
Abstract
The Holocene in Australia is characterised as a period of dramatic social and economic reorganisation in Aboriginal culture across the continent, with some of the most evident changes visible in the archaeological record of subsistence patterns. The zooarchaeology of key sites suggests that during the Mid- to Late Holocene, hunters switched emphasis away from high-yield large macropods to a diversity of smaller-bodied animals as a response to changing environmental conditions, demographic demands and/or available technologies. One such key site is Graman B4 (GB4), where a transition from kangaroo to possum occurred around 3000 years ago. However, the low resolution of legacy data from sites such as GB4, and contrary results from more recent excavations elsewhere, question the integrity of this broader trend. Here, we revisit the issue of subsistence change in eastern Australia through an original analysis of the archaeological fauna from the adjacent site Graman B1 (GB1), which has an older sequence extending into the early Middle Holocene. We find a persistent pattern of macropod-majority meat contributions throughout the entire ~6500 year sequence with varying degrees of reliance on smaller animals. Only within the last 2000 years are there signs of emergent specialisation in possum acquisition, though this is not at the expense of macropod hunting. We also detect a sharp decline in the use of GB1 coincident with the first occupation of GB4. Our results highlight the importance of high-resolution taxonomic identification and close consideration of animal body mass, as well as data from more than one site within an archaeological complex, to the construction of representative chronologies in Australian Holocene subsistence change.
Introduction
The Holocene in Australia has been characterised as a period of dramatic environmental and cultural developments, the impacts of which are reflected in its archaeological record. Widespread changes during this period include major restructuring of technological organisation, residential mobility, land-use patterns and subsistence habits (Hiscock, 1994, 2008). With respect to the last of these, changes in the representation of different taxonomic groups are the theme of most analyses of Holocene archaeological faunal assemblages from Australia. Zooarchaeological evidence from several sites spanning most of the continent suggests a trend of broadening taxonomic diversity in the game used by Aboriginal foragers. This trend begins in the Middle Holocene and is well-developed in the Late Holocene (Balme and O'Connor, 2016; Koungoulos, 2017).
The trend has been characterised as a decrease in the representation of large macropods (kangaroos and wallabies over 15 kg) and an increase in medium (5–15 kg) and small (0–5 kg) animals (Balme and O'Connor, 2016; Dortch, 2004; Fillios et al., 2010, 2012). The specific taxa which increased in importance vary according to the geographic setting of each site, but their ecologies tend to reflect greater, specialised exploitation of environments other than the strictly terrestrial settings where large macropods are found (Hiscock and Sterelny, 2023). These include, for instance: arboreal and hollow-dwelling marsupials from forests (McBryde, 1977); freshwater fish, molluscs and crustacea from rivers, lakes and wetlands (Allen et al., 2023); and marine fish, molluscs, crustacea, seabirds and seals of the intertidal zone, open ocean and offshore islands (Koungoulos et al., 2024). Graman – specifically, Graman Rockshelter B4 (henceforth GB4), is frequently referred to in discussions of Holocene faunal change.
GB4 is the best-known of a complex of rockshelter sites within the Graman district of the New England region in north-eastern New South Wales (Figure 1), originally excavated by McBryde (1968). These excavations produced a large and well-preserved faunal assemblage, which is one of very few known from inland settings in eastern Australia that clearly predate ~3000BP. Chronological change occurring in the taxonomic composition of GB4’s archaeological fauna generated a lively debate over their significance in the late 20th century (David, 1991; Morwood and Trezise, 1989; Sutton, 1990). Subsequently, the site has been widely cited as an example of major changes in Australian Late Holocene subsistence in multiple models attempting to explain this trend (Balme and O'Connor, 2016; Hiscock, 1994; Hiscock and Sterelny, 2023; Koungoulos and Fillios, 2020; Morwood and Trezise, 1989).
Location of the Graman archaeological site complex as defined by McBryde (1968) within Australia (left) and marked by star in inset area of north-eastern New South Wales (right).
There are several problems affecting the use of legacy faunal data from sites such as GB4 in broader models of Holocene change. Mostly published several decades ago, faunal data were usually included as minor components of larger site reports because the authors’ main interests usually lay elsewhere (site chronology and lithic technology). As such, the faunal reporting typically features little to no substantiation of identification and/or quantification methods (see Mein and Manne, 2021), and the actual abundance metrics are either unavailable or very lacking in detail. These low-resolution data are subsequently repackaged and interpreted by others without access to the original raw data or assemblages. In addition, the timing of major inflexions in dietary composition at a given site is often also poorly substantiated, and can vary considerably depending on the way that data are framed. This is exacerbated by the low resolution of these sites’ legacy radiocarbon chronologies, in turn sometimes reliant on potentially questionable results from before widespread radiocarbon dating (e.g. from the Gakushuin lab cf. Blakeslee, 1994).
Unsurprisingly, more recent zooarchaeological analyses by faunal specialists using modern methods note numerous exceptions to the supposed trend described above. In some cases these have revealed trends in the opposite direction, for example in northwestern Australia at the Kimberley sites Riwi (Balme et al., 2018) and Carpenters Gap 1 (Maloney et al., 2018). In others such as at Ngaut Ngaut and Tungawa 2 and 6 on the Lower Murray in Australia’s south, Late Holocene increases in some small-bodied taxa do occur, but are concomitant with declines in others (Allen et al., 2023). Elsewhere, as in southwestern Australia, there seems to have been little change over the Holocene (Dortch et al., 2012) and furthermore taphonomic assessments have revealed that some ‘archaeological’ assemblages are not the result of human behaviour (e.g. Asmussen, 2005). These results suggest a degree of regionalism applying to the direction and nature of Holocene subsistence change in Australia, and a need to scrutinise the underpinnings of claims for widespread Holocene subsistence changes in Australia through revisitation and novel analysis of key sites.
Another site in the Graman complex, Rockshelter B1 (henceforth GB1), features a deeper and older Holocene sequence than GB4. It also preserves a large faunal assemblage which has remained unstudied to date (Boot, 1990; McBryde, 1968). GB1 offers an excellent opportunity to revisit the issue of subsistence change in eastern Australia during the Holocene, particularly for the Graman area. In this work, we present an analysis of the archaeological fauna from GB1 supplemented by new radiocarbon dates, and a revised chronology for the site.
The Archaeology of Graman
The Graman sites are located in the valley of Ottleys Creek (a tributary of the Macintyre River) which lies on the western edge of Inverell Shire in north-eastern New South Wales. The sites sit on the western slopes of the Great Dividing Range, approximately 450–600 m above present sea level (Figure 1). The valley winds through basaltic high hill country, and where the river has eroded basalt, low cliffs of sandstone escarpment have been exposed along its banks. Within three sub-areas of the property along these escarpments, McBryde (1968) recorded 13 sites, including 8 rockshelters, 5 of which had evidence of occupation in their floor deposits. She chose two of these, Sites 1 (GB1) and 4 (GB4), for archaeological excavations, conducted between 1965 and 1967.
Excavations revealed well-preserved flaked lithic, ground stone, bone, shell and plant assemblages at both sites which were discussed in a series of papers (McBryde, 1968, 1974, 1976, 1977, 1985). Faunal analysis was only conducted for one of these sites –GB4 (McBryde, 1977). Graman was subsequently revisited by Boot (1990), who produced a revised chronology and technological analysis for both sites based on small additional excavations and new dating of materials from the original work. All discussion of dates from Graman in this work refers to their present calibrated ranges, generated using the SHCal20 curve in OxCal (Hogg et al., 2020).
McBryde’s (1976, 1977, 1985) faunal data were reported as MNI and derivative meat-weight estimates (reproduced here in Supplemental Information, SI 1). These data suggested major changes in the taxonomic composition of hunted game occurred during the occupation of GB4, specifically, the relative abundances and meat-weights of macropods (eastern grey kangaroo, Macropus giganteus) versus possums (Trichosurus sp.). In brief: in the first occupation phase at GB4 diet appears to have been dominated by kangaroo, but in all succeeding phases of the Late Holocene possums become relatively more numerous as does their contribution in terms of meat-weight. Other animals have a more modest representation compared to macropod and possum throughout, although the relative abundance of bandicoot seems to follow a similar trend to that of possum (McBryde, 1977).
McBryde (1976, 1977, 1985) interpreted varying kangaroo and possum abundance as causally related to a concurrent shift in technological organisation observed at GB4. Backed artefacts, that at the time were generally thought to be used in projectile weapon armatures for hunting large game, are abundant in the lowest levels but are gradually replaced after ~3300BP by ground-edged axes; used in the historical period to procure arboreal species such as possum. McBryde suggested this movement towards the Contact-era structure of Aboriginal subsistence strategies could have arisen out of a change in dietary preference, or as a response to kangaroo overharvesting (McBryde, 1977: 234–237).
Others portrayed subsistence shift at GB4 as reflecting the first integration of a novel technology (ground-edged axe) into their hunting toolkit in order to provision for increasingly large and complex communities (Morwood and Trezise, 1989). Sutton (1990) criticised both models on the basis that they framed the data in a misleading manner and used ethnographic information uncritically. However, Sutton was criticised by David (1991) for not taking into account the different durations of each occupational phase. Each interpretation produced different impressions of the timing and extent of emergent ‘possum preference’ at GB4. It is only uncontested that possums were relatively less important prior to 3250BP (or 3690–3251 calBP), than at any subsequent time. The low taxonomic resolution of the original data, however, which apparently treats all different sized macropods as M. giganteus (with an estimated meat weight of 50 kg) and all arboreal marsupials as Trichosurus possum (estimated meat weight 3 kg; McBryde, 1977: 233), offers little opportunity to explore this issue any further.
Graman B1 Rockshelter
GB1 is located 400 m southwest of GB4. McBryde excavated GB1 in two trenches which progressed from the interior of the shelter to outside of the overhang (Figure 2). As at GB4, each square was excavated in ‘levels’ corresponding to sedimentary units. At GB1, these consisted of three sedimentary units: material on the surface; a unit immediately below the surface (Level I); and a basal unit below this (Level II). In some squares Level II was excavated in two spits but others were removed in bulk. The excavations produced flaked and ground lithic material, shell, animal bone and charcoal indicative of the use of the site as a home base, although the total amount of any material type was considerably less than produced by GB4.
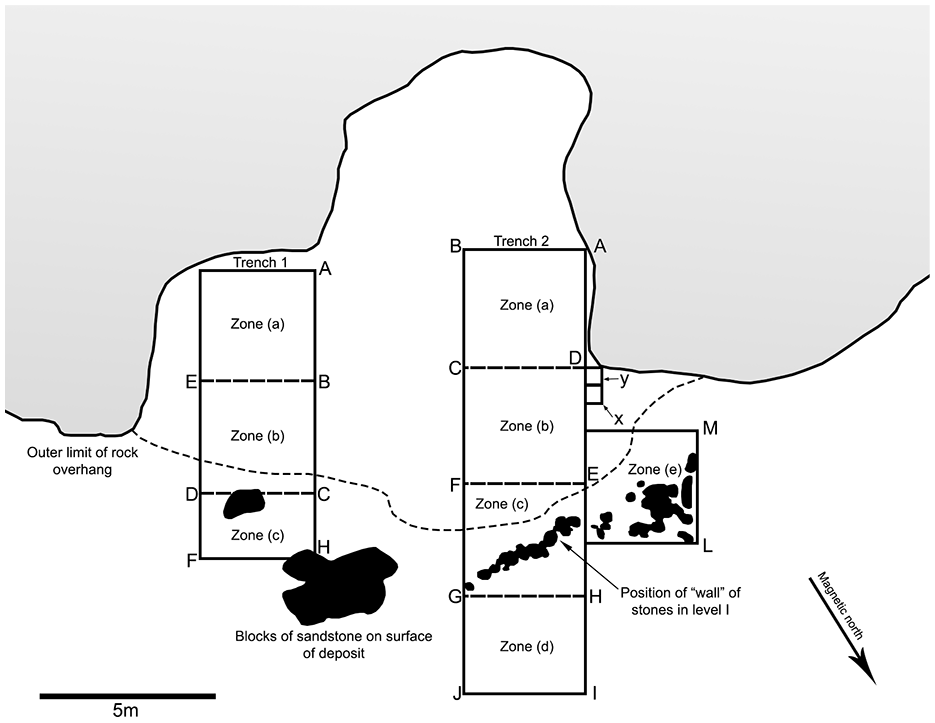
Plan of GB1 rockshelter and excavation trenches. Redrawn from McBryde (1968): 79 (2A), with addition of the additional Zones x and y excavated by Boot (1990).
Radiocarbon dates obtained by McBryde (1968) from charcoal indicated GB1 had a much longer occupational sequence than GB4. With a basal date of 5450 ± 100BP or 6398–5941 calBP (GaK-806) from the lowest part of Level II in Zone (b), Trench 1 and a most recent date of 2040 ± 70BP or 2129–1749 calBP (GaK-1187) from Level I of Zone (d), Trench 2, occupation at GB1 began approximately 1500–2000 years earlier than GB4. Materials found on the surface at GB1 are similarly assumed to derive from depositions between 2000BP and the Contact or modern periods.
The dating of GB1 has been the subject of some uncertainty and revision. There appears to be discrepancies between radiocarbon dates from the same levels from each trench (McBryde, 1974: 321). The basal date for Level II in Trench 2, taken from Zone (c), is just 3950 ± 80BP or 4575–4090 calBP (GaK-1188), 1 compared to the 1500–2000 years older GaK-806 from Zone (b) of Trench 1. Similarly, the date for Level I in Trench 2 (GaK-1187) is more recent than the date for Level I obtained from Zone (b) of Trench 1, at 4640 ± 100BP or 5581–4975 calBP (GaK-805). McBryde (1974: 321) noted that these discrepancies required further attention, but did not attempt to investigate them in later publications about Graman. She ultimately recommended that the GB1 trenches be treated separately as far as the dating and analysis of their contents were concerned (Boot, 1990: 88). Johnson (1979: 131) attempted to explain the discrepancy by suggesting that disturbances in Trench 1 had brought up older charcoal to the surface. However, as Boot (1990: 88) points out, the oldest date of GB1 was taken from charcoal sitting on bedrock so it is difficult to imagine how it could possibly have been derived from an older, lower source.
Boot’s (1990) excavation of the much smaller squares of Zones x and y (Figure 2) to retrieve new lithic samples revealed further apparent inconsistencies in chronology. Boot (1990) obtained a very early date of 7840 ± 370BP or 9538–7938 calBP (ANU-6504) on charcoal from the middle of the Level II unit in Zone y, pushing initial occupation of the site into the early Holocene. A new basal date for Level II in Zone x was also obtained from charcoal at the bedrock that dated to 1500 ± 503BP or 2750–502 calBP (ANU-6506; Boot, 1990: 89). This very small sample of highly fine-grained charcoal was explained as intrusive to the sediment it was found in, having arrived there by water action, as this square was observed during excavation to be subject to inundation (Boot, 1990: 89).
Boot (1990: 89) obtained an additional date on charcoal originally obtained by McBryde from Level II of Zone (b) in Trench 1, specifically from the northern edge of the square near Zone (c). This sample returned a date 3900 ± 240BP or 4959–3624 calBP (ANU-6503). The close correspondence of this date (error margin notwithstanding) to that of GaK-1188 from Level II of Zone (c) in Trench 2, from a similar place just outside the rockshelter overhang, seems to confirm the major discrepancy at GB1 is not between the trenches but rather between the outside and inside of the rockshelter. This is further supported by more recent dates from further beyond the overhang in Zone (d) of Trench 2: the already mentioned GaK-1187 from Spit 1 and a date of 2760 ± 65BP or 2996–2735 calBP (ANU-54) from the base of Spit 1 of Level II, Zone (d) (McBryde, 1968). That no comparable dates to these were available for Trench 1 deposits, is probably because Zone (d) sits much further outside the overhang than any material excavated from Trench 1 (Figure 2) and because no dates from Level I outside the overhang in Trench 1 were obtained.
The additional dates and review of McBryde’s earlier results given by Boot (1990: 88–90) provided a working model of the site’s chronology without a need to invoke disturbances or explain inversions. He suggested that from ~8000 to 4000BP human occupation and depositional activity primarily occurred within the overhang space, but from ~4000 to 2000BP it shifted to area outside the overhang. Considering the current calibrated ranges of published dates, initial occupation of the shelter may have occurred as early as ~9500 calBP, but as the error range for this sample is large, the median of the calibrated range of 8738 years calBP for ANU-6504 suggests it was most likely before 8000BP. Similarly, the transition to use of the outside area may have occurred early as 4500 calBP (the median of the calibrated range for GaK-1188 is 4332 calBP).
To clarify the GB1 sequence, we obtained additional dates for GB1 in the form of direct AMS radiocarbon dates on bone from several contexts. These are presented alongside the existing dates in Table 1. Some of the submitted samples failed owing to a lack of collagen preservation, these being from Level II in Zones (a) and (b) of Trench 1 and (a) in Trench 2 respectively. The four successful new dates allow for an enhanced understanding of the depositional sequence, particularly as it pertains to contexts for which McBryde or Boot did not obtain dates, whilst also raising some new questions about ongoing recent depositions.
Radiocarbon dates for GB1, organised by trench and sedimentary level and inferred occupational phases from least to most recent.
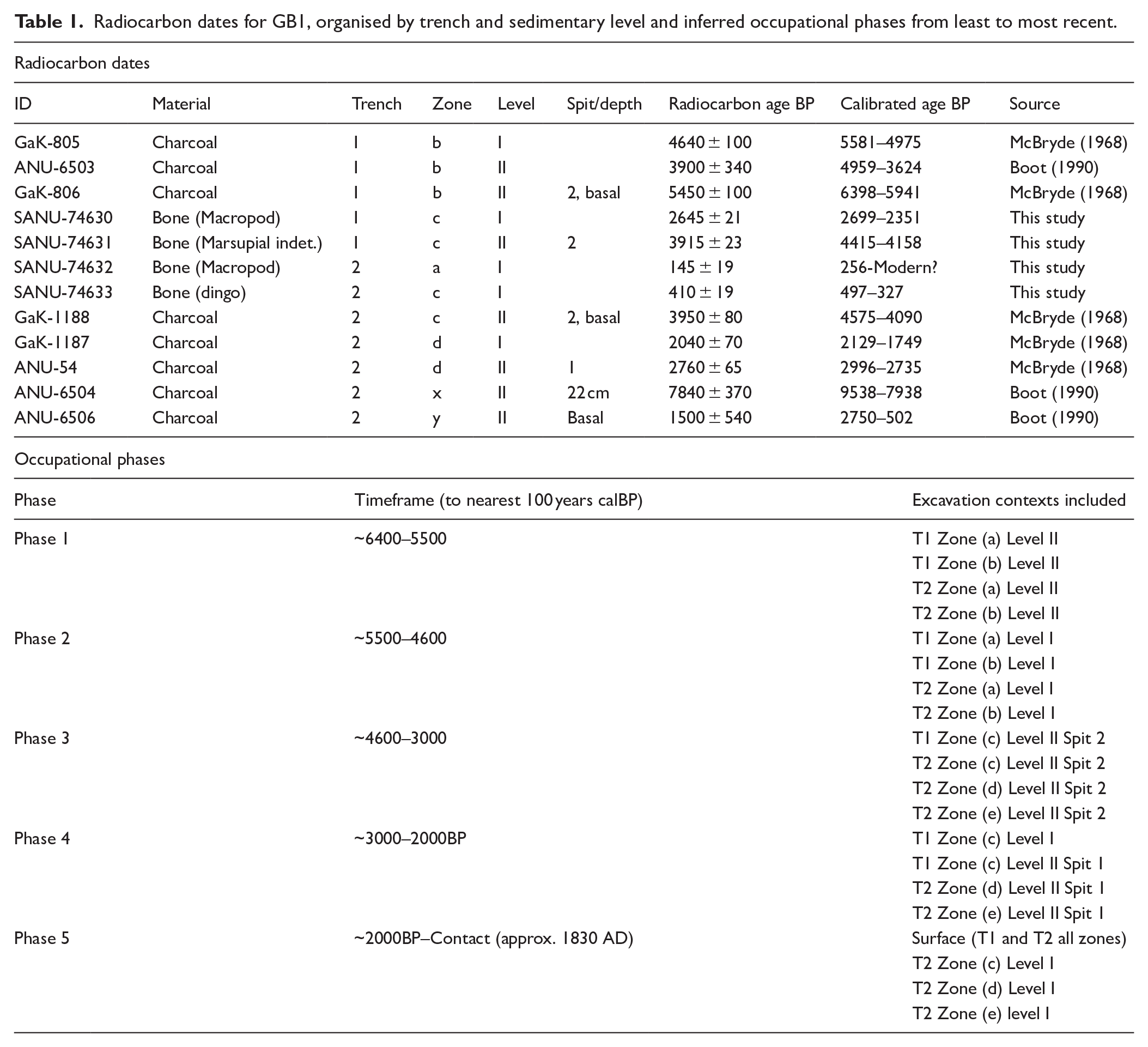
The oldest of the new dates (SANU-74631) is closely congruent with Boot’s (1990) basal charcoal date of the same unit in the adjoining part of Zone (b) (ANU-6503), and with the date from the corresponding part of Trench 2 (GaK-1188). The much-reduced error margin for the new sample indicates that the calibrated range for this sedimentary unit at the edge of the rockshelter overhang is more likely between the ~4400 and 4200 BP section of the calibrated range, rather than the 3500/4500 ends. The next oldest sample (SANU-74630) most closely matches the existing date from Spit 1 of Level II, Zone (d) in Trench 2 (ANU-54); this potentially suggests that Level I began to form directly after Level II.
A pre-Contact date (SANU-74633) is considerably more recent than all previously published dates for the site, including Boot’s (1990) anomalous charcoal date (ANU-6506). It confirms that deposition and stratification of cultural materials at GB1 continued at the edge of or outside the rockshelter overhang well into the first millennium BP rather than ceasing or being limited to surface materials as previously suggested.
The final successful sample which gave a very recent date (SANU-74632), was a fresh-looking, complete large macropod vertebra found with another as if recently articulated. Although nominally from Level I, these specimens are recent intrusions from the surface as their condition is at odds with the rest of the material from Zone (a). The result suggests this bone could have been deposited during the early European contact phase, which began locally in the late 1820s.
In assigning GB1’s excavated contexts to occupational phases for analysis of their contents (Table 2), we follow Boot’s (1990) reasoning that deposits in the interior and exterior of the shelter largely correspond to different occupational periods. The decision to assign contexts without direct dates to a particular phase is made on the basis of position relative to dated contexts (deferring to position in relation to the rockshelter overhang over Trench number), with an additional check performed through comparison of the external condition of bone between the dated and undated contexts grouped into the same phase. It must be noted that this schema is limited by the coarse stratigraphic resolution and radiocarbon boundaries provided by the original excavations and recordings. Nevertheless under the revized chronology, GB1’s faunal assemblage captures hunting activity from the Contact era to the Middle Holocene (Walker et al., 2012), with identifiable remains up to ~6400 calBP.
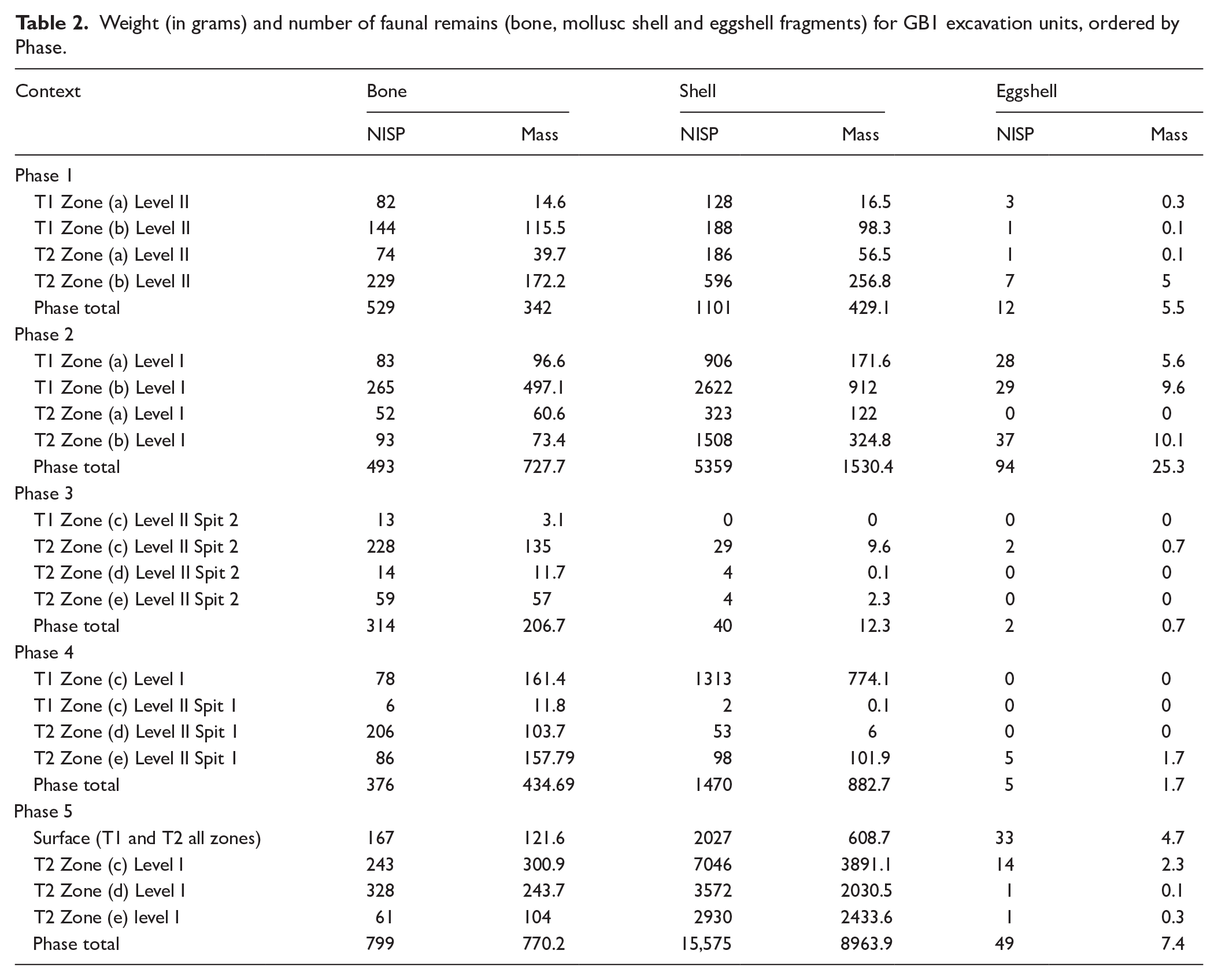
Weight (in grams) and number of faunal remains (bone, mollusc shell and eggshell fragments) for GB1 excavation units, ordered by Phase.
Materials and methods
Each bone specimen retrieved from McBryde’s excavations at GB1 was individually examined and identified to the lowest taxonomic level possible based on element type, completeness and feasibility of separating related taxa of the particular preserved anatomy (see Supplemental Information, SI 2 for information concerning Boot’s excavations). Identification efforts focussed on the mammalian component, and were conducted with the aid of the comparative Mammalogy collections at the Australian Museum, in addition to taxon-specific literature referred to in the Results. Our efforts to identify the reptile, bird, fish and gastropod remains were limited to higher taxonomic levels (Family/Order) due to low numbers and lesser diagnosticity of the specimens in question.
Our analyses of the taxonomic proportions of animals consumed at GB1 consider indicative metrics of MNI, richness (number of species) and diversity (Simpson’s Index as defined in Simpson, 1949). We also use derived body-mass as a proxy for relative dietary composition and therefore economic importance of different taxa. Sources for body-mass reference data and grouping rationale used are provided in Supplemental Information, SI 3.
Results
Overview of fauna
The number and mass of faunal specimens retrieved from each excavation context and occupational phase are presented in Table 2, with illustrative remains of key taxa presented in Figure 3. Almost all bone from GB1 is fragmentary, but a relatively high retention of diagnostic components allowed identification of 51% of all individual vertebrate remains to at least family level (and 95% of total fauna when including shell). Essentially all of the unidentified vertebrate remains appear to be mammal. Comprehensive assessment of thermal modification (Aplin et al., 2016) was not undertaken because many of the specimens, particularly those from older contexts, have adhering sand or calcite cementations which obscure their surficial condition. As a general rule, however, fresh-looking unburned bone was more common in the upper levels and outside the shelter (i.e. more recent contexts) whilst burned and calcined bone were relatively more common in the lower levels of contexts within the overhang.

Key taxa from the Graman B1 archaeological deposit. (a and b) Petrogale penicillata maxilla and premaxilla. (c) Osphranter robustus maxilla. (d) Notamacropus dorsalis mandible. (e) Bettongia gaimardi mandible. (f) Thylogale thetis mandible. (g and h) Thylacinus cynocephalus fifth metacarpal and lower canine tooth. (i) Isoodon obesulus mandible. (j–m) Canis familiaris (dingo) enamel shells of adult lower premolar, canine and carnassial, juvenile mandible with deciduous teeth. (n and o) Trichosurus vulpecula maxilla and mandible. (p) Pseudocheirus peregrinus mandible.
Macropodiform marsupials
Large macropods comprise the majority of identifiable bone specimens from GB1, but individual specimens are predominantly represented by loose molar teeth. Differentiation between the skeletal material of the two largest locally-found macropods, the common wallaroo Osphranter robustus and Macropus giganteus is difficult outside of reference to intact palates, lower incisors or the metatarsal and distal phalanx of the fourth digit (Dawson and Flannery, 1985), all of which are rare in the GB1 assemblage. However, the upper molars of these two genera can be differentiated: Macropus spp. have a strong forelink (an enamel ridge connecting the anterior cingulum with the protoloph), whilst the forelink is either absent or very weak in Osphranter spp. (Dawson and Flannery, 1985; See Figure 3 and Supplemental Information, SI 4 for illustration).
Very few of the GB1 large macropod upper molars display strong or even discernible forelinks, and so most macropod teeth discarded at the site are referable to common wallaroo. Differentiating between isolated lower molars of these two taxa is less reliable, but on the basis of the above trend, and the consistent highly curved and angular nature of their hypolophs (M. giganteus’ tend to be straighter) it is likely that the majority also belong to O. robustus. The very few large macropod distal fourth phalanges recovered from GB1 are not complete but some are sufficient to establish that a dorsoventrally curved form is present, also suggesting identification with wallaroo (Dawson and Flannery, 1985).
Almost all of GB1’s O. robustus dental material displays minimal to no wear (stages 1 and rarely 2 in McArthur and Sanson, 1988). That which does occur is focussed on the deciduous (molariform) premolar and first permanent molar, whilst the last three permanent molars are largely unworn to minimally worn. Given that the majority of these unworn teeth also show little to no root formation at the bases, they probably represent unerupted rear molars still encrypted in the mandible. Indeed in the single complete maxilla and the few reasonably complete mandibular specimens of this taxon, the third and fourth molars can be variously seen within the crypt or just beginning to emerge from it. This would suggest the majority of O. robustus individuals hunted at GB1 were between 2 and 4 years old; the fourth molar is only erupted after 7 years of age (Ealey, 1967).
Unfortunately, no more precision within this age group can be obtained from loose teeth as their state of eruption cannot be determined, and wear stages in relation to age are only provided by Ealey (1967) for the upper fourth molar (thus only applicable to animals >7 years old). Most GB1 individuals might be considered ‘adult’ as. O. robustus becomes sexually mature at approximately 2 years of age (Ealey, 1967). However, wallaroo body mass continues to increase well after this period, particularly for males which continue to grow until approximately 7 years old, whilst females plateau after 5 years (Ealey, 1967). Therefore, it is a reasonable assumption that very few GB1 wallaroos were fully grown as it pertains to musculature, and moreover that the body mass of most individuals harvested was probably in the lower half of the ~10–25 kg range.
In addition, Notamacropus rufogriseus is present in several phases in low numbers; there are also two records of N. dorsalis, and one each of N. parryi and Wallabia bicolor, all from different contexts. Most teeth from N. rufogriseus and M. giganteus appeared much more worn occlusally than the O. robustus teeth, suggesting differences in the age profiles of staple game species versus more incidentally acquired macropods (c.f. McArthur and Sanson, 1988). The relative abundances of these six species of larger macropod taxa represented at GB1 seem to be in line with modern reported densities for macropods in this area, which indicate that M. giganteus and O. robustus are substantially more common than the other, smaller species (Southwell et al., 1995). They also indicate M. giganteus is locally more common than O. robustus, probably because the former tends to live in large groups whilst the latter is essentially solitary but can form loose groups on productive land (Taylor, 1982).
The rarity of the former compared to the latter at GB1 is therefore of interest as it suggests preferential targeting of a game taxon other than the largest available. This may be because the immediate environs of GB1 are the hilly, rocky slopes and ridges typically favoured by wallaroos, although the site today is also close to flat open woodland and the data presented by Southwell et al. (1995) suggest that grey kangaroos are locally more common than wallaroos regardless of variation in terrain relief. The difference in abundance at GB1 could therefore reflect particular hunting methods or general foraging patterns which were better-suited to capturing wallaroos than eastern grey kangaroos. Potentially, this could relate to the fact that the latter are larger, faster, and by virtue of living gregariously, probably more vigiliant and able to escape from and/or defend themselves against human hunters (Colagross and Cockburn, 1993).
As identifiable maxillary/mandibular bone from large macropods is extremely rare in the GB1 assemblage, it is likely that processing activity has affected their presence in the deposit. Aside from breaking the cranium to access the brain, one possibility is that people were extracting large macropod lower incisors (which are almost non-existent at the site and only represented by broken fragments) for ornamental purposes (c.f. Balme, 1979) via percussive fracturing of the mandible. This process could also have freed unerupted rear molars from their crypts for deposition as isolated and sometimes split/broken pieces, whilst removing identifiable evidence of the mandibular housing from the immediate vicinity in a way that generic post-depositional crushing or trampling might not. There is little to no evidence of carnivore activity (c.f. Koungoulos et al., 2018) on GB1 bone material so this is unlikely to have driven maxillary/mandibular bone breakage.
Two smaller macropod species were also found at GB1, the brush-tailed rock wallaby Petrogale penicillata and red-necked pademelon Thylogale thetis. The former is more abundant and a typical inhabitant of the rocky, steep terrain found around the Graman complex. The latter typically inhabits wetter, less open forest. 2 The rarity of maxillary/mandibular specimens observed for the large macropods does not extend to these two small-bodied taxa. Dental eruption and wear patterns for both appear very variable, indicating a wide range of individual ages are represented.
Two species of rat-kangaroo (Potoroidae), the smallest local members of the subfamily Macropodiformes, are also found throughout the GB1 deposit: these are the eastern bettong, Bettongia gaimardi and the larger rufous rat-kangaroo Aepyprymnus rufescens. These taxa are readily distinguished on the basis of morphology of their premolar teeth, and whether the size of teeth in the molar row increases or decreases from the front to back. The latter species is also somewhat larger than the former (approx. 2.5–3.5 kg versus 1–2 kg). B. gaimardi became extinct on the Australian mainland in the early 20th century, whilst A. rufescens is believed to be restricted almost entirely east of the Great Dividing Range and in Queensland (Schlager, 1982).
Other Diprotodont marsupials
Brushtail possums (Phalangeridae: Trichosurus spp.) are abundant in GB1’s faunal assemblage. The vast majority of specimens are clearly referable to the common brushtail possum, Trichosurus vulpecula, although the short-eared possum Trichosurus caninus, which inhabits local wet sclerophyll forest/rainforest was also considered. These taxa are primarily differentiated on the basis of external characteristics, not easily achieved with incomplete/edentulous skeletal elements. The latter species is slightly larger and could account for a couple of the largest trichosurine specimens, which in molar row length fall at or just beyond the upper end of T. vulpecula distribution in Myers and Crosby (2023). Given the drier nature of the hilly open forest nearest to GB1 we expect most phalangerids hunted by its Late Holocene inhabitants to have been Trichosurus vulpecula.
Two species of smaller ‘possum’ were also hunted at GB1, most of which are the common ringtailed possum Pseudocheirus peregrinus (Pseudocheiridae). A single maxillary fragment represents the sugar-gliders (Petauridae), probably Petaurus australis based on overall size and premolar row length. These species are more arboreal than the brushtail possum, and would likely also have required the use of ground-edged axes to extract them from trees.
Agreodont marsupials
Bandicoot (Peramelidae) remains are common at GB1. The majority of identifiable specimens are the now locally-extinct southern brown bandicoot Isoodon obesulus, with a considerably smaller proportion of long-nosed bandicoot Perameles nasuta. Most identifiable bandicoot remains from GB1 are mandibles, and as Perameles crania and mandibles are more gracile than those of Isoodon spp. they are somewhat less likely to be preserved in a state of completeness conducive to identification.
A range of small carnivorous marsupials (Dasyuridae) were found at GB1. From the Dasyuridae are included one partial mandible of antechinus (possibly Antechinus flavipes), and specimens of eastern quoll Dasyurus viverrinus and tiger quoll Dasyurus maculatus, the latter of which is mainland Australia’s largest extant marsupial carnivore. They are amongst the least common marsupials in the assemblage, but their presence may nonetheless be considered notable as they were not frequently noted in the historical diet of Aboriginal people.
More interesting are two specimens of thylacine, Thylacinus cynocephalus (Thylacinidae): a complete left fifth metacarpal from 1(b) II and a fragment of lower left canine from 1(a) I Spit 1 (Figure 3). Thylacines became extinct on the Australian mainland by 3200 calBP alongside the Tasmanian devil Sarcophilus harrisii (Dasyuridae; White et al., 2018), so their presence is useful for relative dating of the GB1 strata they are found in. Thylacine remains are extremely rare in archaeological sites (Knights and Langley, 2021), and the finding of this species at GB1 is significant as the first such record from a large area of eastern Australia.
Placental mammals
The remains of at least two dingoes (Canidae: Canis familiaris or Canis dingo), both immature, are preserved at GB1 in Trench 2 (Figure 3). One individual is a juvenile represented by a right partial mandible from Level 1 of zone (c), with two deciduous premolars indicating an individual age of less than 8 weeks. A microCT scan of the specimen showed the adult carnassial tooth within a crypt below the extant deciduous dentition. This individual is the source of the new AMS date SANU-74633 (410 ± 19BP, 497–327 calBP). The second individual is a young subadult represented by a series of teeth also found in Level 1 of the adjoining zones (c) and (e). The teeth are adult (permanent) teeth consisting of hollow enamel crowns without any root formation, indicating an early stage of development, recent eruption and a probable individual age of less than 6 months.The apparent derivation of the loose teeth from a single individual is useful in determining the relative antiquity of faunal remains in Level 1 of Trench 2 zones (c) and (e). Dingoes breed once annually, with pups born in May or June. The presence of juveniles with deciduous teeth or very new adult teeth (i.e. less than or not much more than 8 weeks old) is therefore indicative of the usage of GB1 during winter to early spring.
Remains of rodents (Muridae) are rare. The few identifiable craniodental elements belong to native bush rat Rattus fuscipes and a species of Pseudomys, probably P. novaehollandiae. The rarity of these animals, as well as others of similar size suggests that no owl roosting took place at GB1, although it is possible that sieves used to process sediment did not have a fine enough mesh to capture them. The larger ‘rabbit-rat’ Conilurus albipes is represented by two fragmentary mandibles and is likely to represent human prey. A small number of bones of domestic sheep (Bovidae: Ovis aries) and one instance of rabbit (Leporidae: Oryctolagus cuniculus), both introduced by Europeans within the last 200 years, were identified from the surface of the deposit. These bones are very clean and complete, lack signs of human modification and probably derive from recent natural deaths.
Other animals
Bird bones are very rare in the GB1 deposit, and nearly all seem to be from very small passerines with a clean, fresh appearance, which may represent non-cultural inclusions resulting from natural deaths. No effort was made to further identify these. Fragments of emu (Casuariidae: Dromaius novaehollandiae) eggshell were found in all zones, at low frequencies. A lack of identifiable emu skeletal remains suggests that exploitation of this species extended only to the collection of eggs from nests. Emus lay eggs from May to June which take about 2 months to hatch, suggesting that occupation of GB1 probably occurred during winter to early spring.
Reptile bones were recovered throughout the GB1 deposit at low frequencies. Of these, shell fragments from at least one species of freshwater turtle (three species are found locally, all in the family Chelidae) and vertebrae of snakes in the families Elapidae or Colubridae are the most abundant. No effort was made to identify these further. A few snake vertebrae are reliably attributed to Pythonidae, and their size and morphology suggests they are likely from carpet python Morelia spilota. Lizard remains are very rare. The few diagnostic specimens are referable to monitor lizard (Varanidae) – probably lace monitor Varanus varius – two species of skink (Scincidae) in the genera Egernia and Tiliqua (blue-tongued lizard), and one species of dragon (Agamidae), probably Amphibolurus sp. Almost all of these are single occurrences.
Fish represents a very small portion of the faunal assemblage, with nearly all identifiable elements being poorly-preserved vertebral fragments (n = 8). No attempt was made to identify these remains. Their size and general appearance suggests they could come from golden perch (Percichthyidae: Macquaria ambigua) or freshwater eel-tailed catfish (Plotosidae: Tandanus tandanus) which are some of the larger-bodied taxa in the Graman area’s waterways.
Freshwater molluscs are well-represented throughout the GB1 deposit. The vast majority are freshwater mussels (Hyriidae), referable to Alathyria jacksoni and Velesunio ambiguus. These taxa are both very closely related (the former is nested within the Velesunio complex genetically) and highly similar morphologically, in addition to exhibiting high phenotypic plasticity (Baker et al., 2004). Considering this and the incomplete condition of the vast majority of shell fragments from GB1, almost all of the mussel shell was only referred to family level.
Other molluscs present include at least three species of very small aquatic gastropods from the families Tateidae and Thiaridae (trumpet snails), in addition to one small bivalve in the genus Corbicula (Cyrenidae). They are found in relatively large numbers in some GB1 contexts, an observation made from freshwater middens elsewhere in NSW, where such small molluscs are often numerically dominant over larger mussels. Given their tiny size it has been speculated that they served some flavour-improvement or possibly medicinal purpose (Garvey, 2017), though elsewhere in Australasia very small gastropods are boiled in large numbers and eaten using picks to extract the meat (S. O’Connor pers. obs., Wetar, Indonesia). Intact gastropods might have entered the deposit through people cleaning innards of fish that are known to eat small aquatic snails whole (namely catfish) though the scarcity of fish bone at GB1 suggests this is unlikely.
Trends in taxonomic representation at GB1
MNI data for identified taxa are presented in Table 3. Most bone material at GB1 comes from the marsupial species still extant in the Graman area today, with much smaller numbers of placental mammals, reptiles, birds, fish and molluscs. Total faunal content deposited increases between Phases 1 and 2, declines substantially in 3 to make a small recovery in 4, then resurges sharply to reach a site maximum in Phase 5. MNI is dominated by molluscs except for Phase 3, with mussels as well as the much smaller snails and clams reaching their maximum numbers in Phases 2 and 5. Mammals are dominant during Phase 3, and elsewhere are the next most abundant group after molluscs. Thylacine is present only during Phases 1 and 2, and has no overlap with dingo which only appears in Phase 5. Aside from turtle, all birds, reptiles and fish are at all times very poorly represented both within and between phases, and can only be considered incidental inclusions.
MNI of identified taxa at GB1 by phase.
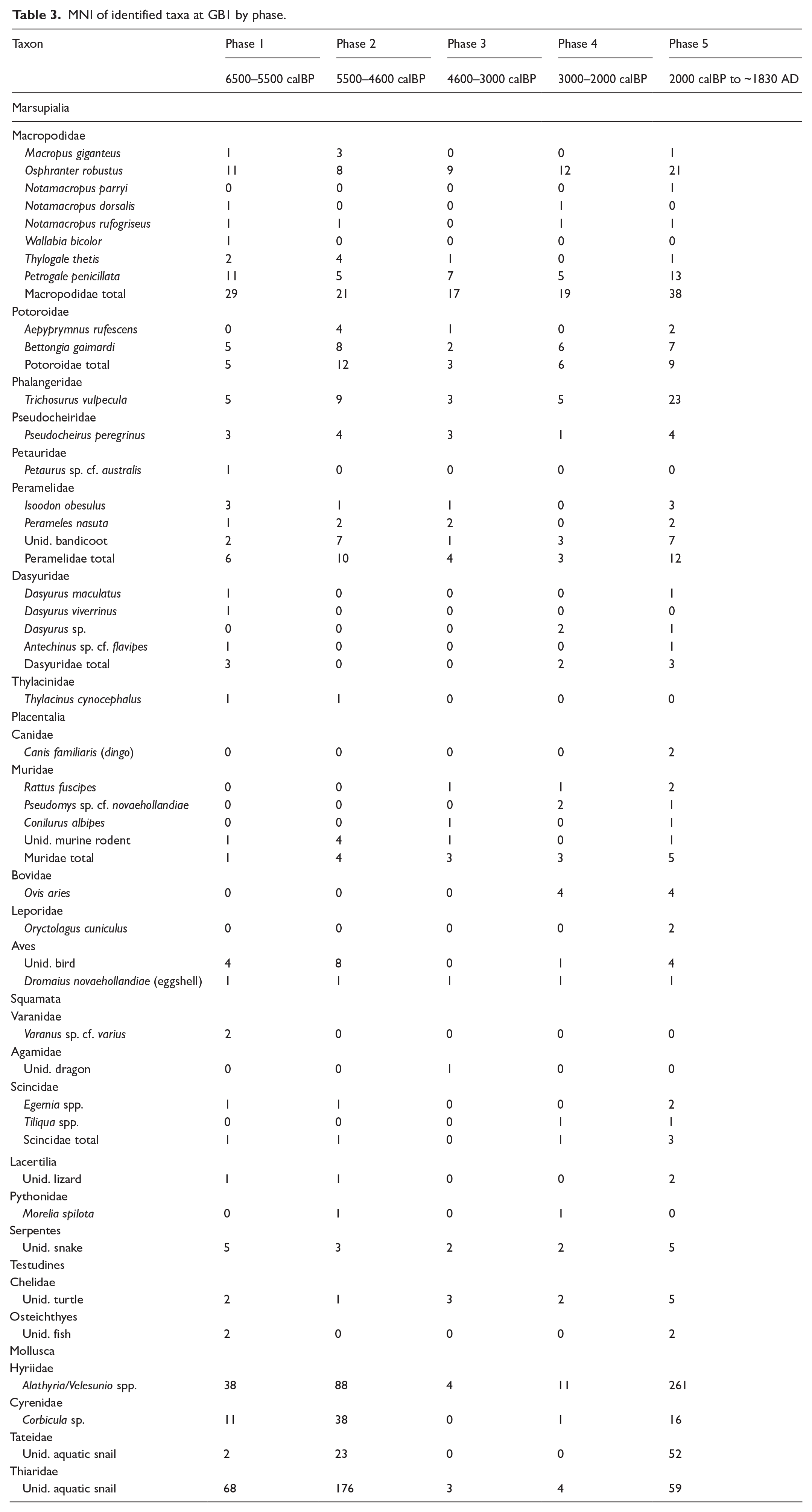
The range of species present indicates extensive exploitation of the wooded, hilly environments and their creeks which surround the Graman site complex, during all periods. The presence of Thylogale thetis indicates hunting at forest edges, where this transitions to a flatter plain of mixed open woodland and grassland, but rarity of M. giganteus, N. dorsalis and N. parryi suggests limited utilisation of said areas which lie beyond the hilly escarpment confining the Graman complex. Macropod taxa present during Phase 3 suggest that utilisation of this area diminished further at this time.
Relative proportions of each major mammal group by phase (Figure 4a) reveal a number of interesting trends. Macropods dominate MNI at the family level, their representation never dropping below 30%. However they become markedly less numerous in Phase 2, before surging to reach a peak in Phase 3. Large macropods specifically are most numerous in Phases 3–4. The remaining mammals – all small taxa except for exceptional cases of thylacine – were collectively most important during Phase 2.
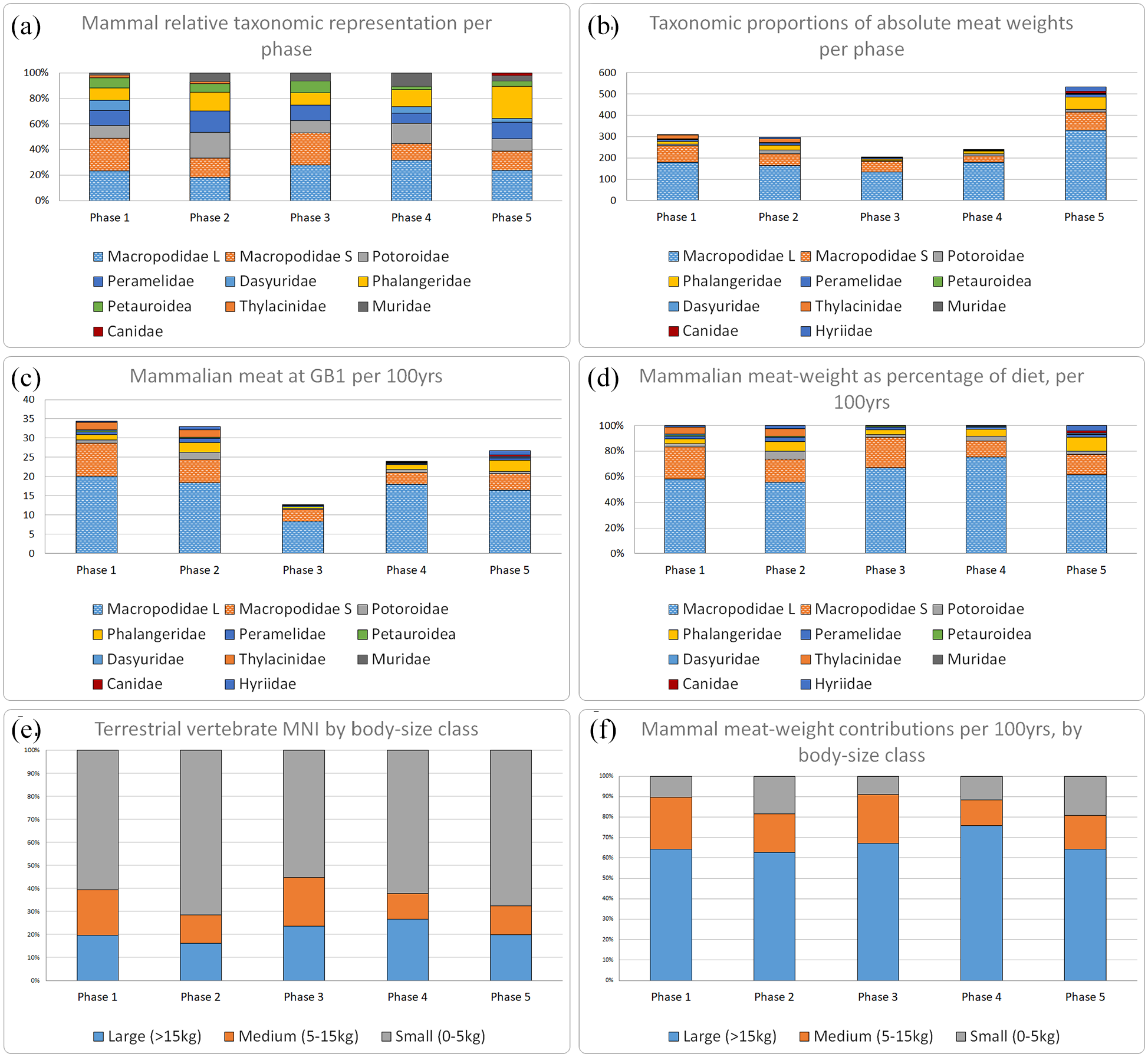
Chronological trends in animal frequency and body-mass at GB1. (a) Proportion of MNI per phase; (b) total body mass of major mammal groups and mussels processed during each phase at GB1, and (c) corrected for phase duration (kg/100 years); (d) proportions of body mass of major mammal groups and mussels processed during each phase at GB1, corrected for phase duration; (e) Proportion of terrestrial vertebrate MNI by body-size; (f) Proportion of mammal meat-weight by body-size.
The proportion of arboreal marsupials (i.e. Phalangeridae and Petauroidea) sits between a relatively minor and stable range of ~15–20% of total from Phases 1 to 4, but in Phase 5 expands considerably in representation, mostly driven by brushtail possum. However, abundances of the arboreal marsupials relative to small terrestrial mammals specifically (i.e. Potoroidae, Peramelidae, Dasyuridae and Muridae) appear to consistently favour the latter at ratios of 1:2 or 1:3 until Phase 5, where they reach approximate parity. Relative to macropods, the MNI of smaller mammals is similar between their peaks in Phases 2 and 5, but the arboreal component specifically is much greater in Phase 5.
Body mass and dietary importance
Chronological trends in absolute meat-weights (Figure 4b) generally follow those of MNI in identifying a major decline in animal-bone deposition at GB1 in Phase 3 from ~4600 to ~3000 calBP, followed by a recovery in the subsequent periods and a peak reached in Phase 5. However, when weight data is considered controlled for duration (kg per 100 years, Figure 4c) it becomes apparent that, during the earliest two occupational periods with corresponding fauna (Phases 1 and 2), relatively greater amounts of meat were processed at GB1 than in any subsequent period. As with MNI there is a noticeable decline in the total amount of meat processed/consumed during Phase 3 which gradually recovers during the subsequent periods.
The animal diet at GB1 was at all stages dominated (~75–90% of any one phase) by macropods, particularly large macropods (~55–75% of any one phase), as indicated by proportions of game body mass corrected for phase duration (Figure 4d). The greatest proportional representation of macropods is in Phase 3. Although vertebrate MNI is generally >75% small or medium taxa (Figure 4e), this macropod dominance translates to majority proportions (>60%) of meat derived from large mammals across all phases.
As the meat-weight value of 15 kg used here for large macropods is very possibly an underestimate, given that this is at the lower end of adult size for O. robustus and M. giganteus, the true proportion may well have been even more in their favour. Relative contributions of the smaller macropod species seems to have been relatively stable but somewhat lowered in the latest Holocene periods (Phases 4 and 5). Despite the large numbers of mussels deposited their actual contribution to the diet is low at all times, even in Phase 5. Mussels were supplements to the GB1 diet rather than staples, as in many other coastal or riverine sites in eastern and southeastern Australia.
Dietary importance of non-macropod mammals was greatest during Phase 2, comprising approximately 25% of total game by body mass. During this period all non-macropod taxa other than brushtail possums reach their maximum contributions to the GB1 diet. Conversely, the importance of brushtail possums specifically was by far greatest in Phase 5, corresponding to the last 2000 years. There is no commensurate increase in ringtail possum or sugar-glider, which were uncommon at all times. No noteworthy trends are apparent for other mammals, aside from an apparent correspondence between increased contributions from smaller non-macropod mammals and mussels occurring in both Phases 2 and 5, whilst the contributions of both are absolutely minimal during Phase 3.
Given the very low number of birds and reptiles identified at GB1, and the much smaller masses of meat each would yield compared to all but the smallest mammals, their inclusion into considerations of meat-weight would not meaningfully change any of the above observations.
Richness and diversity
Chronological trends in richness and diversity for non-molluscan fauna are similar for both measures (Table 4). The high richness and diversity seen in Phases 1 and 2 collapses in Phase 3 and does not fully recover in Phases 4 and 5. Phase 5 is actually least rich and diverse; this appears to be because representation within most popular mammal game categories have become focussed around one species each, despite the large breadth of taxa harvested overall (richness). O. robustus dominates the large macropods, P. penicillata the small macropods and T. vulpecula the arboreal/smaller marsupials, indicating most hunting productivity was derived from the rocky, tree-covered hills and gullies running north and south of the Graman complex. Utilisation of the flatter plain beyond this zone was by comparison increasingly limited. This might be a signal of specialisation in hunting these particular taxa having developed within the last 2000 years, though it is worth noting that the high richness/diversity of Phases 1 and 2 are partly derived from numerous taxa which only appear once.
Diversity and richness of fauna per occupational phase to three significant figures. Measures are also given as proportional to phase duration, based on the assumption that greater time elapsed provides greater opportunity for a taxon to be sampled.

Discussion
Nature and timing of Holocene subsistence change in the Graman area
Our analysis of the GB1 fauna provides a general chronology for changes in hunting habits and subsistence over the last ~6500 years in the Graman area. During the earliest occupation of the site for which bone is preserved (Phase 1, 6400–5500 calBP), the diet of GB1’s inhabitants was mostly comprised of macropod, supplemented by a selection of smaller terrestrial and arboreal mammals. The range of species present indicate foraging predominantly within local open forest hill environments. During the subsequent Phase 2 (5500–4600 calBP), the inhabitants of GB1 made increased use of a relatively larger range of small animals, and correspondingly, their lowest use of macropods at any point of the site’s occupation. The specific reduction in macropod hunting relative to Phase 1 appears to have been in the smaller members of this group (Petrogale and Thylogale spp.).
Phase 3 (4600–3000 calBP) was a markedly different period exhibiting reduced depositional intensity and very low faunal diversity, with hunting almost totally dominated by macropods and particularly the large taxa within this group. Importantly, the onset of Phase 3 coincides with the earliest known occupation of the nearby site GB4 (4575–4090 calBP), where at this time, the faunal remains are also apparently predominantly comprised of macropods (McBryde, 1977). It is unclear whether the lowered depositional rate of Phase 3 reflects lowered local abundances of small animals and a hunting strategy focussed on high-yield large game, or whether it simply reflects the inhabitants of Graman moving their main living space to GB4. Boot (1990) also came to the same conclusion of subsistence activity relocation to GB4 based on other indicators of site occupation (lithic discard). There is no obvious reason in terms of location or rockshelter morphology why the latter might have been motivated. One possibility is that social or ceremonial reasons may have driven more intensive use of GB4 after 4500 calBP, as this site contains painted rock art whereas GB1 does not.
In Phase 4 (3000–2000 calBP) human diets at GB1 were still dominated by macropods, but this period saw reintroduction of small mammals, brushtail possum and potoroids in particular, to the inhabitants’ diet. The final period of occupation, Phase 5 (2000 calBP to early 19th century AD, i.e. Contact) exhibits a further heightened reliance on small marsupials, now focussing to an even greater degree on arboreal possums. The abundance in numbers of faunal remains recovered from contexts corresponding to this period contrasts with low taxonomic diversity, suggesting specialised exploitation of O. robustus and P. penicillata in addition to Trichosurus vulpecula. Exploitation of freshwater mussels also peaked at this time, but still formed only a minor dietary component.
The patterning of animal representation at Graman B1 is similar in some regards that of GB4 as presented by earlier published analyses. Overall, macropods and especially large macropods contributed the greatest share of meat processed at GB1 in all phases. This trend reached its peak between 4600 and 3000 years calBP – where it coincides with a similarly skewed taxonomic division at nearby GB4. It appears that during this time people tightened the range of animals they hunted to these most high-yielding of the locally available game taxa.
By comparison, periods of relatively greater reliance on smaller animals occurred on either end of Phase 3. However, the actual significance in fluctuations of small animal representation at GB1, in terms of what they communicate about hunting strategies at different times remain unclear. At no point during the occupation of GB1 did small animals ever provide a greater or even equal share of meat than macropods. This contributes to one of the starkest differences with GB4: at GB1, the last 2000 years of occupation is still dominated by macropods, rather than possums, despite the latter increasing in representation both against the total diversity of fauna exploited as well as within the small-mammal and arboreal marsupial categories.
We caution that these comparisons to GB4 are only indicative, as they take the GB4 faunal data at face value to be accurate and of sufficient detail to capture prevailing trends. The greater diversity of species now known from GB1, and the predominance of O. robustus over M. giganteus, suggests that this is unlikely to actually be the case, warranting formal reidentification and reanalysis of GB4’s faunal material itself. For instance, the premise that the last 2000 years at GB4 only featured possum and no macropod is based on identification of only cranial material found on the surface, and seems unlikely to withstand scrutiny.
One of the problems facing interpretation of GB1 is the punctuation of a trend of established small game exploitation by heightened macropod reliance, unfortunately coupled with reduced deposition. In the absence of existing data from GB4 showing considerable occupation beginning there at the same time, this may have been interpreted as partial abandonment of the area. Instead, it may simply reflect reduced use of GB1 following relocation of animal processing/consumption activities to a more favoured site nearby. This finding highlights the importance of multiple-site frameworks in developing accurate understandings of local and regional chronologies. It also emphasises the need to revisit and reanalyse GB4’s faunal materials to ensure that the taxonomic proportions of each phase are accurately known, which will inform whether the composition of Phase 3 as it appears at GB1 is a realistic reflection of diet at the time or skewed by reduced depositional activity.
Significance and factors of Holocene subsistence patterns in eastern Australia
The most important finding from our zooarchaeological analysis of GB1 is the clear lack of evidence for a significant shift developing in the Late Holocene away from macropods and towards a focus on smaller animals. This alone challenges the notion that substantial change in Aboriginal Australian diets necessarily occurred during the latter half of the Holocene, and contributes further to the growing picture of developments in animal diet composition occurring on a regional rather than continental basis. Additionally, the consistent, and at times intensifying use of macropods during the Late Holocene at GB1 does not appear to align with the typical predictions of various models invoking the effects of changing climate, dingoes, new hunting technologies, and human population size/social organisation.
The periods during which non-macropod vertebrates made their greatest contributions to diet (Phases 2 and 5) are not consecutive, being separated by several millennia with a period of near-total macropod reliance in between. Hence, it seems that the fluctuations seen at GB1 are likely to be driven by a combination of different phenomena, and cannot be explained by a simple causal model. While it is beyond the scope of this paper to examine in depth the implications of GB1’s results for each model of change, is nevertheless worth considering here whether any of their proposed factors may be related to the changes which are observed at GB1.
Climate and ENSO-related aridity
Climate change, and its impacts on rainfall, has long been the primary external driver cited as a driver of Holocene cultural changes in Australia. Specifically, the El Niño-Southern Oscillation (ENSO), a climatic phenomenon with effects in eastern Australia amounting to unpredictable, often arid and drought-inducing conditions, which caused fluctuating and depressed resource availability for Australian foragers (Hiscock, 1994). Large macropods, for example, suffer high mortality rates under drought conditions (Caughley et al., 1985; Robertson, 1986), which would be expected to significantly impact subsistence regimes reliant on them. Following a period of above-average moisture in the early Middle Holocene (c. 9000–6000BP), the onset of intensified ENSO conditions is generally considered to have begun between approximately 5000 and 4000BP (Marx et al., 2009; Reeves et al., 2013; Shulmeister and Lees, 1995; Williams et al., 2015). Subsequently, from 2000 to 1500BP, there then appears to have been a general amelioration characterised by more regular warmer, wetter La Niña conditions (Williams et al., 2015). Some interpretations characterise a two-step ENSO intensification process, with the initial onset of aridification at ~5000BP and intensified drying from ~3000 to 2000BP (Asmussen and McInnes, 2013).
However, there exists considerable variation within palaeoenvironmental data as to when precisely the moister period gave way to drier conditions, and how long these lasted, in any particular locality (Donders et al., 2007), even within relatively small portions of mid-latitude, subtropical to temperate eastern Australia that Graman is part of. This is probably due to strong local biases resulting from variable factors in ‘geology, catchment size, altitude, topography, vegetation, fire history, lake water depth and water chemistry’ (Hembrow et al., 2017). The nearest recently-published record to Graman is from Little Llangothlin lagoon (approx.132 km southeast of Graman), where plant macrofossils indicate that the period between 6100 and 1000BP was drier than that from 9300 to 6100BP (Woodward et al., 2014). However, this site does not provide higher-resolution information as to variation within this period, in other words, when GB1 was occupied, meaning that other sources must be consulted. Geochemistry of acid peats at Carlo Creek, 415 km to the northeast of Graman, is indicative of a wet period between 4500 and 2500BP which was preceded by a drier period beginning in the Holocene (Krull et al., 2004). At Bundjalung National Park, 220 km east of Graman, pollen records conversely indicate a wet period from 7500 to 6400BP, followed by drying until 3200BP, then a return to slightly wetter conditions leading to 700BP (McGrath and Boyd, 1998). Another available record from the mainland comes from Barrington Tops, approx. 310 km south of Graman. Here, pollen suggests increased moisture relative to today from 7800 to 3700 calBP, with an apparent two-step aridification beginning around 5000 calBP, increasing from 3100 calBP onwards until finally stabilising around 1400 calBP (Dodson et al., 1986).
The most detailed palaeoenvironmental records for this region of Australia come from large sandy islands lying closely off the east coast, in particular Minjerribah (North Stradroke Island, approx. 325 km from Graman). Here, assessments of aeolian sedimentation from Native Companion Lagoon on Minjerribah supposed a pronounced increase in aridity from 5700 calBP, peaking at twice at 4690 and 3890 calBP before decreasing at around 2000 calBP (McGowan et al., 2008). However, at Swallow Lagoon on (North Stradbroke island), carbon-isotope ratios from Melaleuca quinquenervia leaves indicate that the wet period from the beginning of the lagoon’s record at ~8000 calBP shifted only after 3200 calBP to ENSO-driven drier conditions (Barr et al., 2019). A weaker ENSO from 5500 to 3500BP on Minjerribah is also supported by oxygen isotopes from diatoms at Blue Lake (Maxson et al., 2022).
Some analogous studies from K’Gari (Fraser Island, approx. 500 km from Graman and less than 150 km from Minjerribah) such as at Lake Jennings also indicate significantly drier conditions after 3500BP (Hembrow et al., 2017). Conversely records from several other lakes on K’Gari variably suggest periods of aridity from 9500 to 6300 calBP, stabilisation of low rainfall until 5100 calBP and then rises in rainfall until 2600 calBP (Hidden Lake; Longmore, 1998); or aridity from 6100 to 2500 calBP (Lake McKenzie; Atahan et al., 2015); and finally aridity from 6500 to 5400 calBP followed by moist conditions until 3000 calBP (Lake Allom; Atahan et al., 2015; Donders et al., 2006).
The general contradictions in these records and their implications for attempting to interpret human behaviour from the archaeological record over the same timescale are self evident. This is made more difficult by the uneven use of radiocarbon date calibration and in some cases inferred rather than direct ages for transitional boundaries (derived from age-depth curves). The nature of the last 2000–1500 years is also unclear. Most analyses agree that it was generally less arid and more stable, but some (also based on Minjerribah records) posit that mean rainfall was lowest during after 2000 calBP and that this late period was also the most variable at an interdecadal level (Cadd et al., 2024). Such inconsistencies make the application of broad models of climatic change derived from specific localities to other places troublesome, offering little confidence in their robusticity as analogues.
Accordingly, for the purposes of this paper we opt to defer to the most local palaeoenvironmental record available, which is derived from pollen analysis stemming from McBryde’s excavations. The palynological data from GB1 and other sites from within the Graman complex indicate the extent and direction of local environmental changes during GB1’s occupation (Bowdery, 1984; Sandor, 1969). They suggest that between 5500 and 4600BP (i.e. during the transition from Phases 1 to 2), grasses tolerant of drier conditions increased in abundance around the Graman complex. This presumably indicates the onset of relatively drier conditions locally, and seems to coincide with the period of greatest small-game utilisation and lowest macropod exploitation. Conceivably this could signal drought-related suppression of (medium-sized) macropod stock, and hunters’ heightened reliance on lesser-affected smaller game taxa as alternatives. Curiously, however, this also coincides with a steep rise in abundance of aquatic molluscs compared to Phase 1. This might be interpreted evidence contrary to increased aridity, although we note that both the mussels and small freshwater snails remain widespread and abundant also in riparian semi-arid environments (Garvey, 2017). It is possible that increased mollusc reflects altered foraging choice, in line with the greater diversity of other animals exploited at this time, rather than any environmental differentiation.
There is some palynological indication of increased water availability near one of the other Graman sites after 3000BP (=Phases 4–5), but otherwise there seems to be little evidence of further vegetation change after 4600BP (Boot, 1990). This would suggest that changes in the patterning of animal consumption subsequent to Phase 4, which include a resurgence in macropod hunting in Phase 3, did not accompany substantial change to the floristic communities supporting local vertebrate game. If the amelioration of ENSO and the onset of broadly warmer, wetter climatic conditions in eastern Australia after 2000BP has some relation to the specialised hunting of O. robustus, P. penicillata and T. vulpecula it does not appear to be causally reflected in any structural change to vegetation around Graman.
The evidence for climate variation impacting hunting patterns at GB1 is therefore equivocal. The general impression is that local vegetational change was minor and that a shift was only effected once, and it is difficult to identify any obvious correlates with further faunal change either at an individual taxonomic level or more broadly. This is a useful observation, however, as it perhaps reflects the long-term stability of environmental support for the large and medium-sized macropod game that the inhabitants of Graman continually relied upon through this and the following periods. A novel reanalysis of pollen and other palaeonvironmental indicators from the Graman complex sites’ excavated sediments, using modern methods is in order to further investigate this issue.
Arrival of dingoes
The dingo arrived in Australia sometime shortly prior to 3300 calBP (i.e. late Phase 3), by which time it had dispersed throughout the mainland of the continent (Balme et al., 2018). Based on inference from ecological and ethnographic data, it is suggested this event could have increased peoples’ reliance on small and medium-sized game. This occurred through a combination of predatory competition for large game (Fillios et al., 2010, 2012), and their use as tamed hunting assistants which altered the composition of hunting yields in favour of smaller taxa perhaps by increasing opportunistic encounters with such animals (Balme and O'Connor, 2016: 778; Koungoulos, 2017). At GB1 there is certainly not any evidence that local macropod stocks were meaningfully depleted by dingoes during Phase 3, since their portion of dietary composition only increases relative to Phase 2. There is a slight increase in small game representation subsequent to the dingo’s arrival in Phase 3, although this never again reaches the levels observed in what we assume to be the pre-dingo period (Phases 1–2). It is therefore difficult to determine whether this represents a gradual return to a previous norm after an unusual, disruptive Phase 3, or the influence of a novel factor.
An alternative scenario is that early tame dingoes were used to procure medium and large-sized macropods in communal drive hunts (Koungoulos and Fillios, 2020). This type of hunting seems to have historically occurred with greatest frequency in the forested, hilly middle latitudes of eastern Australia (Koungoulos and Fillios, 2020), with examples including the areas close to Graman itself (McPherson, 1860 in McBryde, 1976: 207–210); many reports remark on the capability of such hunts to acquire large numbers of macropods in a single episode. It is therefore possible that tame dingoes may have been used to facilitate the relatively intensified hunting of macropods during or subsequent to Phase 3. Of particular interest is Phase 5, which leads into the historically-observed cases above, and where specialised hunting of one medium (P. penicillata) and one large-sized macropod (O. robustus) in particular is inferred from their relative taxonomic dominance. There are two means by which the potential scenario of dingo involvement in faunal acquisition could be tested, although neither are conclusive.
The first is seeking evidence for the presence of domestic dingoes at GB1. This proves to be very scarce, aside from the late Phase 5 remains of individuals which may have been game themselves. A very small proportion of macropod bones (n < 10) from GB1 bear evidence of carnivore tooth-marks that are probably from dingoes, although an alternative attribution to thylacine cannot be ruled out at this time. Although tooth-marks are generally found on a very small proportion of bone, there is also no sign of the extensive gnawing, destruction of bone and redeposition of acid-digested bone through scat that might be expected from a resident population of tame ‘camp’ dingoes (Solomon and David, 1990) regularly used in hunting over several millennia. In the referenced historical accounts of hunting in the region of eastern Australia that includes Graman, the species of ‘kangaroo’ or ‘wallaby’ hunted with dingoes are not specified. ‘Kangaroo’ is commonly used in vernacular to denote any macropod, and in this diverse region of the continent could refer to anything from M. giganteus to T. thetis. As such it is not possible to infer from the angle of taxonomic representation whether dingoes were more or less likely to be involved in the acquisition of macropods at GB1 via large-scale communal drives. Given the above uncertainties, our assessment is that a potential role of dingoes in human dietary change at GB1 remains at best unclear.
Novel extractive technologies
During the latter half of the Holocene, two novel lithic technologies – backed artefacts and ground-edged axes – proliferated throughout much of the Australian mainland (Hiscock and Maloney, 2018). Each technology has been linked to specialised hunting practices of macropods and arboreal marsupials, respectively including specifically in the context of Graman (McBryde, 1977). Backed artefacts in particular have also been associated with more generalised, multi-functional foraging or processing capacities (Hiscock, 1994). Both technologies occur at Graman; abundances for each are given in the Supplementary Information, SI 5. Backed artefacts occur in Phases 1 and 2 at GB1, but they are far more abundant in Phases 3 and 4. Interestingly, backed artefacts still occur in reasonable numbers in some of the uppermost units of Trench 2, which are now understood to include evidence of occupation within the last 500 years; elsewhere, their manufacture and use is believed to cease after approximately 1500BP (Hiscock, 2008).
It is tempting to interpret a connection to the sharply increased importance of macropods in these periods compared to Phases 1–2, and sustained macropod hunting into Phase 5. However, it is not clear precisely when within this last period that backed artefacts ceased to be used and hence when the two phenomena may have become unlinked. Additionally, it is of interest that the proliferation of pointed forms in particular at GB1 lags the earlier efflorescence of geometric microliths (more symmetrical) forms, only appearing in abundance in Phase 5 contexts (Supplemental Information, SI 5). Boot (1990: 167) through experimental use-wear replication concluded that ‘most of’ GB1’s geometric microliths were used as barbs in spear armatures; but points were more commonly used for multiple other functions including cutting and scraping. Although the data are coarse our results tentatively support the notion that intensity and relative importance of macropod hunting at GB1 coincided with the peak of geometric microlith production. We do not find evidence that backed artefacts are associated with greater dietary breadth (using richness and diversity as proxies) at least as it pertains to taxonomic representation of vertebrates, because these measures are greatest before the most significant production of said technology at GB1. However, these concepts require further formal statistical investigation and integration of data from other sites that also contain both backed artefacts and well-preserved fauna, which will be the subject of future work.
There are no intact examples of ground-edged axes within the GB1 deposit (Boot, 1990), although fragments apparently struck from them are found in levels corresponding to Phases 2, 4 and 5 as are portable grinding slabs potentially used in their manufacture or maintenance (Supplemental Information, SI 5). Based on their chronology at GB4 ground-edged axes probably first entered use locally between 4575–4090 and 3690–3251 calBP, but certainly by the lower end of the second calibrated range (Boot, 1990). Historically, ground-edged axes were used for several tasks including, importantly, ascending trees to capture arboreal marsupials, as well as cutting ground-dwelling animals such as bandicoots out of hiding places. McBryde (1977) linked the representation of ground-edged axes (or fragments) at GB4 with increasing possum consumption through time. Between contexts bounded by dates of 3690–3251 to 2364–2070 calBP and 2364–2070 to −1836 calBP, ground-edged axes increase in abundance by 25 times (from 0.1% to 2.5% of all implements) concurrent to a possum MNI increases in abundance from 9.4% to 24%.
Conversely axes are most common on the surface deposit of GB4 (10.3%) where possum also dominates (72.5%) but there is very little animal bone of any kind on the surface, giving an exaggerated impression of their importance. The resulting question of whether the apparent association of possum exploitation and ground-edged axe usage continued in Graman after ~2091–1836 calBP is answered by the more recent dates obtained from GB1. At GB1, the only notable increase in arboreal marsupial abundance and their dietary importance occurs in Phase 5, which begins after 2000 calBP.
This peak in arboreal marsupial consumption at GB1 therefore seems to coincide with the peak of ground-edged axe deposition at GB4. It is also worth pointing out that bandicoot (Peramelidae) consumption experiences a resurgence in Phase 5, and that an increase in this family’s representation (from 2.1 to 9.5 and then 21.8%) is also observed at GB4 concurrent to the possum increase. Again we caution that there is no indication as to the specific period within the last 2000 years from which the surface materials of either site derive – hence, there is no particular confidence or indication that the stratified arboreal marsupials from GB1 are particularly closely linked to the GB4 surface axes, which might be considerably more recent. At face value the general premise that adoption of ground-edged axes facilitated and resulted in increased exploitation of arboreal marsupials (and perhaps bandicoots) is acceptable.
Changes in demography, social and economic organisation
A well-known continental model of Aboriginal demography generated using radiocarbon data and geospatial techniques (Williams et al., 2015) has made the following assertions about changes during the Holocene. Optimal conditions in the Early to Middle Holocene led to the rapid expansion of and growth of human populations, but the onset of ENSO (in this model dated c. 4500–2000BP) resulted in fragmentation of these populations, abandonment of economically marginal areas and a sharp contraction (26%) in average ranging territory. During this period it is posited that a broadening of the diet occurred, in combination with intensified ‘low-level food production and hunting strategies’. With the subsequent climate amelioration, people were able to re-colonise formerly marginal areas, and soon the packing of people into environments covering the entire continent resulted in the establishment of strong territorial boundaries (Williams et al., 2015). This model of change invokes both the climatic change as well as technological innovation covered in the previous sections.
Economic intensification and broadening of forager diets have long been thematic areas in Australian archaeology. ‘Intensification’ (Lourandos, 1983) has been used as term describing the economic adaptations to an internally driven increase in complexity of Aboriginal social organisation during the Late Holocene. Under this model, a broadening of the resource base beyond the largest macropod species – assumed to be the highest-ranked in preference amongst foragers – was necessary to sustain the nutritional demands of populations which were growing in size, density, and social complexity (with several flow-on effects, e.g. creating obligations to provide more food for larger ceremonial gatherings). Foragers under increased pressure to provide thus began to draw more regularly upon small ground-dwelling and arboreal marsupials, reptiles, birds and freshwater/marine resources, in addition to carbohydrate-rich seeds, thereby intensifying the extraction of resources within given occupational areas. This subsistence reorganisation was facilitated by a diversification of technologies and methods used to procure and process lower-ranked resources observed in various archaeological sites (Beaton, 1982; Lourandos, 1985; Morwood, 1986, 1987; Ross, 1985; Smith, 1986).
However, the same changes in the archaeological record have been interpreted alternatively (though not always in a mutually exclusive sense) as a response to foraging risk. The risk-reduction model described by Hiscock (1994) is a well-known example; it posits that Australian societies underwent considerable economic and technological reorganisation as a response to fluctuating climatic conditions, which generated instability in environments (including sea level rises and falls) and the resource bases they offered to foragers. People managed the risk of foraging failure in ‘unfamiliar landscapes’ by developing multifunctional, curated toolkits and modes of mobility capable of efficiently utilising resources as and when they were encountered. The risk-reduction model has found support in analyses of Late Holocene strategies of lithic reduction (Maloney and Hiscock, 2021; Veth et al., 2011) and plant processing (Asmussen and McInnes, 2013), and although explicit arguments from zooarchaeology are absent, it is implicit in this model that risk-reducing foragers cannot rely on a single taxon but must utilise a broader resource base (Hiscock, 1994; see also Winterhalder, 1986). A newer iteration of economic restructuring in response to resource depression emphasises that people resorted to intensified exploitation of resources offering lower nutrition at higher extractive labour costs, made more efficient through technology (Hiscock and Sterelny, 2023).
At GB1, the first (Middle Holocene) occupation phase already represents a broad and diverse diet, and in fact these measures only decrease through time when corrected for phase duration, reaching their lowest levels in Phases 3 through 5. Similarly, amounts of animal meat consumed per 100 years at GB1 appear to decline severely beginning in Phase 3, and do not ever seem to recover to the levels observed prior to ~4600 calBP. Therefore, we find little to no support for the notion that Aboriginal economic structure either became restructured around more intensive or broader-spectrum foraging patterns during the period generally associated with intensified ENSO conditions, at least as it pertains to animal foods. Although again these patterns may be heavily affected by the relocation of subsistence activities to GB4, it is difficult to entertain that this evidence signals any heightened human activity or increased population size within the local area.
The focussed hunting of two species of macropod and one species of arboreal marsupial observed within the last 2000 years, in Phase 5, is more interesting. This could plausibly be interpreted as an ‘intensified’ economic development, in so far as it describes specialised exploitation of these at the expense of others’ representation in the animal diet at GB1. It is important to note that one of the targeted species (O. robustus) is the second-largest by body mass local to the area and as such would have always been highly-ranked. However, the increasing importance of T. vulpecula, as a small-bodied taxon with a high extractive cost (requiring acquisition and maintenance of axes, laborious cutting of footholds or other means of accessing possums, and dangerous ascents into tree crowns), is certainly more aptly described as intensified reliance on an lower-ranked resource. The peak of freshwater mussel collection during this phase might also be considered a similar case, though without the same extractive costs.
In the context of social organisation, it interesting to consider that in Phase 5 the majority of animal foods appear to have come from the immediate hilly, woodland environs of Graman, with apparently reduced use of the flatter plains areas and grassy open woodlands beyond. In addition to generally signalling a tightening of foraging focus on the resources of a specific habitat type from an angle of economic specialisation, another potential interpretation of this pattern is that emergent territorial boundaries established by or during Phase 5 had restricted access to such regions from within. However, people at Graman clearly remained connected with the broader landscape because the raw material for axes used at these sites, metamorphosed boles, came from a hilly outcrop more than 15 km away (Boot, 1990). As such the inhabitants of Graman were either travelling to external localities across the flat plains, or were in contact with people able to access said stone resources.
Conclusion
Observed trends in animal consumption at GB1 do not suggest a broadened, diverse diet incorporating substantially larger proportions of smaller animals over macropod hunting developing during the Holocene, as has been suggested by previous interpretations of other sites across Australia, including nearby GB4. Instead, a series of changes in animal subsistence occur in various directions at different times, but with an underlying and constant reliance (by weight of meat) on macropods for the vast majority of game throughout the site’s occupation. The final phase of occupation, corresponding to the last 2000 years, saw increased specialisation of hunting one of each specific large, medium and small-sized marsupial taxa within the site’s local environmental setting. As such, GB1 joins a growing list of Australian archaeological sites that do not cleanly conform to predictions of ongoing development of generalised greater dietary breadth and/or increased smaller-game utilisation during the Holocene.
We find that palaeoenvironmental data for Graman support the notion that local environments continued to sustain a stable supply of medium and large-sized macropods throughout the Holocene. Our results also suggest a chronological association of increased macropod consumption with backed artefact deposition and, although more tentatively, increased arboreal marsupial consumption with ground-edged axe deposition. These results suggest causal relationships between novel Holocene extractive technologies and exploitation of specific marsupial taxa, although the strength of this argument is constrained by relatively coarse radiocarbon resolution, particularly for the last 2000 years. The role of climate change and the effects of dingoes as competing predators conversely remain dubious, as over the period of time in they were active in the area they do not appear to have impacted macropod availability to any notable degree.
Finally, the tightened focus of hunting around O. robustus, P. penicillata and T. vulpecula in the last 2000 years is suggestive of intensified exploitation of these resources within the immediate vicinity of Graman, although we find no evidence of predicted increased dietary breadth or intensified general economic activity prior to this time. Further study of additional faunal assemblages spanning the Middle to Late Holocene of eastern Australia, as well as reassessment of the GB4 fauna, will help elucidate the timing, direction and factors involved in subsistence change amongst the Aboriginal population of this region.
Supplemental Material
sj-docx-1-hol-10.1177_09596836251340811 – Supplemental material for Holocene subsistence change in eastern Australia: The zooarchaeology of Graman B1 Rockshelter
Supplemental material, sj-docx-1-hol-10.1177_09596836251340811 for Holocene subsistence change in eastern Australia: The zooarchaeology of Graman B1 Rockshelter by Loukas George Koungoulos, Jane Balme and Sue O’Connor in The Holocene
Footnotes
Acknowledgements
We are grateful to the Ashford Local Aboriginal Aboriginal Land Council for permission to conduct the study, and for the assistance of Mariko Smith, Rebecca Jones and Niamh Formosa in facilitating access to the Graman assemblage. We also give thanks to Sandy Ingleby, Collections Manager of Mammalogy at the Australian Museum for facilitating access to comparative materials for identification and to Tom Peachey for performing the microCT scan.
Author contribution(s)
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This study was funded by an Australian Government Discovery Project Grant (DP210101960).
Ethical approval and informed consent statement
Destructive analysis of bone for AMS radiocarbon dating was performed under authorisation of Heritage NSW, Department of Planning and Heritage (AHIP 5054) and the Australian Museum Trustees, with approval and permission of the Ashforld Local Aboriginal Land Council for the study.
Data availability statement
All data discussed in the study are available in the main text or supplementary information.
Supplemental material
Supplemental material for this article is available online.
Notes
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.