions between c. 1700 and 1900 CE (Figure 2). This is supported by a clear minimum in the proportion of tree pollen (Figure 3). Very low Fagus pollen frequencies indicate that the presence of Fagus on the surrounding slopes was limited during this time, and more fragmentary pollen records and lower frequencies of Quercus, Carpinus, Tilia, Fraxinus and Ulmus suggest that the populations of most slope forest species decreased. Lower pollen frequencies indicate that the Alnus population in the fen area and at the lake margin also was reduced. However, initially unchanged and subsequently increased pollen frequencies indicate that the local Betula population remained intact and later even expanded. Exceptionally high Juniperus pollen frequencies provide clear evidence of very open conditions. Juniperus therefore probably continued to expand on the slopes around the lake as the shading from trees decreased, but the maximum frequencies recorded within the period of c. 1700–1900 CE are difficult to explain without invoking a more large-scale Juniperus expansion on the plateau above. Moreover, if these unusually high Juniperus pollen frequencies only represent a slope population, it would be expected that some macroscopic Juniperus remains, for example needles, were recorded in the sediments, which is not the case. However, among the other pollen taxa that potentially represent heathland vegetation, that is Calluna, Empetrum and Poaceae (<40 µm), only Calluna shows a minor increase after c. 1700 CE, but this may be explained by a proportionally larger expansion of Juniperus. Interestingly, a major decrease in Juniperus pollen frequencies occurs at c. 1830 CE, coinciding with the previously noted Betula pollen increase. This is interpreted as the beginning of a phase of tree expansion that restricted the distribution of Juniperus and initially mainly involved Betula. Because Betula is a common component of heathland vegetation, Juniperus may have been affected by increased shading from Betula both on the slopes and on the plateau. As indicated by the estimated establishment age of the oldest Fagus tree analysed (Figure 4a), Fagus may also have started to expand on the slopes around the lake at this time. The continued rich occurrences of Sphagnum macrofossils and spores support generally open conditions around the lake between c. 1700 and 1900 CE.
An increase in Betula macrofossils is registered at c. 1900 CE, indicating that Betula continued to expand on the slopes surrounding the lake (Figure 2). Fagus macrofossils reappear in the sediments around the same time and soon become as abundant as between c. 1450 CE and 1550 CE, which is consistent with the dendrochronological data, indicating widespread colonisation by Fagus during the late 19th century (Figure 4a). Together with single macrofossils of Carpinus, Quercus and Alnus, this shows that there was a general expansion of trees on the slopes, at the lake margin and in the fen area. However, the tree pollen records suggest that Fagus was less dominant on the slopes relative to the situation before c. 1550 CE (Figure 3). Fagus frequencies are substantially lower, while especially Betula, but also Quercus and Carpinus, frequencies are higher. The continuously high Betula frequencies after c. 1900 CE indicate that Betula managed to maintain parts of its recently expanded slope population, probably on and next to the lower talus areas where competition from Fagus was limited. However, the generally high Betula frequencies suggest that Betula also remained an important component of the vegetation on the plateau. The Picea pollen record is interpreted as reflecting the appearance of Picea plantations in nearby areas of the Söderåsen horst, and planting within former heathland is indicated by decreased pollen frequencies of Juniperus, Calluna and Empetrum. Introduction of new methods of forest management may also help explain the generally high Betula frequencies after c. 1900 CE, in particular clear-cutting that provides suitable open habitats. The almost continuous presence of Ericaceae pollen is likely to mainly reflect Vaccinium growing as understorey, primarily in Picea forest but possibly also in relatively open areas of Fagus-dominated forest on the slopes around the lake. The Sphagnum macrofossil record is sparse after c. 1900 CE, indicating deteriorated light conditions in the near-shore environment as an effect of tree expansion on the slopes.
Historical processes shaping the present Fagus-dominated forest
By 1450 CE, when deposition of the analysed sediment sequence from Odensjön began, Fagus had been present locally in southernmost Sweden for several thousand years (Björkman, 1999), but its early history on the Söderåsen horst is unclear. Based on pollen data, Göransson (1999) suggested that Fagus was established about 2000 years ago at a small peatland site on the horst plateau c. 2 km NW of Odensjön (Figure 1), but there are no radiocarbon dates to support this interpretation. Pollen records from two other small peatlands located on the plateau 3–4 km NW of Odensjön (Figure 1) indicate that Fagus-dominated forest developed in that area at c. 800–1000 CE (Bergman, 2000). During the preceding 2–3 millennia, the area is interpreted as having been characterised by mixed broad-leaved forest dominated by Quercus. At one of the sites, a short-lived event with high frequencies of Poaceae precedes the development of Fagus-dominated forest. This is interpreted by Bergman (2000) to reflect local establishment of Fagus facilitated by a change to more open conditions caused by an onset of intensive grazing and a subsequent expansion of Fagus when grazing pressure decreased. At the two other plateau sites, development of Fagus-dominated forest also coincides with a change to more open conditions, but these records do not show any evidence of a similar preceding event of strong grazing pressure (Bergman, 2000; Göransson, 1999).
Little is known from historical sources about vegetation and land use on the Söderåsen horst at the time when the oldest analysed sediments from Odensjön were deposited (c. 1450 CE), but this agriculturally marginal area was most likely dominated by forest. However, agricultural land must have been present around villages and farms, and some forest areas are known to have been used for grazing (Persson, 1971). This general picture is consistent with the macrofossil and pollen records from Odensjön, reflecting a relatively closed Fagus-dominated forest on the slopes surrounding the lake between c. 1450 and 1550 CE (Figures 2 and 3). The first map with information on land use in Scania is a military map published by Buhrman in 1684 CE, which shows the Söderåsen horst as almost completely covered by Fagus forest, while surrounding lowland areas largely lack forest (Riksarkivet, 2025a). The large extent of Fagus forest at this stage may partly be explained by legislation that for a long time had prevented farmers from felling mature Fagus and Quercus trees on their land, as these belonged to the government (Persson, 1971).
Historical documents show that the forests on the Söderåsen horst were increasingly used for grazing during the 17th and 18th centuries (Campbell, 1928; Håkansson, 1948; Persson, 1971). This increasing grazing pressure, mainly from cattle and horses, made forest rejuvenation more difficult. However, because its leaves are unattractive to animals, the impact on Fagus was probably not as strong as on most other trees (Emmanuelsson et al., 2002). In addition, the forests were used as a source of firewood and other resources by local farmers, and logging was carried out by the government (Persson, 1971). Clearing of forest followed by burning to improve conditions for grazing probably also occurred (Lindquist, 1931). Because of the abundance of nuts on the forest floor, mainly beechnuts but also acorns, large areas were also used for pig grazing. In contrast to other types of grazing, this is considered to promote Fagus rejuvenation as it results in some nuts being buried in the soil (Emmanuelsson et al., 2002; Sjöbeck, 1927). Together, these land-use practices are likely to have promoted the dominance of Fagus in the forests of the Söderåsen horst as shown by the 1684 AD map (Riksarkivet, 2025a) and as reflected by pollen evidence from the plateau area dated to around the same time (Bergman, 2000). The more well-dated and highly-resolved macrofossil and pollen records from Odensjön provide a similar picture, with strong dominance of Fagus in the forest, but they also indicate that intensified grazing was already causing opening of the forest on the slopes around the lake, and probably on similar talus slopes along the margins of the horst and its interior fissure valleys, by c. 1550 CE (Figures 2 and 3). Moreover, our new records suggest that the forest on the plateau above Odensjön was beginning to be replaced by heathland with Juniperus, Calluna and Empetrum around the same time. This is consistent with substantial increases in Juniperus and Calluna frequencies in the uppermost parts of the pollen records from the site c. 2 km to the NW studied by Göransson (1999; Figure 1), although the lack of dating hampers linking of these events. The absence of Juniperus and Empetrum pollen, as well as the lower Calluna pollen frequencies, in the records presented by Bergman (2000) indicate that the grazing pressure during the 17th and 18th centuries was weaker in the more central part of the horst plateau.
Despite the lack of Fagus macrofossils at c. 1700–1900 CE (Figure 2), the pollen record suggests that a Fagus population, although significantly reduced, continued to exist on the slopes around Odensjön during that time. The dendrochronological data also show that Fagus was present at least from c. 1830 CE (Figure 4a). However, no symbols representing trees are evident on the lake slopes on a military map of Scania published in 1812–1820 CE (Riksarkivet, 2025b), but the scale of this map together with the shading used to illustrate topography makes identification of any such symbols difficult. However, the map clearly shows that trees were essentially absent from the plateau above Odensjön and within an area of about 2 km to the W and 1 km to the NW. Further north, the plateau is depicted as being tree-covered to a varying degree, consistent with the pollen records from that area (Bergman, 2000; Göransson, 1999). This pattern of forest cover is supported by a cadastral map from 1833 CE (Lantmäteriet, 2025a) with tree symbols restricted to the slopes around the lake and adjacent horst margin slopes. However, in the map description the slopes are included in a larger area of ‘open grazing land’ reaching c. 1 km to the W on the plateau. A military map from 1865 CE (Lantmäteriet, 2025b) also shows forested slopes around the lake and along neighbouring parts of the horst margin, while the plateau area W and NW of the lake is shown as predominantly open. Further north, where the sites studied by Göransson (1999) and Bergman (2000) are located, the degree of tree cover is generally lower compared with the map from 1812 to 1820 CE, suggesting that maximum openness was reached later in that part of the plateau. Based on the description accompanying a cadastral map from 1866 CE (Lantmäteriet, 2025c), the slopes around Odensjön were used for grazing at least as late as in the 1860s. In 1793 CE, the legislation restricting farmers’ use of mature Fagus trees on their land was repealed (and in 1830 CE for Quercus). This may have contributed to the very open conditions during the first half of the 19th century, but probably there were few trees suitable for logging left by then.
Based on the pollen record, landscape openness was at its maximum in the Odensjön area during the late 18th and early 19th centuries (Figure 3), which is supported by the evidence of intensified grazing on the Söderåsen horst revealed by historical documents (Campbell, 1928; Håkansson, 1948; Persson, 1971). This increasing pressure on the landscape is likely to be linked to the general increase in rural population occurring within this period in large parts of Sweden. In Röstånga Parish, where Odensjön is situated, the population almost tripled from 1699 to 1830 CE (Figure 5). This increase in local population is consistent with general increases in birth numbers, child survival and life expectancy in Sweden during this time and the break in this development may be explained by urbanisation and emigration. Interestingly, the demographic change around 1830 CE coincides with the first pollen evidence of forest regrowth on slopes surrounding the lake and on the plateau above, that is, the decrease in Juniperus and increase in Betula (Figure 3). The oldest Fagus tree sampled on the slopes also germinated at this time (Table 1, Figure 4a). In addition to changes in population development, land-use changes on the Söderåsen horst during the first half of the 19th century may be explained by effects of the land reforms introduced in Sweden during that time. Of these, ‘Laga skifte’ probably had the strongest influence on the Söderåsen landscape, and one of its consequences was that grazing became more restricted to areas in close proximity to the farms and less common in the surrounding forests (Persson, 1971). The area around Odensjön was subjected to ‘Laga skifte’ in 1833 CE, and the map from that year (Lantmäteriet, 2025a) was produced in that context. The subsequent forest regrowth on the slopes around the lake and on the plateau above indicated by the macrofossil and pollen records, and supported by the historical maps, may therefore partly be a result of this reform.
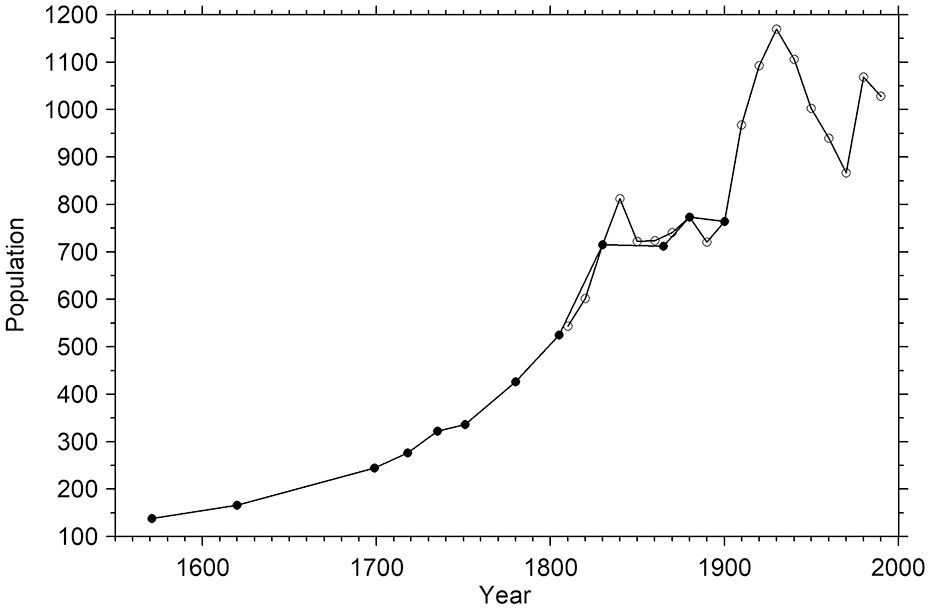
Population development in Röstånga Parish 1571-1990 CE. The data series with filled circles (1571-1900 CE) is based on the subdivision ‘socken’ (Palm, 2000), and the data series with open circles (1810-1990 CE) on the subdivision ‘församling’ (Demografiska databasen, CEDAR, Umeå universitet, 2022). This reflects a change in the administrative basis of population statistics in Sweden. In most cases the areas covered by a ‘socken’ and a ‘församling’ are identical.
A drawing of a view across the lake from the south-western edge of the plateau, published in a book with plates illustrating picturesque views from Sweden, Norway and Denmark (Bonnier, 1868), provides a fascinating insight into the surrounding landscape in the mid-19th century (Figure 6). Except for an area with exposed rocks in the uppermost part and some of the lower areas covered by stones and boulders, the slopes are illustrated as having a relatively dense tree cover. Although identification is difficult, Betula appears to be dominant, but some Fagus may be present in the highest parts. No Juniperus is visible on the slopes, and the small area of the plateau from where the artist overlooks the lake has a more open vegetation compared with the slopes, and with the same area today (Figure 6). Clearly, this illustration is consistent with a relatively open slope forest dominated by Betula, especially in the lower parts, during the mid-19th century, as inferred from the macrofossil and pollen records. Interestingly, it also supports our interpretation that the very high Juniperus pollen frequencies recorded do not primarily represent vegetation on the slopes, but most likely heathland on the neighbouring plateau. A very small area with Calluna-dominated heath that may be a remnant of this heathland is found today next to the viewpoint from where the drawing was made.

Comparison of views across Lake Odensjön from the south-western plateau edge towards the outlet area in the north. (a) Drawing made by an unknown artist in the mid-19th century (Bonnier, 1868). Note the absence of trees on the eastern talus slope and the rocky area above to the right, and the open vegetation at the viewpoint from where the drawing was made. Public Domain. (b) Outlook from approximately the same viewpoint in October 2024. Dense tree cover at and below the plateau edge makes it difficult to see the lake. Note the bedrock outcrop also visible in the drawing in (a). (c) Outlook from the same viewpoint as in (b) but when standing immediately at the plateau edge. Although branches of trees on the slope below prevent a good view of the lake and its surroundings, the rocky area above the eastern talus slope seen in the drawing in (a) is not visible in 2024 because of the presence of Fagus on the upper slope.
The slopes around Odensjön and along the neighbouring areas of the horst margin are illustrated as being covered by trees on a regional economic map published in 1910–1915 CE (Lantmäteriet, 2025d), which indicates that the forest in these areas by then had recovered after grazing was abandoned, consistent with the evidence of Fagus expansion on the slopes surrounding the lake based on our macrofossil and pollen records (Figures 2 and 3). Moreover, the horst plateau area W and NW of the lake, including the northern part where the sites studied by Göransson (1999) and Bergman (2000) are located, is covered by tree symbols, although their density is lower in some areas. This suggests that grazing had decreased substantially or had been abandoned also on the plateau. In addition, the presence of a few small areas with symbols representing conifer trees is the first evidence of planting of Picea on the plateau, and one of these areas is located only 100–300 m NW of the lake. This fits very well with the beginning of the Picea pollen record dated to c. 1910 CE (Figure 3). An aerial photograph from 1947 CE (Lund University, 2025) reveals several Picea plantations on the horst plateau, for example in the same area as indicated on the 1910–1915 CE map (Lantmäteriet, 2025d) and another one only 100 m SW of the lake. It also shows that most of the plateau was covered by dense forest at that time, but tree cover was generally lower W of the lake. Interestingly, an open area with only scattered trees or bushes occurs 200–600 m WNW of the lake, probably representing a small remnant of the previously widespread heathland. This area appears to still be relatively open on an aerial photograph from c. 1960 CE (Lantmäteriet, 2025e), which also shows that Picea plantations covered larger areas of the plateau at that time. However, the previously open area is almost completely covered by forest on an aerial photograph from c. 1975 CE (Lantmäteriet, 2025f).
Comparison with other palaeoecological studies of the development of Fagus-dominated forest in southern Sweden
In pollen records from sites within Fagus-dominated forests in southern Sweden, situated outside the natural range of Picea, evidence of local Fagus establishment is dated to between c. 2000 BCE and 1200 CE (Björkman, 1997, 2001; Hannon et al., 2018, 2022; Hultberg et al., 2010; Lindbladh et al., 2008). Despite these large differences in timing, the establishment is associated with increasing Poaceae pollen frequencies at all sites, and often also with increases in Juniperus or Calluna, or both. Local Fagus establishment is therefore inferred to be dependent on opening of the forest related to human activity, mainly introduction of grazing or increasing grazing pressure. These studies also show that the timing of local Fagus expansion varies from immediately following establishment to c. 2500 years after establishment. Moreover, Fagus expansion is inferred to have occurred in more than one step at several sites. Like establishment, expansion of Fagus is typically found to be associated with increases in pollen taxa indicative of open conditions, for example Poaceae, Juniperus and Calluna. Consequently, also Fagus expansion is attributed to increasing grazing pressure.
Maximum Fagus pollen frequencies are attained at the very top of some of the records, indicating recent, and possibly on-going, expansion of Fagus (Hannon et al., 2018, 2022; Hultberg et al., 2010), while other records display lower Fagus values in the uppermost part. However, interpretation of recent Fagus population dynamics is complicated in some studies by the establishment of taxa with high pollen productivity and efficient pollen dispersal, usually Betula or Alnus, on the peatland surface, resulting in a proportional decrease in Fagus pollen frequency that may be mistaken for a recent reduction of the local Fagus population (Björkman, 2001; Lindbladh et al., 2008). This problem is pointed out by Bergman (2000) when interpreting the two pollen records from the Söderåsen horst plateau discussed above, and it probably also applies to the two records presented by Göransson (1999).
Interestingly, several of the pollen records from the region show increasing frequencies of Poaceae, Juniperus and Calluna during the 17th–19th centuries (Björkman, 1997, 2001; Hultberg et al., 2010; Lindbladh et al., 2008), indicating an opening of vegetation resembling that inferred from the macrofossil and pollen records from Odensjön. However, a subsequent expansion of Fagus is not apparent from some of the records, which may be explained by the proportionality effect related to recent establishment of other tree taxa on the studied peatlands. Comparison with the pollen record from Kullaberg presented by Björkman (2001) is particularly relevant, because this site is located on another horst ridge in NW Scania. Relative to the other records from the region, Juniperus and Calluna frequencies are relatively high within the period of c. 1650–1800 CE, reaching around 15% and 25%, respectively. This is interpreted by Björkman (2001) as an event of local deforestation followed by heath expansion, which is supported by historical documents and maps showing that the entire plateau area on the Kullaberg horst was covered by intensively grazed heathland in the mid-18th century. As in the pollen record from Odensjön, Fagus frequencies are very low during the period with high Juniperus values. Fagus frequencies recover slightly as Juniperus values start to decrease, but they never reach the levels attained before c. 1650 CE. However, this is explained by the proportionality effect related to establishment of Betula on the peatland surface (Björkman, 2001). Although the present forest surrounding the Kullaberg site is less dominated by Fagus compared with the slope forest around Odensjön, and despite the much less well-constrained chronology of the Kullaberg record, the similarities in forest development are striking. This is interpreted as reflecting a common land-use history during recent centuries in these two, closely spaced (c. 50 km) areas with comparable topographic, geological and climatic settings. Given the strong similarities observed, the inferred history of Fagus-dominated forest at Odensjön is proposed to be representative not only of Söderåsen National Park and large parts of the Söderåsen horst, but also of other agriculturally marginal areas in similar settings within Scania, and probably also within other parts of southernmost Sweden and neighbouring countries.
Implications for forest management
As inferred from the macrofossil and pollen records from Odensjön, and supported by historical maps, documents and aerial photographs, Fagus continued to be present, although with a reduced population, on the slopes surrounding the lake during the period of maximum landscape openness in the late 18th and early 19th centuries. This is probably explained by the fact that these slopes are generally very steep and largely covered by sharp-edged stones and boulders, making them difficult to access for grazing animals. As indicated by historical maps, open forest with Fagus also occurred on neighbouring slope areas along the horst margin and is likely to also have done so on similar slopes along other parts of the horst margin and the deep fissure valleys characteristic of the Söderåsen horst landscape. Although scattered Fagus individuals suffering from heavy grazing probably occurred within the plateau heathlands, the continuous presence of relatively dense Fagus populations in these slope areas is likely to have facilitated the rapid development of Fagus-dominated forest on the Söderåsen horst in the late 19th century. Forest survival in slope areas during the period of maximum grazing pressure probably also provided continuous availability of suitable habitats for some of the rare species belonging to different organism groups found within the park today, in particular mosses, lichens and fungi.
As Fagus rapidly expanded in previously vast heathlands, large parts of the Söderåsen horst became covered by Fagus-dominated forest with a homogenous age structure. Where later forest management has been limited, the effect of this process is visible today as the characteristic, almost monospecific and mature Fagus stands with limited shrub-, field- and ground-layer vegetation that attract many visitors to Söderåsen National Park. However, trees within these stands are now reaching an age when they become susceptible to attack by fungi and insect herbivores, and to wind-felling, which creates open patches. Although this probably facilitates Fagus regeneration, the resulting gradual replacement of trees over the coming decades is likely to result in Fagus stands with a more heterogenous age structure. Maintaining Fagus-dominated forest similar to that found in many parts of Söderåsen National Park today may therefore require successive opening of large areas where new Fagus stands can develop through regeneration within a few decades. This could be achieved through step-wise thinning, possibly complemented by planting. Such management may also be appropriate in other protected areas within southernmost Sweden and neighbouring countries where there is an ambition to maintain large Fagus-dominated forests with a homogeneous age structure. However, this type of forest management is not consistent with the aim to allow broadleaved forest to develop naturally within the park (Naturvårdsverket, 2001), which highlights some key questions within nature conservation: what is natural, which ecosystems do we want to preserve, and why, and what do we need to do to preserve them? Clearly, the type of management that we outline above may be questioned, because it aims at preserving an ecosystem that has developed in response to historical land use and therefore can be described as anthropogenic. However, it may also be questioned whether forest development according to the current management plan will result in an ecosystem that can be characterised as natural.
Footnotes
Acknowledgements
Yevhenii Rohozin is thanked for help during collection of the freeze core from Lake Odensjön in 2017. We are also grateful for guidance from Ethan Silvester during subsampling of the core in 2020. The Söderåsen National Park administration was helpful in the planning of the fieldwork. Jonas Ising is thanked for preparation of the map in ![]() .
.
Author contributions
Data availability
The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: The palaeoecological and dendrochronological analyses forming the basis for this study were performed within a MSc project at the Department of Geology, Lund University, carried out by Maja Damber. The department financially supported the laboratory work and part of the fieldwork.
Ethical approval statement
Not applicable.