
Abstract
During the Holocene, Iberian ecosystems have changed dramatically in response to different biotic and abiotic drivers. Typically, the impact of these changes on ecosystems is assessed in terms of the variation of diversity in the communities that compose them. However, there are a wide variety of population-specific parameters of vital importance for the maintenance of the structure and health of these ecosystems. In this study, we have focused on the drastic decrease in body size (body mass) during the Holocene in three large mammal species in the Cantabrian Mountains. Namely, the brown bear (Ursus arctos), the chamois (Rupicapra pyrenaica parva) and the wild horse (Equus ferus ferus). The research combines the analysis of new findings with an exhaustive compilation of previously published data to assess changes in the body size of these species at different geographical (European, Iberian and Cantabrian Regions) and chronological scales (during the Quaternary – i.e. Pleistocene and Holocene). We aim at assessing the impact of anthropogenic and climatic factors on macromammal populations in the Iberian Peninsula, using new data obtained from the new archaeo-palaeontological site of Cueva de Llamazares (León, Spain). The results show that, although the final decline in body size during the Holocene occurred in the three species, each of them seems to have been driven by different factors. Bear populations seem more influenced by their mobility versus the potential impact of changing in human hunting dynamics, while changes in vegetation cover would have been most important for chamois, and the gradual effect of climate changes for horse populations. Research covering broad chronological ranges in the past helps understanding the present and future dynamics of these species, thus contributing to their conservation and management.
Introduction
The Iberian Peninsula is home to a variety of native species and subspecies of iconic large mammals of key ecological and socio-cultural importance (Pascual et al., 2011). These include the Iberian lynx (Lynx pardinus), the Iberian wolf (Canis lupus signatus), the brown bear (Ursus arctos) and the Cantabrian chamois (Rupicapra pyrenaica parva), among others. Nevertheless, they are just a few of the species that inhabited Iberian ecosystems over the last thousands of years (Crees et al., 2016; Lorenzen et al., 2011; Puzachenko and Markova, 2019; Turvey, 2009). Many direct and indirect anthropogenic or purely climatic causes have been considered as the origin of an extreme loss of diversity of large wild mammals (e.g., extinction of wild horses, Equus ferus ferus; Bendrey, 2012; Sommer et al., 2011), genetic impoverishment (e.g., for bears; Valdiosera et al., 2008), and niche displacement in the populations that remained (Sillero et al., 2022; Wilson et al., 2020). Drastic changes in the distribution of some species (e.g., Iberian lynx; Clavero and Delibes, 2013; Gil-Sánchez and McCain, 2011), have also occurred, due to habitat loss and fragmentation and human persecution.
Specifically, the Cantabrian Mountains, located in the north of the Iberian Peninsula, have played a crucial role as a climatic refuge and source of endemisms over the last thousands of years (Abellán and Svenning, 2014; Macaluso et al., 2023; Sommer and Nadachowski, 2006). Precisely, its richness, diversity and indices of endemisms are among the highest in the Iberian Peninsula, especially for mammals, giving this region the consideration of a hotspot (Pascual et al., 2011). Among its fauna, we still find populations of large carnivores, such as the brown bear (Penteriani et al., 2020) and the Iberian wolf (Ordiz et al., 2022).
Brown bears display great ecological range, with historical populations ranging from Mexico to Siberia, that is, occupying most of the northern Hemisphere, down to North Africa (Davison et al., 2011). Nowadays, bears are in risk of extinction in the Cantabrian Mountains (Penteriani et al., 2020). Although the population has shown some recovery over the last two decades, effects of climate change on crucial bear food items could lead to a severe bear population decline in the next 50 years (Penteriani et al., 2019). It has been hypothesised that Cantabrian bears are the result of isolation in glacial refuges during the Last Glacial Maximum (LGM), forming the theoretical ‘Iberian Refuge’ (Davison et al., 2011). Other proposals suggest a possible rise of bear populations from the central and southern Iberian Peninsula at the end of the LGM (García-Vázquez, 2015). Subsequently, several bottleneck phenomena have greatly conditioned the genetic diversity of the current Iberian population (Valdiosera et al., 2008). In any case, both genetically and phenotypically, the history of this bear population poses great unknowns that seem strongly conditioned by the effect of changes in climate (García-Vázquez et al., 2015) and a potential human-animal conflict that is resurfacing in recent years (Krofel et al., 2020).
On the other hand, less data are available on the geographic distribution patterns and population changes of Cantabrian chamois (R. p. parva; García-González and Herrero Cortés, 2007) during the Quaternary. These small bovids are included in a genus currently distributed in several of the main European mountain ranges, forming a mosaic of metapopulations that reflect a complex biological history of population movements and isolations (Masini and Lovari, 1988). In particular, the current Cantabrian subspecies is phylogenetically closer to the Pyrenean chamois (R. p. pyrenaica) and the Appenine chamois (R. p. ornata) than to the populations distributed in the Alps (R. rupicapra; Lovari, 1987). Still, the Cantabrian population presents a series of peculiarities that differentiate it from their neighbours in the Pyrenees, both anatomically and ethologically, suggesting a relatively ancient separation of the populations (Lovari and Scala, 1980). Interestingly, this population context becomes more complicated when we include palaeontological data in the equation, highlighting different characteristics in the Pleistocene chamois populations of the Cantabrian Mountains compared to the present-day ones, mostly in terms of individual body size (body mass; Arceredillo et al., 2011; Arceredillo, 2015; Rodríguez-Gómez et al., 2022). The susceptibility of this species to geographic isolation due to climatic effects (Domínguez Sanjurjo et al., 2009; Pérez-Barbería and Palacios, 2009), together with its status as the main source of meat for human populations that inhabited alpine environments prior to sedentarisation (Rodríguez-Gómez et al., 2022), and the subsequent start of trophy hunting (Pérez-Barbería and Robles, 2009), are factors to evaluate when trying to understand the current peculiarities of chamois populations in the Cantabrian Mountains.
One of the extreme cases of loss of wild populations leading to regional extinction during the Holocene has been that of the wild horse (E. f. ferus). In the past, this large herbivore was distributed over virtually the entire northern hemisphere, being mainly associated with large open spaces (from tundra-steppes to great plains from Alaska to the Iberian Peninsula; Bennett and Hoffmann, 1999). This species has been recorded in the peninsula since the Middle Pleistocene (Maldonado, 1997), with populations widely distributed throughout the territory. This situation possibly continued until the Bronze Age (Sommer et al., 2011), although historical records of hunting in wild populations are preserved until second century ACE (Hoyo, 2002; Montaner, 2001). Unfortunately, little is known about the processes that led to the extinction of this species throughout the peninsular territory (Bernáldez-Sánchez et al., 2024). In this regard, it is worth highlighting the theoretical importance of the human impact due to hunting and provisioning (Carbajo, 2020; Galindo-Pellicena et al., 2017) and the impact of the synchronous beginning of the domestication of these animals with the extinction of wild populations (Gardeisen, 2005), with the Iberian Peninsula possibly acting as one of the nuclei of this process (Warmuth et al., 2011).
Surprisingly, these three species display a common biological phenomenon: a large decrease in the size of the individuals of their recent Iberian populations with respect to the populations of the Late Pleistocene or Early Holocene. This phenomenon has been documented in Alaska (USA) in the last populations of wild horses before their extinction (Dale Guthrie, 2003), in European lions (Marciszak and Gornig, 2024) and in Italian wild boars (Iannucci et al., 2020). More difficult to decipher are the potential causes that have led to this situation. In some cases, climate change (Gardner et al., 2011) or resource availability (McNutt and Gusset, 2012) have been hypothesised as major factors for the decline in size (Iannucci et al., 2020). In turn, the human effects seem to have played an important role in this phenomenon through indirect alterations of the above parameters or direct pressure on mammal population density and structure (Mason et al., 2014). To understand this situation in detail, it is first necessary to determine when the decline in size of the individuals of these populations began and the trends of change with which the decline occurred, which would allow us to infer the ecological pressures present in the ecosystems in these chronologies.
This paper presents novel data on the biological characteristics of the populations of bears, chamois and wild horses in the Cantabrian Mountains during the Holocene, obtained from individuals preserved in the recently studied archaeo-palaeontological site of Cueva de Llamazares (León province, northern Spain). Radiocarbon dating and a detailed study of the individuals presented here have revealed certain phenotypic peculiarities of the populations of these Cantabrian mammals with respect to the known data on their current populations. These findings, together with an exhaustive compilation of data previously provided in other scientific publications, have allowed us to evaluate the changes in body size of the three species at different geographical (European, Iberian and Cantabrian) and chronological scales (during the Pleistocene and Holocene). The main objective of this analysis is thus to assess the impact over time of anthropogenic and climatic factors on large mammal populations in the Iberian Peninsula, highlighting the potential of studies covering wide chronological ranges in the past for the correct modelling of the present and future status of the studied species.
Material and methods
Study site: Cueva de Llamazares
Cueva de Llamazares (42°58′N, 5°26′W) is a karst cavity of 776 m in length and 51 m in vertical range, located at 7 km from the water divide of the Cantabrian Mountains (Supplemental Figure 1a, available online). The cave entrance is at 1460 m altitude and 230 m from the base of the mountain where it is located (Supplemental Figure 1b and c, available online). The cave is a regional geosite used as a show cave, and as a result, the original entrance and the eastern gallery have been modified, likely impacting the palaeontological site. The two main cave galleries respond to the karstification of Carboniferous limestone and fluvial incision occurred during the Quaternary (Supplemental Figure 1d). At a climatic level, the region currently experiences a cold climate with dry and cool summers in the mountainous areas and a temperate climate with dry, mild summers in the valleys, corresponding to Köppen-Geiger classifications Dsc and Csb, respectively (Beck et al., 2018). The average temperature is around 7°C, with annual precipitation reaching approximately 1040 mm, according to climatic data from the Spanish Meteorological Agency (www.aemet.es).
In the area closest to the current entrance to the cave there is a lower level of small passages with some debris deposits resulting from breakdown processes over the years (Supplemental Figure 1e, available online). It is in this area where the skeletal specimens analysed in this study were found (Supplemental Figure 2a, available online). Together with these specimens found on the surface, it has been possible to locate stratigraphic levels of greater or lesser strength that contain archaeological materials that will be the subject of future research. Despite the occasional mention in popular publications (Fernández-Martínez et al., 2021; Fidalgo et al., 2022), this work presents the faunal site of Cueva de Llamazares to the scientific community for the first time.
Material
The analyses presented here include four partial skeletons (and a specimen of a fifth individual), hitherto unpublished, found at the site of Cueva de Llamazares: three brown bears (Ursus arctos; we will call them ‘bear 1’ (Supplemental Figure 2f and h, available online) and ‘bear 2’ (Supplemental Figure 2f and h, available online); and a pisiform of a third individual, ‘bear 3’ (Supplemental Figure 3h, available online)), a chamois (Rupicapra pyrenaica; Supplemental Figure 2i, available online) and a horse (Equus ferus; Supplemental Figure 2g, available online). The spatial distribution of the individuals in the site can be seen in Supplemental Figure 2a, available online. All specimens were collected on the surface (as can be seen in Supplemental Figure 2b–e, available online), together with information on their spatial arrangement in the cave. All materials are temporarily preserved in the Palaeontology Area of the University of León. The 3D models of the specimens used in the analyses of this work are available in the Sketchfab platform (https://skfb.ly/p8pzS).
In order to approach the changes in the biometric characteristics of the individuals of the brown bear, chamois and horse populations, data have been compiled from a large number of bibliographic sources, as explained in the Supplemental Tables 1–3. In the case of bears, we include data from individuals from sites in the Cantabrian Mountains dated to different Holocene chronologies, together with brown bear data from current populations in this mountain range. The chamois data correspond to present-day individuals from populations of the different European subspecies proposed and fossil specimens from the Middle and Late Pleistocene and Holocene of Italy, France and the Iberian Peninsula. For horses, information from European fossils from the Middle and Late Pleistocene of Equus ferus and Equus hydruntinus populations, wild individuals from Holocene sites, domestic horses from historical sites and present-day domestic and wild individuals (Equus przewalskii) have been used.
Dating
The four partial skeletons found in Cueva de Llamazares have been dated using radiocarbon techniques (Calibrated Radiocarbon Age) on bone (Supplemental Figure 2j, available online). The bear individuals were dated using the services of Beta Analytic Testing Laboratory, while the chamois and horse were dated using the services of Vilnius Radiocarbon, Center for Physical Sciences and Technology. The dated specimens correspond to LLAMA_141, LLAMA_433, LLAMA_461 and LLAMA_188.
The chronologies used for the bibliographical data are taken from the studies from which the biometric information was obtained. The specific data for each specimen can be found in the section ‘Characterisation of the individuals from Cueva de Llamazares’. In the case of bears, the chronologies represent the results of calibrated radiocarbon analyses and are limited to the Holocene. Due to the lack of absolute chronological data for the majority of chamois and horse individuals, broader time classes have been used, assigning individuals to the Middle Pleistocene, Late Pleistocene, Holocene, Historic and present day.
Taphonomic analysis
Taphonomic analysis of the individuals shows no evidence of anatomical bias in the accumulations. The individuals have a representation of anatomical elements from a large part of the skeleton, including small sesamoids, although the lack of complete skulls and mandibles due to the subsequent looting and deterioration of the site is noteworthy. We do not rule out the loss of materials due to vandalism, as it has been a cave locally known for ages. There is no evidence of human exploitation of the carcasses, with a total absence of cuts, bites, intentional breakage or burning processes. We only found some old post- or peri-mortem fractures in the case of the bears, covered by a thin layer of carbonate, although they are far from intentional fractures and it is difficult to approximate the exact time of fracturing. In all cases, there are a large number of present-day fractures, possibly derived from trampling in recent times.
No marks of carnivore exploitation of the carcasses were found, except for a few bite marks on the proximal and distal ends of two ribs of bear 1. Also on bear 1, we found some marks of access by rodents on the sharp edges of some ribs, naviculars and metatarsals. No other macroscopic biological activity marks, such as insects, roots and fungi, have been found. Likewise, the absence of marks of physical processes of mobilisation of the bony elements, such as drag marks or rounding of the edges, is also evident. On the contrary, in all cases there is a large amount of evidence of chemical processes of bone alteration, potentially caused by having been submerged in small bodies of water for long periods of time. Finally, almost all the specimens are partially or completely covered by a more or less thin layer of naturally deposited carbonate, depending on the activity observed in this cave.
Because of this, it is inferred that the horse, chamois and bears (with the exception of bear 3, represented by a pisiform) entered the deposit site in one piece. In general, these individuals must have remained at or near the original deposit site from the time the carcass was brought in. These carcasses must have remained largely isolated, avoiding access by humans, carnivores, rodents and other biological disturbance factors throughout the decomposition process. At some point between deposition and extraction, the specimens had to remain for long periods of time partially submerged in water and exposed to the gradual surface deposition of carbonates on them. Finally, the loss of information due to selective spoliation and fragmentation by continuous trampling seem to be evident.
The faunal assemblage accumulated in Cueva de Llamazares through various mechanisms depending on the taxon. In the Cantabrian Mountains, the brown bear is a species that utilises frequently karst caves as shelter, including for hibernation (e.g., Grandal-d’Anglade et al., 2019). Therefore, the ursids likely ventured approximately 20 m into the interior of the Cueva de Llamazares in search of refuge. Large herbivores typically do not voluntarily enter caves, so their presence in palaeontological cave deposits is generally attributed to accidental falls or being brought in by carnivores or hominins (e.g., Álvarez-Lao, 2014; Álvarez-Lao and Méndez, 2016). However, certain caprines (e.g., Capra pyrenaica) that inhabit rocky areas use cave entrances as shelter. This is the case with the Cantabrian chamois, whose fossil remains were found just 5 m from the current entrance of Cueva de Llamazares. Unfortunately, the entry of the wild horse remains uncertain. The horses inhabit open herbaceous areas (Sommer et al., 2011) and do not typically use caves as refuge, making their presence in or around Cueva de Llamazares unexpected. This suggests that humans or carnivores may have introduced the horse, although the lack of evidence for tooth or cut marks on the bones prevents this interpretation from being definitively confirmed.
Phenotypic analysis
To facilitate the analysis of the materials from Cueva de Llamazares, all the specimens were scanned in three dimensions. The scans were carried out using the Creality Lizard device (Accuracy 0.05 mm; Resolution 0.1–0.2 mm) and edited in the CR Studio software. All biometric data of the specimens were measured on the 3D models with the use of MeshLab software.
The proximal epiphysis of the humerus specimen from bear 2 (Supplemental Figure 3d, available online) has been virtually reconstructed by simulating this segment on the opposite side of the same individual, superimposing both 3D models and merging them. CR Studio, Blender and GeoMagic software have been used in this process.
The taxonomic, ontogenetic, sexual and population determination of the unpublished materials has been carried out by combining qualitative anatomical study and quantitative analyses with linear biometric data. The characterisation of the changes in the populations of these mammals over time has been carried out through the analysis of linear biometric data and the estimation of the total body size of the individuals:
Brown bear: the age of the individuals was estimated by analysis of the fusion of the epiphyses of the long bones and vertebral discs, and of the dental wear in the lower (m1) and upper (M1) first molars. For the biometric analyses we used linear measurements of m1 and M1 and body size estimations with humerus and radius measurements (according to the equations of Christiansen, 1999 and Viranta, 1994) from sites in the Cantabrian Mountains dated to different Holocene chronologies. Together with these data, we have included the known ranges of body size of female and male brown bears from current populations in the Cantabrian Mountains.
Chamois: the estimation of the age of the individual has been carried out through the analysis of skeletal fusion and dental wear in the mandibular dental series. For the biometric analyses, we used linear measurements of bone cores of current individuals from populations of the different European subspecies proposed, with special emphasis on data from both sexes in Rupicapra rupicapra and Rupicapra pyrenaica. On the other hand, measurements of bone cores from Middle and Late Pleistocene and Holocene individuals from Italy, France and the Iberian Peninsula have been used. Independently, body size estimates have been included with linear measurements of humeri and radii (according to equations of Scott, 1990, as in the work of Rodríguez-Gómez et al., 2022) of Middle and Late Pleistocene and Holocene individuals (including those from current populations) from the Iberian Peninsula. Together with these data, the body size ranges directly observed in the populations of females and males of the present-day populations of the Iberian Peninsula have been included.
Horse: the age of the individual was estimated by skeletal fusion and dental wear analysis. For the biometric analyses, linear metacarpal measurements (MC III) of European Middle and Late Pleistocene fossils of Equus ferus and Equus hydruntinus populations, wild individuals from Holocene sites, domestic horses from historical sites and present-day domestic and wild individuals (Equus przewalskii) have been used. Body size estimates of the above classes calculated from different limb bone dimensions have been used independently (according to equations of Alberdi et al., 1995 and Scott, 1990 or the mean of both estimates; for more information see Supplemental Table 3, available online).
In the case of the temporal trend in body size of chamois and horses in the Iberian Peninsula, the data were sufficient for statistical tests. Due to the small sample size and the non-normality of the data, the non-parametric Kruskal-Wallis test with its associated Mann-Whitney post-hoc test was used. Analyses were performed in Past 4.03 software (Hammer et al., 2001).
Characterisation of the individuals from Cueva de Llamazares
This work has been able to characterise the presence of three bear’s individuals at the site. Individual 1 (bear 1) was located right at the entrance to the lower galleries, with some specimens deposited by gravity in the first of the lower areas, overlapping the specimens of the horse individual (Supplemental Figures 2 and 3). The absolute fusion of all the epiphyses of the long bones and vertebral discs of this individual, together with the presence of small areas of exostosis in the metatarsals, allow us to assert that it was of adult age (over 9 years old; Weinstock, 2009), although the moderate wear of its m1 distances us from the image of a senile individual (Supplemental Figure 3e, available online). Direct radiocarbon dating of this individual gives a chronology of 8724–8546 cal a BP (calibrated; Supplemental Figure 2j, available online). Individual 2 (bear 2) was more dispersed in the lower cave passages, with some specimens close to one of the entrances to this area, a large part of the skeleton above a small cave section and the rest of the specimens recovered inside the passage (Supplemental Figures 2 and 3, available online). Following Weinstock’s (2009) proposed long bone epiphysis fusion series, an approximate age of 6–7 years is estimated for this individual, taking into account the fusion of the olecranon of the ulna together with the lack of fusion in the proximal epiphysis of the humerus. The absolute chronology of this individual corresponds to 8025–7939 cal a BP (calibrated; Supplemental Figure 2j, available online). The third individual (bear 3) is only represented by a small pisiform found in the first of the galleries of the visitable wing (Supplemental Figure 3h, available online). In all three cases, the anatomical characteristics of all the specimens are typical of Ursus arctos and the dating rules out the presence of individuals belonging to other species of bear on the Iberian Peninsula (Torres, 1984). The finding of the baculum of bear 1 (Supplemental Figure 3c, available online), together with the analysis of the proportions of m1 (Supplemental Figure 3e and Figure 1a, available online) confirm its assignment to a male. Analysis of M1 from bear 2 also points to the male sex of this individual (Supplemental Figure 3f and Figure 1b, available online).
The partially preserved chamois individual was found completely accumulated in the small pit located in the gallery where bear 2 was located (Supplemental Figures 2 and 4, available online). A hemimandible, the base of the bone core with part of the skull, several elements of the appendicular skeleton and a large part of the axial skeleton were recovered (Supplemental Figure 2i, available online). The dental eruption with the presence of the third lower molar and the degree of dental wear inform us that this is a fully adult individual, but without reaching senile age (Supplemental Figure 4d, available online). The absolute chronology obtained from the individual corresponds to 5326–5266 cal a BP (calibrated; Supplemental Figure 2j, available online). The comparison of the dimensions of the bone core highlights more biometric similarities of the Llamazares individual with the current chamois of the Alps (Rupicapra rupicapra rupicapra) or of the Iberian Peninsula (Rupicapra rupicapra pyrenaica), being outside the range of variability of the rest of the Rupicapra subspecies (Figure 2a). If we include sexual dimorphism in the analysis, the specimen studied would be closer biometrically to what is expected in the males of the Iberian Peninsula chamois or in the females of the Alps (Figure 2b). If we consider the measurements of individuals of this geographic range at different times of the Pleistocene and Holocene we can see a great homogeneity between French, northern Italian and Iberian Peninsula individuals in the Late Pleistocene and Holocene. The Llamazares individuals looks especially close to Holocene individuals from France and the Iberian Peninsula and to the average of the Upper Pleistocene individuals from the Grotta di Equi (Italy; Figure 2c).
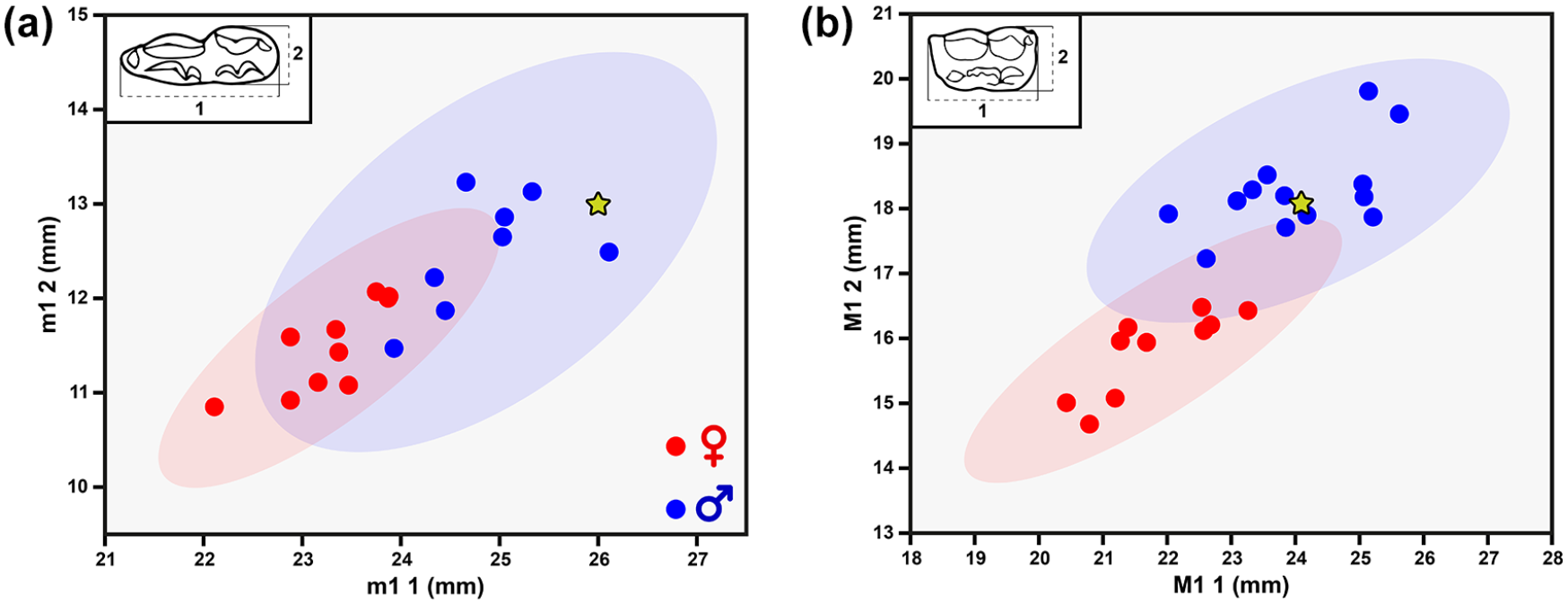
Bivariate plots of lower ((a); see Supplemental Figure 3e, available online) and upper ((b); see Supplemental Figure 3f, available online) first molar measurements indicating sex estimates for each individual. The stars mark the specimens from Cueva de Llamazares. The original data can be seen in the Supplemental Table 1, available online.
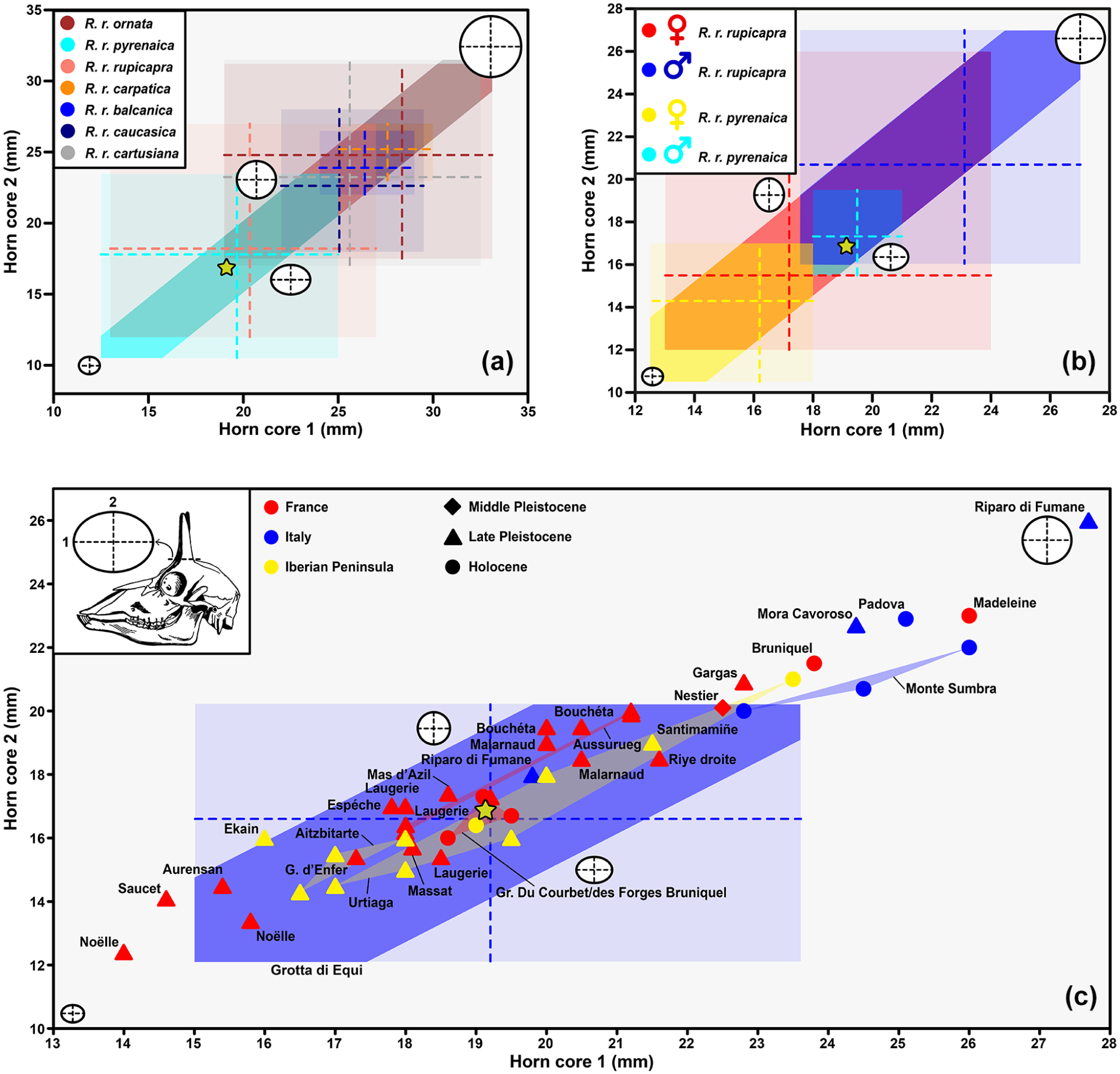
Representation of biometric analyses of chamois horn. (a) horn base proportions of the different subspecies (or species depending on which author) of chamois, (b) horn base proportions of the two sexes in the two subspecies (or species depending on which author) of chamois inhabiting the Iberian Peninsula today, (c) horn base proportions of Pleistocene and Holocene individuals of chamois found in the Iberian Peninsula, Italy and France. The stars mark the specimens from Cueva de Llamazares. The original data can be seen in the Supplemental Table 2, available online.
The partial equid skeleton located in the lower part of the wash where bear 1 was found preserves almost all of its anatomical elements, although the skull and mandible have suffered a great deal of fragmentation (Supplemental Figures 2 and 5, available online). The dentition of this individual is definitive, although with a low degree of dental wear (Supplemental Figures 5a–i, available online). This characteristic together with the absence of fusion of the vertebral discs in some thoracic vertebrae informs us that it would correspond to a young adult. The presence of pli-caballin and the shape of the ectoflexid confirm the attribution of the individual to the species Equus ferus (Supplemental Figure 5, available online). The large proportion of the lower canine in relation to the size of the skull and the mandible point to a male (Supplemental Figure 5k and l, available online). It is more difficult to ascertain the attribution of the individual to a completely wild population or to a domestic population. The absolute chronology obtained for the specimen places it at 2056–1882 cal a BP (calibrated; Supplemental Figure 2j, available online). The biometric analysis of the metacarpals highlights affinities of the Llamazares individual with wild horse populations of the Late Pleistocene and Holocene, and with the first phases of European domestications (Figure 3). It should be noted that there is no evidence of domestic use of the animal, no stress marks in the axial and appendicular skeleton, no differential wear in the dentition due to the use of oral appliances and no other pathology associated with anthropic exploitation.
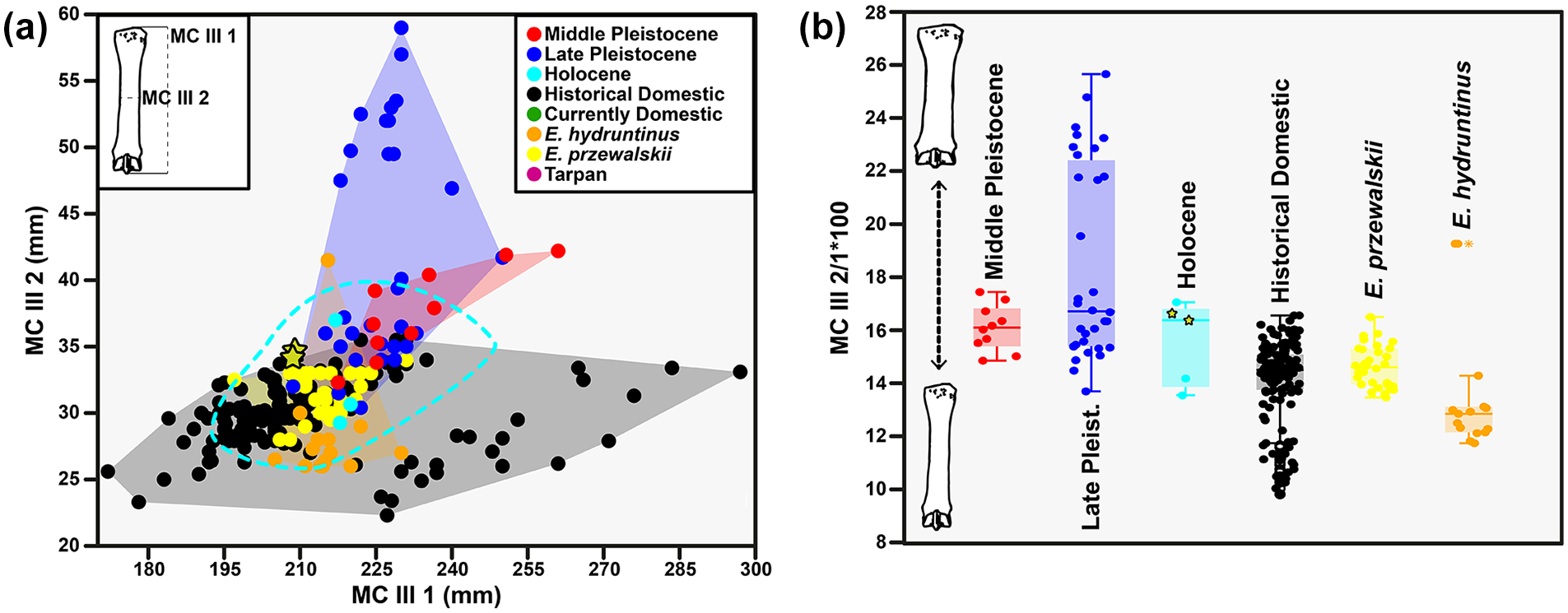
Representation of the biometric analyses of horse metacarpals. (a) bivariate analysis of linear measurements and (b) robustness indices of horse metacarpals distinguishing chronological classes, domestication phases and current equid populations with potential for the assignment and study of Cueva de Llamazares individuals. The stars mark the specimens from Cueva de Llamazares. The original data can be seen in the Supplemental Table 3, available online.
Results
The analysis of the body size estimates of the known bear individuals from the Cantabrian Mountains throughout the Holocene shows that bears 1 and 2 from Cueva de Llamazares are larger than those estimated in the current individuals of the Cantabrian populations, reaching or exceeding 250 kg compared to a maximum of 163–200 kg for a current male (Figure 4). In fact, this phenomenon is observed in all individuals recorded up to ca. 7000 cal a BP, although it is true that there is limited information on absolutely dated individuals from that time to the present. Even so, the presence of individuals of very small size (close to 40 kg) ca. 4800 cal a BP at the Sima de Somiedo site, similar to the lower range observed in present-day female Cantabrian bears, marks a clear differentiation with what has been observed in previous chronologies. This phenomenon of size variation seems to have occurred rapidly, as demonstrated by the discovery of one of the largest Holocene individuals from the Cantabrian Mountains at the site of La Braña (ca. 7840-7660 cal a BP), weighing approximately 335 kg, with a chronology three millennia earlier than the much smaller individuals found in the Sima de Somiedo.
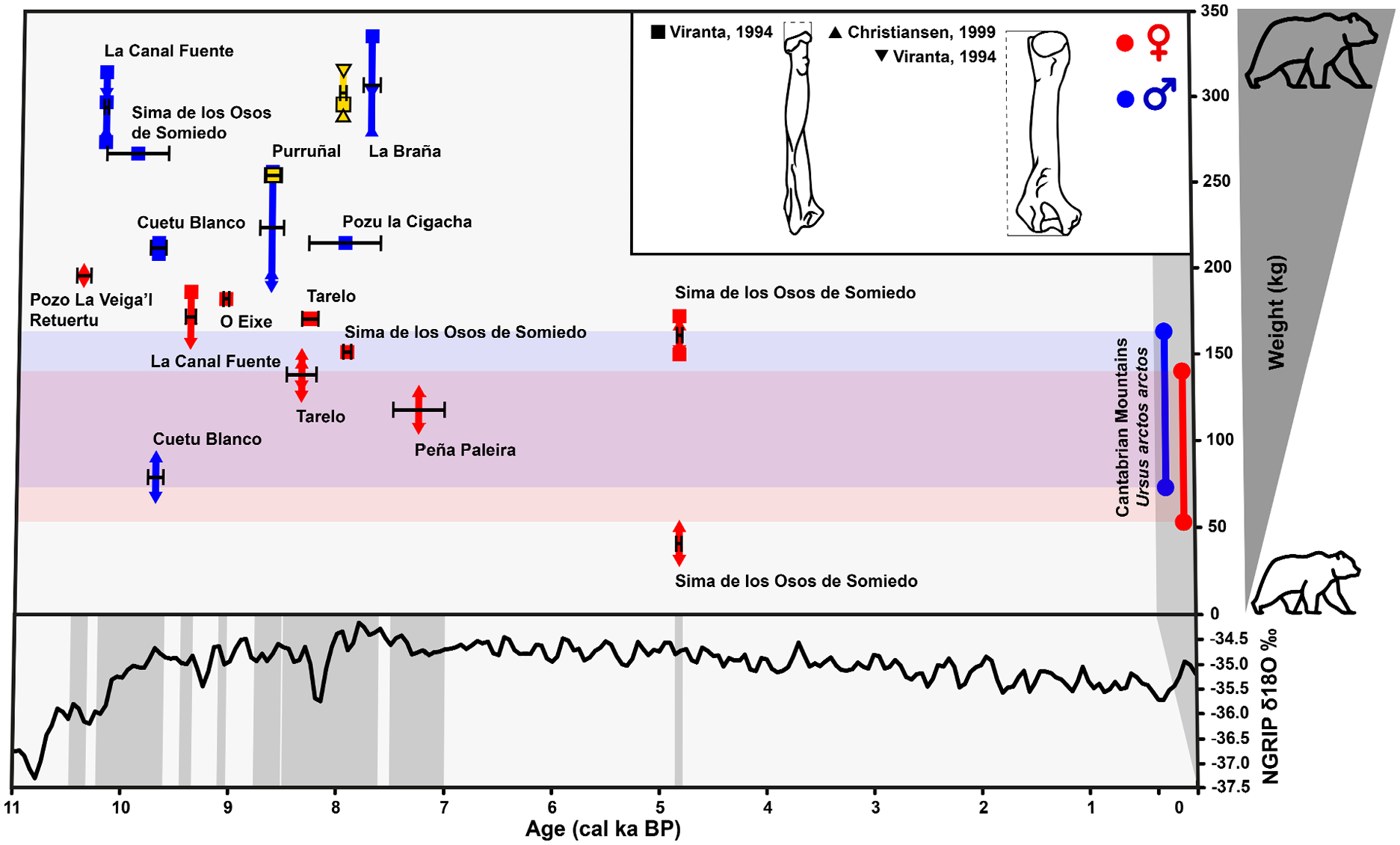
Representation of the estimated weight of brown bear (Ursus arctos) individuals found in Holocene sites in the Cantabrian Mountains together with weight estimates of present-day Cantabrian bears. The chronological range of each individual has been obtained from available radiocarbon dates. Weight estimation has been carried out with the biometric equations for radius and humerus of Viranta (1994) and Christiansen (1999). The original data can be seen in the Supplemental Table 1, available online. An estimate of climate changes inferred from the δ18O isotopic curve obtained from Andersen et al. (2004) is shown at the bottom of the graph.
The results of body size estimation of chamois individuals from the Cantabrian Mountains including the Basque Country through the Pleistocene and Holocene show a statistically significant trend of increasing size of individuals from the Middle Pleistocene to the Mid-Holocene (Figure 5). The dimensions of the Middle Pleistocene individuals resemble those expected in present-day individuals (with the sample from the Valdegoba site standing out), although with a greater range of variability. During the Late Pleistocene, a slight increase in size is observed, which seems to be quite homogeneous in almost all known localities. The contrast between the small size of the current individuals from the Cantabrian Mountains and Pyrenees compared to the maximums found at the beginning of the Holocene in the Basque area/country (Urtiaga cave) stands out. Including data from the Llamazares individual reveals the small size of the chamois in the Iberian Peninsula from the middle of the Holocene, in contrast to the data with slightly earlier chronologies.
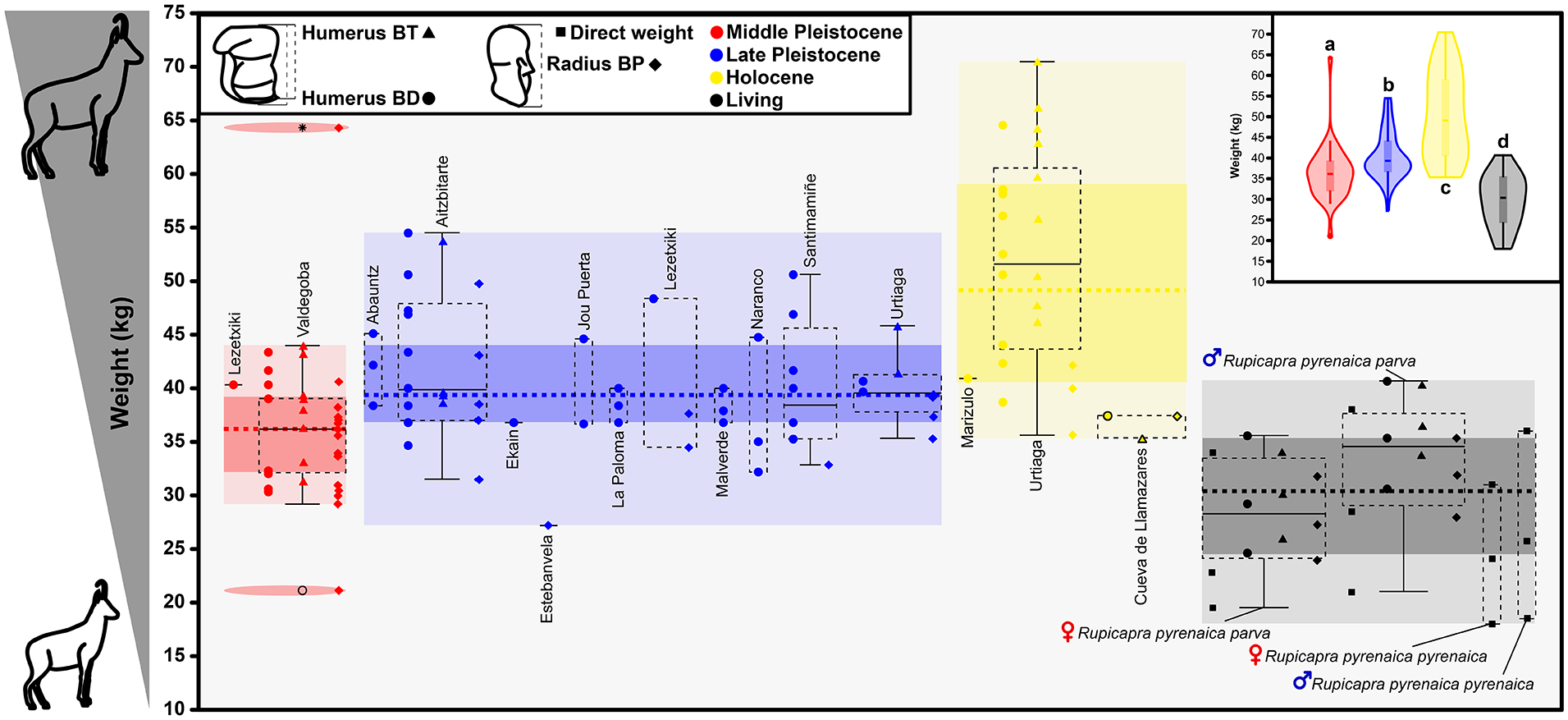
Representation of the estimated weight of chamois (Rupicapra spp.) individuals found in Iberian Peninsula sites from the Middle Pleistocene to the Holocene together with the weight ranges of present-day chamois from the Cantabrian and Pyrenees. The chronological ranges of each individual have been taken from the original publications. Body size estimations have been made using biometric data of humeri and radii following the equations of Scott (1990) as in Rodríguez-Gómez et al. (2022). The original data can be seen in the Supplemental Table 2, available online. The class groupings resulting from the statistical tests are represented as the letters of the superscripts of the violin plots.
The weight estimation data for horses in Europe and the Iberian Peninsula show a sharp decline in the body size of individuals until the eventual increase in morphotype diversity with domestication (Figure 6; Supplemental Figure 6, available online). In non-Iberian Europe this decline in body size appears to be gradual from the beginning of the Middle Pleistocene to the end of the Late Pleistocene, but becomes extremely acute in the transition to the Holocene (Supplemental Figure 6, available online). In the Iberian Peninsula there in no evident change in the Middle-Late Pleistocene, but the decrease in body size from the Late Pleistocene to the Holocene is significant (Figure 6). The apparent contrast in the body size of Iberian individuals compared to those of the rest of the European populations during the Middle and Late Pleistocene is noteworthy (Supplemental Figure 6, available online). However, this possible difference seems to disappear in the Holocene. In the Holocene, the average body size of wild horses in the Iberian Peninsula and the rest of Europe seems to conform to that known from the last tarpanes and przewalskii horses. There are insufficient data to observe the changes in body size of individuals from these populations within the Holocene, especially linked to the entry of domestic equids into Europe. However, the Llamazares individuals, as well as those from Casetón de la Era, Cerro de la Virgen and the other European localities, show a more homogeneous carving and proportions that could be in line with some of the earliest known domestic individuals in Europe (Figures 3 and 6).
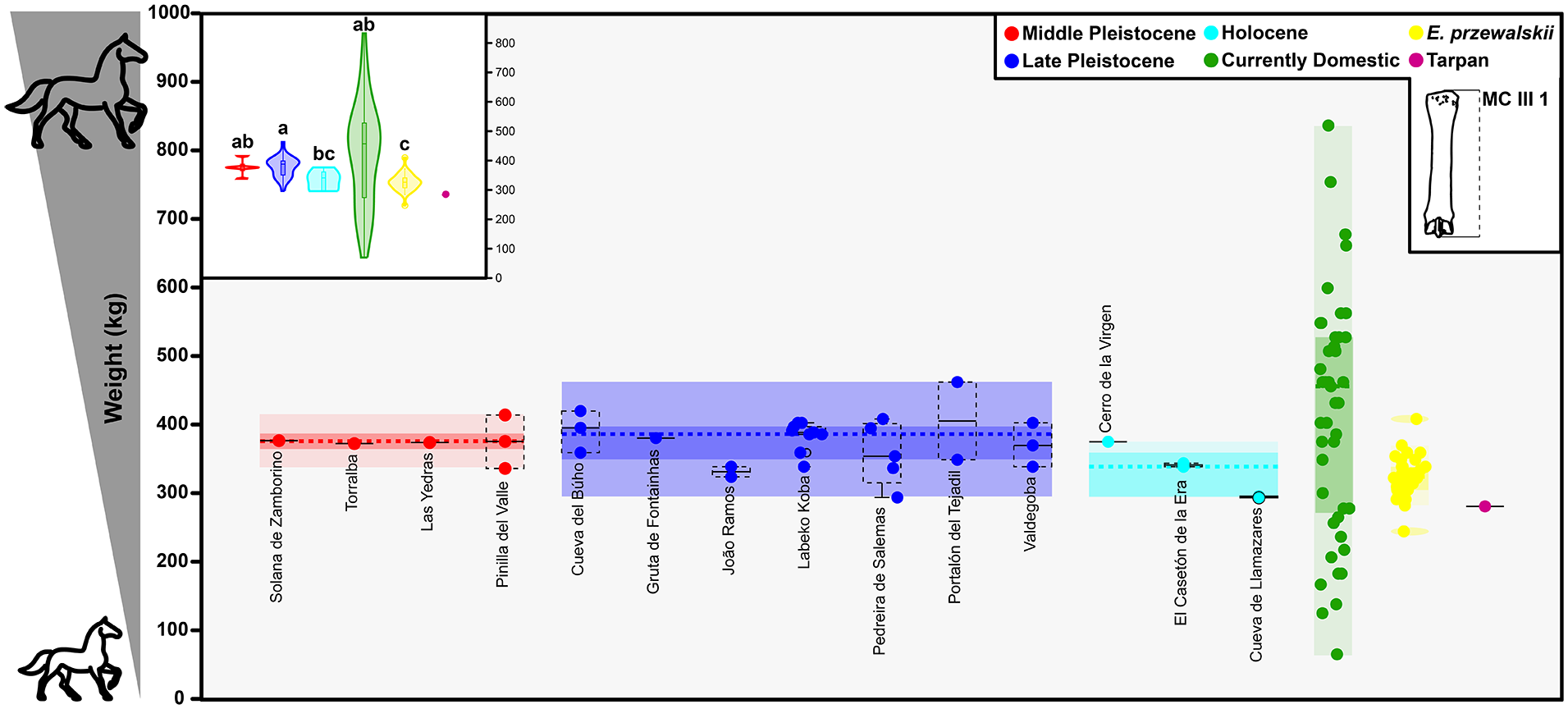
Representation of the estimated weight of horse (Equus ferus) individuals found in different parts of the Iberian Peninsula from the Middle Pleistocene to the Holocene together current weight variability of domestic horses and available data for przewalskii and tarpan horses. Chronological ranges are taken from the original publications or from the review by Saarinen et al. (2021). Weight estimates have been made from the mean of the estimation with the length of the third metacarpal, according to equations of Alberdi et al. (1995) and Scott (1990; for more information see Supplemental Table 3, available online). The class groupings resulting from the statistical tests are represented as the letters of the superscripts of the violin plots.
Discussion
The results of our study show a common pattern for the three species, with a decrease in individual body size to minimum values in the Mid and Late-Holocene (Figures 4–6). Nevertheless, the drivers of the observed pattern might be different, at least partially, for the three species.
The drastic reduction in body size of bears in the Mid-Holocene could be due to a population shift after a strong bottleneck in the previously settled populations (Valdiosera et al., 2008), or a migration from another population (García-Vázquez, 2015). The bottleneck would have implied a bias towards smaller individuals. According to the data of Valdiosera et al. (2008), two important bottlenecks were detected in Iberian bears during the Holocene, one about 10,000 years ago and another, a very recent one, during the last 350 years. Our results point to an onset of body size decline in Cantabrian individuals since at least ca. 5000 years ago, as evidenced for the bears found in the Sima de los Osos de Somiedo (García-Vázquez, 2015). However, data scarcity prevents a clear determination of the starting time of body size decline, which might rather be linked to the genetic bottleneck dated ca. 350 years ago (Valdiosera et al., 2008).
On the other hand, the hypothesis of migration from another bear population nucleus would require that the individuals of the source nuclei were smaller than those previously located in the Cantabrian Mountains. Based on molecular data, a displacement from the south of the Iberian Peninsula has been hypothesised, generating ‘refuge within refuge’ dynamics (García-Vázquez, 2015). The presence of bear populations in the central and southern Iberian Peninsula until, at least, the 16th century (Nores and Naves, 1993) may have allowed that dispersing process, particularly for a species with large home ranges and dispersing capacity (e.g., Støen et al., 2006). Body size reduction would also be expected for bears in the Pyrenees, the mountain range between France and Spain, during the Holocene, because Cantabrian and Pyrenean bear populations were connected until the XVII–XVIII centuries (Nores and Naves, 1993). Interestingly, despite the large morphological change observed in Cantabrian bears during the Holocene the results presented by García-Vázquez et al. (2018) do not seem to show large differences in their niche dietary parameters, an appreciation previously raised by García-Vázquez et al. (2015). A more extensive collection of case-specific data with absolute dating seems necessary to complete the biological history of Iberian bear populations, especially in chronologies after the Mid-Holocene, when their presence in archaeological sites becomes scarcer (García-Vázquez et al., 2015).
At the worldwide scale, changes in net primary productivity and winter temperature during the Holocene directly affected the population dynamics of the brown bear, by reducing the species’ reproductive rate (Albrecht et al., 2017). At the European scale, increasing human land use and winter temperature were the principal drivers of extinction, suggesting that increasing winter temperatures facilitated land use in regions with formerly too-harsh climates, thus expediting overhunting and habitat loss (Albrecht et al., 2017). Bears, as other large carnivores, have historically being persecuted by humans, but the length and intensity of persecution varied at trans-continental scale (Ordiz et al., 2011). Some particularly important findings for conservation and management of present-day bear populations is that bear mean litter size (i.e. reproductive rate) is positively linked with mean adult female body size and, strikingly, a long persecution history (Zedrosser et al., 2011). Observed trans-continental variation in body size and other important parameters in present-day bear populations are thus likely a response to human persecution (Zedrosser et al., 2011). Our findings for Iberian bears align with the patterns observed for other heavily persecuted bear populations in Europe, in both historical (Albrecht et al., 2017) and present times (Zedrosser et al., 2011).
As for bears, a decrease in body size of chamois would also be linked to great anthropic impact and the reduction of suitable habitat. The general trend shown in our results for Iberian chamois up to the Mid-Holocene seems to point to an increase in the average size of individuals with a widening of their size variation. This is far from a pattern of gradual decrease in size due to changes in climatic dynamics (Gardner et al., 2011). This phenomenon is especially recognisable in the comparison of chamois from the Late Pleistocene levels with those from the Early Holocene at Urtiaga (northeast Spain; Altuna, 1972). Subsequently, as the size estimated for the Llamazares individual is at the minimum values estimated for the chamois from the Early Holocene of Urtiaga (Altuna, 1972), between the Mid-Holocene and the present day, there has been a clear decrease in the average size of the chamois and its variability. As for the increase in anthropic pressure on chamois populations, chamois were already an important part of the diet of human populations from the Middle Pleistocene (Rodríguez-Gómez et al., 2022). However, subsistence hunting is not as selective as trophy hunting (Allendorf et al., 2008; Coltman et al., 2003). The presence of trophy hunting (or prestige hunting) in Roman and post-Roman times seems to be accepted (Colominas et al., 2017; Fernández-Rodríguez, 2003; Toynbee, 1973), with the consequent possible effect of size reduction. Even so, it is true that this type of hunting practices seem to have focused more on other taxa such as deer, wild boar or even bears or horses (Colominas et al., 2017; Fernández-Rodríguez, 2003; Toynbee, 1973). In fact, a lesser presence of chamois seems to be evident in the Cantabrian sites after the settlement of populations with an agro-livestock economy. Interestingly, the rise of the Roman Empire 2000 years ago triggered large-scale extinctions of other large mammals (as bears), followed by increasingly fast range loss and fragmentation (Albrecht et al., 2017). The debate on the existence of trophy hunting earlier, in prehistoric times, is still open (Baquedano et al., 2023; Brugal, 2016).
Furthermore, a strong relationship has been found between the habitat structure of chamois populations and the body size of their individuals, with the decrease in size being conditioned by the loss of forest mass (Reiner et al., 2021), and the decrease in food resources (Mason et al., 2014). As for bears, more data on chamois in the Cantabrian Mountains from the second half of the Holocene are needed to assess more precisely which factor is most implicated in the striking change in the mean size of its populations during this time.
In contrast to chamois, the gradual decline in the size of wild horses from the Middle Pleistocene to the Late Pleistocene seems to point to a climatic basis (Alberdi et al., 1995; Forsten, 1991, 1993). In this respect, the importance of substrate characteristics and the density of the vegetation through which they move is also well known as an important factor influencing changes in the size and body proportions of large ungulates (Alberdi et al., 1995; Eisenmann, 1984; Sorbelli et al., 2023). A marked difference in the size of horses in Iberian populations compared to those in the rest of Europe seems clear from the middle of the Middle Pleistocene. This smaller size of Iberian horses until the Holocene may be the result of an independent and isolated evolutionary history of Iberian populations (Uzunidis et al., 2024) or discordances in the characteristics of Iberian ecosystems compared to those of other areas of Europe in terms of substrate and vegetation (Diedrich, 2014; Kahlke et al., 2011; Puzachenko et al., 2021). It should be noted that, according to genetic data, approximately 150,000 years ago Equus ferus ferus and Equus przewallskii diverged somewhere in Eurasia (Goto et al., 2011). This information reinforces the consideration of the presence in this area of horse populations with large biological differences between them since the Middle Pleistocene. The biogeographical and ecological implications of this great regional complexity of horse populations in Eurasia are not well known at present (Lovász et al., 2021).
The large decrease in the average size of horses across Europe between the Late Pleistocene and the Holocene stands out. Considering the global climatic dynamics and evolutionary history of these equids, it seems necessary to consider the possible effect of the Last Glacial Maximum (Guthrie, 2003) and the potential human impact of domestication (Davis, 1981; Eisenmann and Beckouche, 1986) on this process. According to Forsten (1991, 1993), this decrease in size in European horses may also be related to the decrease in food availability and the hardening of environmental conditions. The effect of climate change and declining population sizes in horses seems to be sufficiently proven according to observations by Guthrie (2003), yet the exact causes of this phenomenon are not known in detail (Sommer et al., 2011). From a regional point of view, horse populations in the Cantabrian Mountains may have been affected by extreme habitat loss phenomena at the end of the Late Pleistocene. Typically, in this area wild horses have been considered to have a special relationship with lowland environments, such as the Duero Basin, the terraces and alluvial fans of the Cantabrian valleys or the marine terraces/lands of the Asturian coast (Álvarez-Lao, 2014; Mielgo et al., 2024; Rivals and Álvarez-Lao, 2018). Due to the incision of the hydrographic network in the Cantabrian Mountains and the retreat of the coastline, the extent of palaeosurfaces, terraces and alluvial fans seems to have been reduced from the Late Pleistocene to the Holocene (at least from MIS 4-3), reducing the extent of the flat areas (Adán et al., 2009; Ballesteros et al., 2017; Flor-Blanco et al., 2022; Jordá-Pardo et al., 2023). As for chamois, these habitat loss processes could be related to the decrease in size (Mason et al., 2014; Reiner et al., 2021).
The use of horses as part of the diet of human groups in different chronologies is well known (Carbajo, 2020; Galindo-Pellicena et al., 2017), as documented for chamois. The process of anthropogenic selection for domestication is likely a source of phenotypic changes during the Holocene (Davis, 1981; Gardeisen, 2005), and the small size of the individual horses of the Holocene wild populations are very similar to those of the first known domestic horses (Martín et al., 2016; Olsen, 2006; Zeder, 2012). This concurs with the case of the Llamazares horse. More complex to explain is the possible pathway of preferential selection of small-sized individuals in the domestication process, as horse domestication has generally tended to increase size and variability in domestic individuals (Kiesewalter, 1888; Tchernov and Horwitz, 1991). These reasons seem to point to a decline in size mainly due to climatic and ecosystemic reasons. This is reinforced by the late onset of horse domestication (ca. 5500 cal a BP; Outram et al., 2009) in different populations, including the Iberian Peninsula (Warmuth et al., 2011).
Overall, environmental and anthropogenic changes from the Pleistocene to the Holocene have led to smaller body size in mammals (Smith et al., 2018; Svenning et al., 2024). This process includes the loss of larger taxa in most areas of the planet (Albrecht et al., 2017; Brook and Barnosky, 2012; Smith et al., 2018; Stuart, 1991) and a decrease in the size of individuals of species that did not become extinct (Dale Guthrie, 2003; McNutt and Gusset, 2012). Although the latter phenomenon is not generalisable to all geographic regions (Prothero et al., 2012). In particular, the Cantabrian Mountains has suffered the effect of both processes. On the one hand, at the end of the Pleistocene most large carnivore species became regionally extinct (Sanchis et al., 2024; Sauque et al., 2017; Sauqué and Cuenca-Bescós, 2013; Stuart and Lister, 2007), with the exception of the brown bear (Nores and Naves, 1993) and the wolf (Blanco et al., 2007). More recently, during the Holocene, the disappearance of the last populations of aurochs Bos primigenius (Crees et al., 2016) and equids (Crees and Turvey, 2014; Sommer et al., 2011) left significant niche gaps among large herbivores (Plasteeva et al., 2020; Svenning et al., 2024).
Our study shows a decrease in individual body size of the species that survived in the Cantabrian ecosystems in the recent past. Further research should explore the actual array of affected species and the specific causes of size variation for each of them, but changes in climate, access to resources, habitat availability and direct anthropogenic pressure stand out as factors to be assessed (Dale Guthrie, 2003; Gardner et al., 2011; Marciszak and Gornig, 2024; Mason et al., 2014; McNutt and Gusset, 2012; Rode et al., 2010; Scheffer, 1955). The individuals from Cueva de Llamazares presented for the first time in this paper help us increase the resolution of the record of changes in brown bear, chamois and horse populations in chronologies that have been little studied from a biological point of view.
In turn, changes populations and individual parameters of large mammals can modify the structure of ecosystems (Sillero et al., 2022; Wilson et al., 2020). The extreme case of this phenomenon is the loss of niches with local, regional or global extinctions, as in the case of mammoths (Putshkov, 2003). Similar effects are potentially extensible to the cases of a wide variety of macromammals that have gone extinct or changed their population characteristics in recent times (Crees et al., 2016; Lorenzen et al., 2011; Plasteeva et al., 2020; Stuart and Lister, 2007; Svenning et al., 2024). The decline in size of bears, chamois and wild horses may have had major implications for both the productivity of ecosystems, through reduced survival of individuals and population size (Mason et al., 2014), potentially affecting ecosystem structure and functioning (Berger et al., 2001; Ordiz et al., 2013).
The definitive extinction of wild horses implied the loss of a very specific pattern of use and maintenance of grasslands, which is actually the reasons for ‘rewilding’ projects in Europe (e.g., Linnartz and Meissner, 2014; Sandom et al., 2014). These results highlight the importance of considering not only the change in the taxonomic diversity of ecosystems when assessing changes in their functioning, but also the importance of knowing the variations in the ecological parameters of the species that persist in those ecosystems (McNutt and Gusset, 2012). Monitoring the response of the different species that make up the communities under study over long chronological periods provides more accurate pictures of their potential roles in ecosystems and distribution ranges (Clavero and Delibes, 2013). These data are of vital importance if one intends to opt for ecosystem conservation through rewilding or population reinforcement (IUCN/SSC, 2013). In this context, palaeontological, archaeological and historical heritage studies from a biological perspective stand out as a very useful tool (Clavero and Delibes, 2013; Domínguez-García et al., 2024; Lazagabaster et al., 2022; Nores and Naves, 1993).
Supplemental Material
sj-png-1-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-1-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-png-2-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-2-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-png-3-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-3-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-png-4-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-4-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-png-5-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-5-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-png-6-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-png-6-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-xlsx-7-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-xlsx-7-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-xlsx-8-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-xlsx-8-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Supplemental Material
sj-xlsx-9-hol-10.1177_09596836251333290 – Supplemental material for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene
Supplemental material, sj-xlsx-9-hol-10.1177_09596836251333290 for Highlighting the drastic body size decline in Cantabrian Brown Bear, Chamois and Wild Horse during the Holocene by Darío Fidalgo, Carlos Fernández-Rodríguez, Daniel Ballesteros, Andrés Ordiz, Christian Pérez de la Viuda, Juncal A. Cruz, Sara García-Morato and Esperanza Fernández-Martínez in The Holocene
Footnotes
Acknowledgements
We would especially like to thank Cueva de Llamazares team (Sandra Fernández y Elena Fernández) for the facilities for the study of the cave and the support provided. We are grateful for the collaboration of the Laboratorio de Paleontología of the Universidad de León (especially Dr. Ismael Coronado). We would also like to thank Dr. Joan Madurell-Malapeira and Dr. Guillermo Rodríguez-Gómez for their friendly feedback on the work and methodologies. Special thanks to all the volunteers who have helped in the field. We thank the editor (Prof. John A. Matthews) and two anonymous referees for their contributions, which have helped to significantly improve the outcome of this work.
Author contributions
Darío Fidalgo: Conceptualisation; Data curation; Formal analysis; Funding acquisition; Investigation; Methodology; Project administration; Resources; Software; Supervision; Validation; Visualisation; Writing - original draft; Writing – review & editing
Carlos Fernández-Rodríguez: Data curation; Formal analysis; Investigation; Validation; Writing – review & editing
Daniel Ballesteros: Investigation; Validation; Writing – review & editing
Andrés Ordiz: Investigation; Validation; Writing – review & editing
Christian Pérez de la Viuda: Software; Visualisation; Writing – review & editing
Juncal A. Cruz: Validation; Writing – review & editing
Sara García-Morato: Validation; Writing – review & editing
Esperanza Fernández-Martínez: Investigation; Project administration; Resources; Supervision; Validation; Writing – review & editing
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research activity was funded by the Research Group Q-GEO (Geología Ambiental, Cuaternario y Geodiversidad, Universidad de León, León, Spain). D.F. was supported by the Ayuda del Programa de Formación de Profesorado Universitario (FPU20/03389) and is a Ph.D. student at the Programa de Doctorado en Biología at the Universidad Complutense de Madrid.
ORCID iDs
Data availability statement
All data used in the preparation of this paper are included in the supplementary material and links provided in the text.
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.