
Abstract
Multiple pre-Columbian (pre-1492 CE) archaeological sites now challenge the traditional portrayal of Amazonia as a ‘pristine wilderness’. This is especially true within the forest-savanna mosaic landscapes of lowland Bolivia, where the pre-Columbian Casarabe Culture constructed hundreds of settlement mounds, integrated with a dense causeway-canal network – one of the most complex, stratified societies yet discovered in Amazonia. Excavations at previous sites indicate that this culture sustained itself by practicing large-scale, maize-based agriculture. However, the Casarabe Culture’s mounds have also been found within the riparian forests abutting major river systems, where their inhabitants could have benefitted from greater access to forest resources and local fish species. To determine whether these differences influenced how the Casarabe Culture utilised the landscape, we conducted palaeoecological analysis on the sediments collected from Laguna Loma Suarez (LLS), an oxbow lake situated adjacent to a monumental habitation mound within these riparian forests. Our analysis reveals that, despite significant differences in natural resource availability, the Casarabe Culture continued to cultivate maize locally around LLS for over a millennium, between 280 BCE and 1130 CE, with anthropogenic fires largely restricted to the open savannas. Our record also suggests that the Casarabe Culture possibly delayed either forest recovery or natural forest encroachment until after the nearby settlement mound was abandoned. These findings, when compared with those of other sites in the region, show that maize was an important crop in pre-Columbian times, irrespective of major differences in natural resource availability across the complex forest-savanna mosaic settings of Amazonian Bolivia.
Introduction
A number of archaeological sites (Heckenberger et al., 2003; Iriarte et al., 2012; Rostain et al., 2024; Schaan, 2016; Science Panel for the Amazon (SPFA), 2021) now challenge the traditional portrayal of Amazonia as a ‘pristine wilderness’, little affected by its past human inhabitants (Denevan, 1992b; Meggers, 1971). Of particular interest in this regard is the Llanos de Moxos (LM; red box, Figure 1a), a large (120,000 km2) seasonally flooded savanna in southwestern Amazonia that possesses an extensive record of past human activity (Denevan, 2001; Lombardo et al., 2020). While today, this sparsely-populated landscape is primarily used for cattle ranching and mechanised rice agriculture (Denevan, 1963; Gobierno Autonomo del Beni (GAB), 2019; Lombardo, 2023), the LM was once home to multiple indigenous groups who constructed a wide variety of earthworks prior to European Contact (Carson et al., 2014; Denevan, 2001; Erickson, 2000a; Walker, 2008). The most striking of these earthworks can be found within the LM’s southeastern sector, a region occupied by the now-extinct Casarabe Culture between 400 and 1400 CE (Jaimes Betancourt, 2012, 2015). This culture favoured the construction of large settlement mounds (lomas; see Erickson, 2000b), which they embedded within a matrix of causeways, canals, and lakes. Recent LiDAR scans attest to the size and spatial complexity of these features (Prümers et al., 2022), with each mound estimated to cover an average 5.5 ha surface area and reach up to 20 m high (Lombardo and Prümers, 2010). Based on this evidence, the Casarabe Culture is regarded alongside the Upano Valley culture in Ecuador (Rostain et al., 2024), as one of the clearest examples of low-density agrarian urbanism in pre-Columbian Amazonia (Prümers et al., 2022).
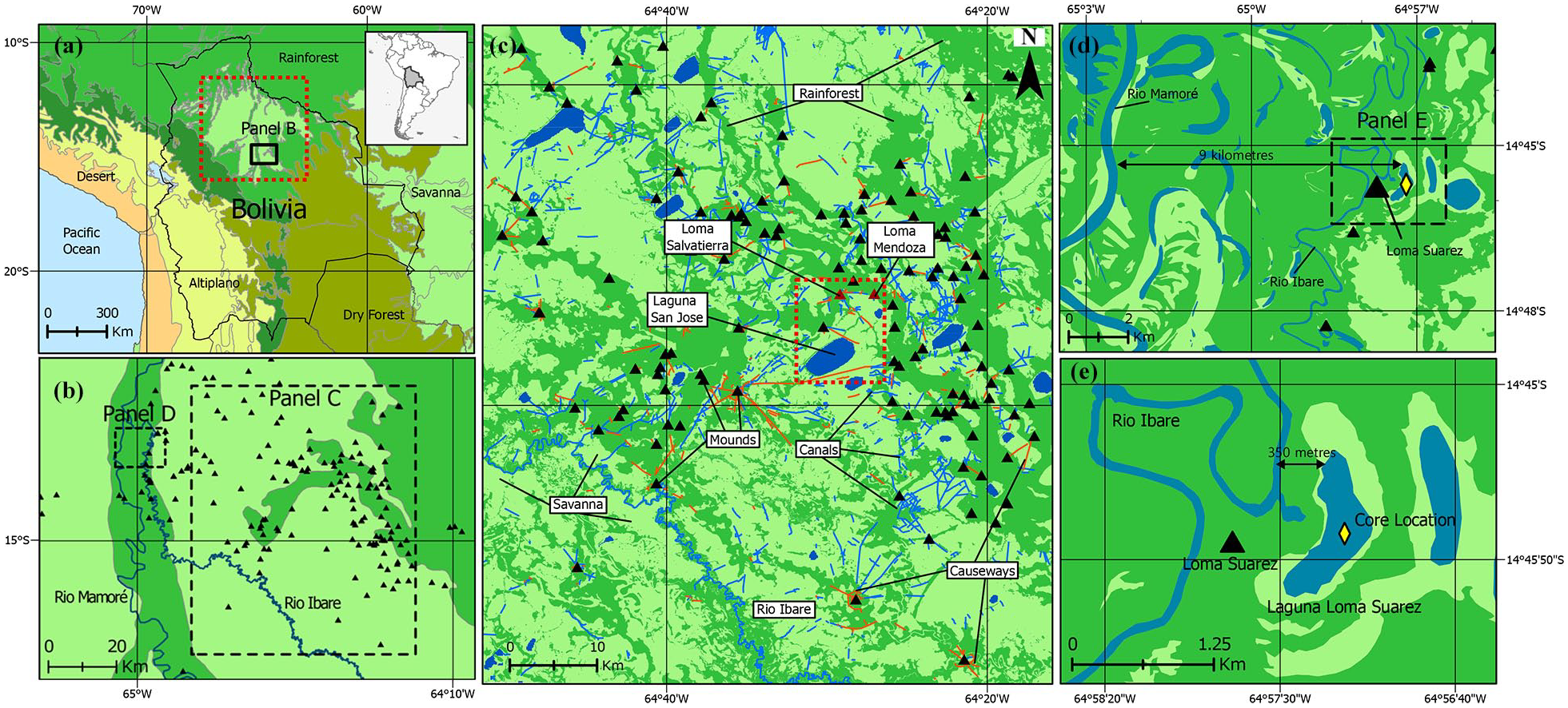
(a) Map displaying the location of our study area in relation to Amazonian Bolivia. Colours reflect a categorised version of the terrestrial ecoregions of Olson et al. (2001). Colours denote areas of: Tropical Forest (medium green); Savanna (light green); Yungas (dark green); Tropical Dry Forest (olive); Altiplano (yellow); and Desert (orange). (b) Map displaying the location of our study area (panel D) in relation to the region of earthworks mapped in detail by Lombardo and Prümers (2010) in the southeastern LM. Black triangles denote the location of monumental settlement mounds. (c) Map displaying the earthworks of the LM, modified from Lombardo and Prümers (2010). Black triangles denote the location of monumental settlement mounds, while blue and orange lines reflect identified canals and causeways respectively. The red box marks the locations of the two mounds, Loma Mendoza and Loma Salvatierra (both shown in red), which have been excavated in detail (Prümers, 2007, 2015), along with Laguna San Jose (LSJ; Whitney et al., 2013). Earthworks are plotted onto a Forest/Savanna land cover map (d) Map displaying our study area in relation to the Rio Mamoré. (e) Map displaying the location of our core site in relation to Loma Suarez and the Rio Ibare. In panels B–E, colours denote areas of: Tropical Forest (medium green); Savanna (light green); and open Water (blue).
While archaeobotanical evidence shows that the Casarabe Culture utilised a wide variety of crops, including squash (Cucurbita sp.), sweet potato (Ipomoea sp.), manioc (Manihot esculenta), yam (Dioscorea sp.), peanut (Arachis hypogaea), beans (Phaseolus vulgaris), cotton (Gossypium barabadense), peppers (Capsicum sp.), and various palm fruits (Arecaceae spp.), the sheer ubiquity of maize suggests that it represented a staple component of their diet (Bruno, 2010, 2015; Dickau et al., 2012). This culture is thought to have developed in the southeastern LM because, unlike the nutrient-poor soils found elsewhere in the LM (Lombardo et al., 2015), the region contains fertile fluvial sediments deposited during an avulsion of the nearby Rio Grande during the Late Holocene (Lombardo et al., 2012, 2013). Previous research proposes that the Casarabe Culture exploited these sediments by using their causeway-canal network as a drainage system to practice maize-based agriculture in the open savannas (Hermenegildo et al., 2024; Lombardo et al., 2012: 2025). However, it remains unclear whether they universally adopted the practice of maize cultivation across this diverse landscape. Just two neighbouring mounds (lomas) have been excavated in any detail (Prümers, 2007, 2015) and the only detailed palynological record available, obtained from Laguna San Jose (LSJ; Whitney et al., 2013), solely documents regional-scale palaeoenvironmental trends. Moreover, as shown in Figure 1c, all three of these sites are located within a small subsector of the southeastern LM (100 km2; see red box), away from any major watercourses and dominated by open savannas. The Casarabe Culture is also known to have constructed mounds in more heavily forested areas, including in the varzea (riparian) forests running parallel to the Rio Mamoré and the Rio Ibare in the south-central LM (Prümers et al., 2022). Settling in this environment would have provided access to a variety of natural resources, including edible arboreal fruits from the nearby riparian forests and the fish available within both rivers (Lauzanne and Loubens, 1985), but whether maize remained a key crop in this quite different environmental setting remains unknown.
In this study, we present the results of palaeoecological analysis carried out on a sediment core from Laguna Loma Suarez (LLS), a small oxbow lake situated adjacent to a settlement mound in the riparian forests abutting these rivers. Our aims are twofold:
To determine whether the Casarabe Culture cultivated maize and/or other crops in the local vicinity of LLS. We hypothesise that the local abundance of aquatic and riparian resources (e.g. arboreal fruits, fish) led the inhabitants of Loma Suarez to adopt different subsistence strategies compared with the maize-based agrarian economy in the savanna-dominated heartland of the Casarabe Culture further east, away from major rivers.
To establish the extent to which these people cleared the local riparian forests. Given the close proximity of a large habitation mound, we hypothesise that the construction and occupation of the nearby Loma Suarez habitation mound necessitated local-scale forest clearance.
Study area
LLS is a small (0.5 km2) oxbow lake situated 9 km from the Rio Mamoré and 350 m east of the Rio Ibare in the south-central LM (14°45′56.31″S, 64°57′20.51″W; Figure 1b, see panel D). The local climate is tropical, with temperatures averaging 25.2°C except for when cold air masses periodically encroach from the south (sures or surazos), causing them to temporarily drop as low as 10°C (Hanagarth, 1993). Precipitation is highly seasonal, averaging 1850 mm yr−1, but with the majority concentrated between December and March (Fick and Hijmans, 2017; Pouilly et al., 2004). Due to a combination of the LM’s flat topography (slope = <10 cm km−1) and its impermeable clay soil (Langstroth Plotkin, 2011; Lombardo et al., 2012), this precipitation causes up to 78,460 km2 of the region to flood annually (Hamilton et al., 2004). Being situated adjacent to two large rivers that regularly overtop their banks (Bourrel et al., 1999, 2004, 2009; Figure 1d and e), LLS is particularly susceptible to seasonal flooding. Despite possessing no permanent inflows or outflows, watermarks on the trees surrounding the lake indicate that flood levels regularly vary by as much as 3 m, connecting it to the adjacent rivers during the rainy season. However, these two rivers significantly differ in character. While the whitewater Mamoré originates in the Andes and is a major tributary of the Amazon, the clearwater Ibare’s head-waters lie 180 km southeast of LLS and primarily serves to drain floodwater from the open savannas (Loubens et al., 1992). Both rivers are also lined by distinct vegetation communities, with Navarro and Maldonado (2002: 183–186) describing two main types of local riparian forest: (i) a whitewater floodplain forest lining the Rio Mamoré, dominated by Hura crepitans, Pithecellobium corymbosum, and Calophyllum brasiliensis and (ii) a clearwater forest surrounding the Rio Ibare, dominated by Calycophyllum spruceanum (also see Castellon (1999)).
Lying adjacent to LLS is Loma Suarez (Figure 1e), a 6 m tall settlement mound built by the Casarabe Culture that today hosts a village of the same name. No archaeological excavations have yet been carried out here, though it is assumed that the mound was occupied over a similar time frame (400–1400 CE) as recorded at the excavated Mendoza and Salvatierra habitation mounds to the east (Jaimes Betancourt, 2012, 2015; Figure 1c). Loma Suarez has been continuously inhabited since the colonial period, when it was sold to become a livestock and administrative centre (Parada, 2013). The local environment around LLS remains in use today by the inhabitants of this mound, who regularly burn and clear localised land patches for cattle grazing.
Methods
Sediment collection and field surveys
An 82 cm long sediment core was extracted from LLS in July 2011 in two separate, overlapping drives. The upper drive (0–62 cm below the sediment-water interface) was obtained using a standard surface corer consisting of a Perspex tube and piston, with the uppermost 18 cm extruded into vials in consecutive 0.5 cm increments due to its high water content. The lower, overlapping drive (42–82 cm) was collected using a Colinvaux-Vohnout Livingston piston corer (Colinvaux et al., 1999). A subsequent visit to the site (September 2022) was made to conduct a rapid local vegetation survey within a 100-m radius of the lake shoreline to aid in interpreting the pollen assemblages.
Chronology
In total, 14 samples were selected for AMS 14C radiocarbon dating (Table 2), all of which were bulk-sediment samples due to the absence of terrestrial macrofossils or sufficiently large charcoal particles. Twelve of these dates were processed at the Oxford Radiocarbon facility based solely on the insoluble (humin) carbon fraction. The remaining two dates were analysed at the Beta Analytic laboratory, and performed on a combination of the insoluble and soluble (humic) fractions. While previous research has shown any difference registered between these fractions should be comparatively minor for sediments in the LM (May et al., 2015), we nonetheless emphasise that the potential downward mobility of soluble carbon means these two dates should be interpreted as minimum ages. The absence of carbonate bedrock means that the possibility of contamination due to the hard-water effect can be excluded. All ages were calibrated using the SHCal20 curve (Hogg et al., 2020) and placed within a Bayesian model using OxCal 4.4.4 (Bronk Ramsey, 2009a). The Poisson deposition model created includes a central k parameter estimate of 1, an interpolation rate of 2, and a k distribution of U (−2, 2; Bronk Ramsey, 2008). The latter allows k to vary by two orders of magnitude in either direction (from 0.01 to 100). In addition, a ‘General’ outlier model was applied to all dates, with each being given a 5% prior probability of being an outlier (Bronk Ramsey, 2009b).
Pollen and charcoal
Pollen analysis was undertaken at 2 cm resolution, prioritising the surface core where possible due to its better pollen preservation. 1 cm3 samples were prepared according to a modified version of the standard protocol (Faegri and Iversen, 1989) described in Whitney et al. (2012), which includes an additional coarse-sieving stage (>53 μm) to concentrate cultigen pollen grains. Exotic Lycopodium spores were added to enable pollen concentration calculations (Stockmarr, 1971), while pollen percentages were based on a sum of at least 300 terrestrial grains. When recording the presence of cultigen grains, twice as many Lycopodium spores were counted in the coarse fraction as in the fine fraction. Pollen grains were identified using the Neotropical pollen reference collection at the University of Reading, pollen atlases (Colinvaux et al., 1999; Lorente et al., 2017; Roubik and Moreno, 1991), and the digital catalogue developed by Bush and Weng (2007). The criteria of Holst et al. (2007) were used for maize pollen identification.
Charcoal analysis was conducted in contiguous 0.5 cm intervals, with samples prepared according to a modified version of the protocol by Whitlock and Larsen (2001; Stevenson and Haberle, 2005). Hot sodium hexametaphosphate (Na6[(PO3)6], 80°C) was used to disaggregate the clay-dominated sediments, followed by immersion in hydrogen peroxide (H2O2) overnight to bleach organics. Both the 125–250 and >250 μm charcoal size fractions were counted. ‘Herb’, ‘Wood’ and ‘Leaf’ charcoal fragments were differentiated according to the characteristics of Umbanhowar and Mcgrath (1998), with unidentifiable charcoal particles categorised as ‘Other’.
Sediment characteristics and geochemistry
X-ray Fluorescence core scanning (XRF) analysis was performed on the sediment record from LLS to: (i) provide a basis for precise cross-correlation between the surface core and adjacent overlapping Livingstone core, and (ii) serve as a proxy for catchment erosion. We hypothesised that the rate of catchment erosion would have been highest during the period when Loma Suarez was occupied – due to maintenance of open habitation areas on the mound leading to enhanced surface runoff – before declining due to forest regrowth following the Casarabe Culture’s collapse. XRF is well-suited to detecting changes in catchment erosion, since it can indicate the concentrations of several elements (Si, K, Ti, Fe, Rb, Zr) commonly associated with the deposition of detrital lithogenic material (Davies et al., 2015). This analysis was conducted at 2 mm resolution on both cores at the British Ocean Sediment Core Research Facility (BOSCORF) using an Itrax core scanner (Croudace et al., 2006), with a Molybdenum (Mo) tube set at 30 kV/30 mA and an average exposure time of 15 s (Jarvis et al., 2015). Measurements returned with counts per second (CPS) values beyond 1.5 standard deviations of the dataset mean were considered errors and thus excluded (~5% data loss). We present our results as log-ratios to establish a direct relationship with geochemical composition (Weltje and Tjallingii, 2008). As with previous work undertaken in the LM (Giesche et al., 2020), we base our palaeoenvironmental interpretations on the lithogenic and hydromorphic elements present within the sediment.
To determine water and organic content, we oven-dried the samples at 110°C and conducted loss-on-ignition (LOI) at 550°C, respectively (Dean, 1974; Heiri et al., 2001), both in consecutive 0.5 cm increments. High-resolution core photographs, with quantitative colour data, and magnetic susceptibility analysis were also undertaken at BOSCORF, and can be found within the supplementary information (Figures S8 and S9).
Numerical analysis
We employed a CONISS agglomerative clustering algorithm to separate pollen data into statistically significant zones (Grimm, 1987). To determine whether the pollen data from LLS respond either linearly or unimodally with regards to species turnover, we employed a detrended correspondence analysis (DCA; Hill and Gauch, 1980; Legendre and Legendre, 2012). The resulting length of the primary axis, which is thought to approximate β-diversity, was 1.06, indicating that pollen exhibited a linear response. On this basis, we performed a principal components analysis (PCA) on the correlation matrix of our pollen data to explore its main underlying trends without down-weighting rare taxa. This analysis was carried out using the vegan package (version 2.6.4) in R version 4.3.1 (Oksanen et al., 2022; R Core Team, 2023). Both analyses consider all taxa with a maximum abundance of ⩾1%. Prior to the analysis, the pollen data were transformed using the Hellinger distance metric (Legendre and Gallagher, 2001).
A PCA was also conducted for our geochemical data utilising the ItraxR package (Bishop, 2023). Elements were selected for inclusion based upon an autocorrelation function with a 10-sample lag period. Prior to analysis, data were transformed using the ‘robust centre log-ratio’ method to mitigate the effects of compositionality bias (Aitchison, 1986). In both cases, PCA biplots were set to maximise the accuracy of site scores to aid in identifying compositional changes through time.
Results
Rapid vegetation survey
During our rapid vegetation survey, we identified 36 species belonging to 20 families in the local vicinity of LLS (Table 1). Vegetation at the site is typical of riparian forest, aside from a 10 to 100 m strip of herbs around the lake shoreline. The identified taxa were distributed across three distinct vegetation communities similar to those described by Navarro and Maldonado (2002). The immediate shoreline was characterised by herbaceous taxa common to both semi-aquatic (e.g. Eichhornia crassipes and Enydra fluctuans) and open terrestrial (e.g. Trifolium sp., Polygonum spp.) plant communities. Approximately 20–50 m beyond this zone lies a community of pioneer shrubs, comprising a mixture of flood-tolerant (e.g. Erythrina fusca) and thorny scrub taxa (e.g. Mimosa pigra and Combretum laxum). To the west of LLS is a third plant community comprised of arboreal taxa typical of riparian forest (e.g. Maprounea guianensis), with evidence of recent disturbance (e.g. Cecropia trees and Brachiaria grass commonly grown for cattle grazing). An arboreal community also lines the eastern shore of LLS, but was inaccessible and therefore could not be surveyed.
Species recorded in the rapid vegetation survey conducted around the shoreline of LLS. Habitat codes denote where taxa were located: (1) Herbaceous/Aquatic shoreline vegetation; (2) Pioneer scrub; (3) Whitewater riparian forest; (4) Clearwater forest. Abundance values: +++++ = >20 individuals counted; ++++ = 10–20 individuals counted; +++ = 5–10 individuals counted; ++ = 2–5 individuals counted; + = 1 individual counted.
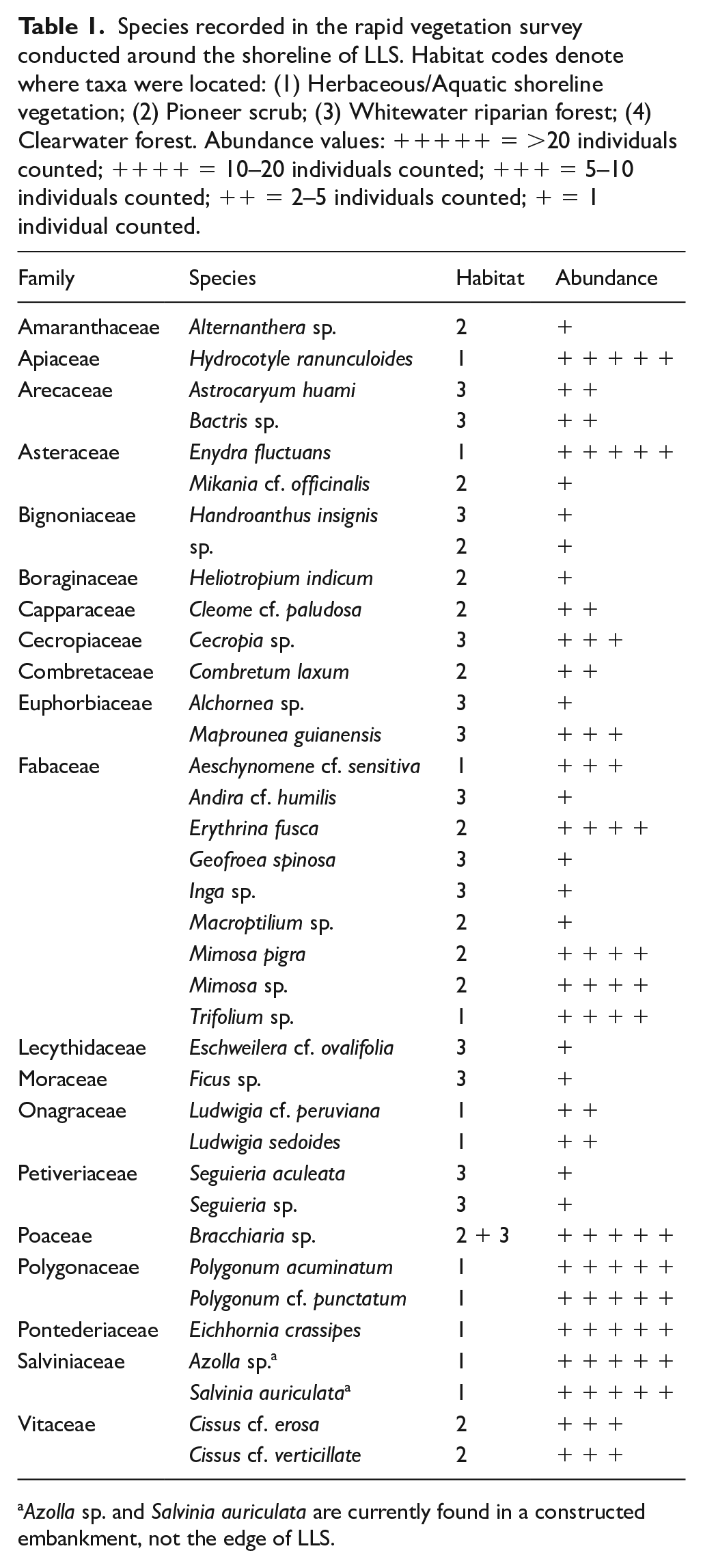
Azolla sp. and Salvinia auriculata are currently found in a constructed embankment, not the edge of LLS.
Stratigraphy and chronology
Using the geochemical data obtained from XRF analysis (see Figure S8), we successfully cross-correlated the two overlapping cores comprising the 82 cm sediment record from LLS. This record primarily consists of dark yellowish brown to dark greyish brown clays (Munsell Soil Colour Codes 10YR 4/4 and 2.5Y 4/2, respectively; LOI 550°C = 6–9%). LOI values decrease, and the sediment becomes more greyish with increasing depth, becoming particularly dark between 63 and 72 cm (LOI 550°C = 10–12%). The lowermost 12 cm differs notably from the rest of the record, characterised by its colour (2.5Y 5/3 light olive brown) and comparatively low LOI 550°C values (4%–6%; see Figure S8).
The 14 samples sent for radiocarbon dating initially returned conflicting 14C ages, making it impossible to generate a Bayesian age-depth model. Particularly anomalous are the ages obtained from the sediment unit between 43 and 50 cm, which are either modern or incompatibly young compared with the samples collected above and below it. The characteristics of this sediment unit are notably distinct from the surrounding material, consisting of olive brown (2.5Y 4/3) clays with a reddish-brown mottling typically associated with iron precipitation, and demarcated by sharp contact boundaries (Figure 2). These characteristics, along with its higher water content (56%–60%) are more consistent with sediment at the sediment-water interface. Additionally, pollen assemblages from this unit are similar to those of the surface sediments, especially with respect to Cecropia and Ipomoea percentages. Based on these anomalies, we infer that this sediment unit reflects the intrusion of modern material into the core, and have therefore excluded it from our palaeoenvironmental reconstruction. In the graphs presented below, the maximum depth of sediment below 50 cm has been reduced by 7 cm to account for its removal. The full set of radiocarbon ages, including four from the removed unit, are provided in Table 2. A full palaeoecological diagram incorporating the removed sediment can also be found within the Supplemental Information (SI; Figures S3 and S4).

High-resolution photograph of the surface core recovered from LLS. A red box highlights the sediment unit removed from our sequence, with grey dotted lines demarcating its sharp upper and lower boundaries. Details on how the core was photographed are provided in the Supplemental Information.
Conventional and calibrated radiocarbon dates for samples obtained from LLS, calibrated using the SHCal20 calibration curve (Hogg et al., 2020). Bayesian radiocarbon age models are available for this record within the Supplemental Information (Figures S1 and S2). Calibrated age range follows 95.4% CI.
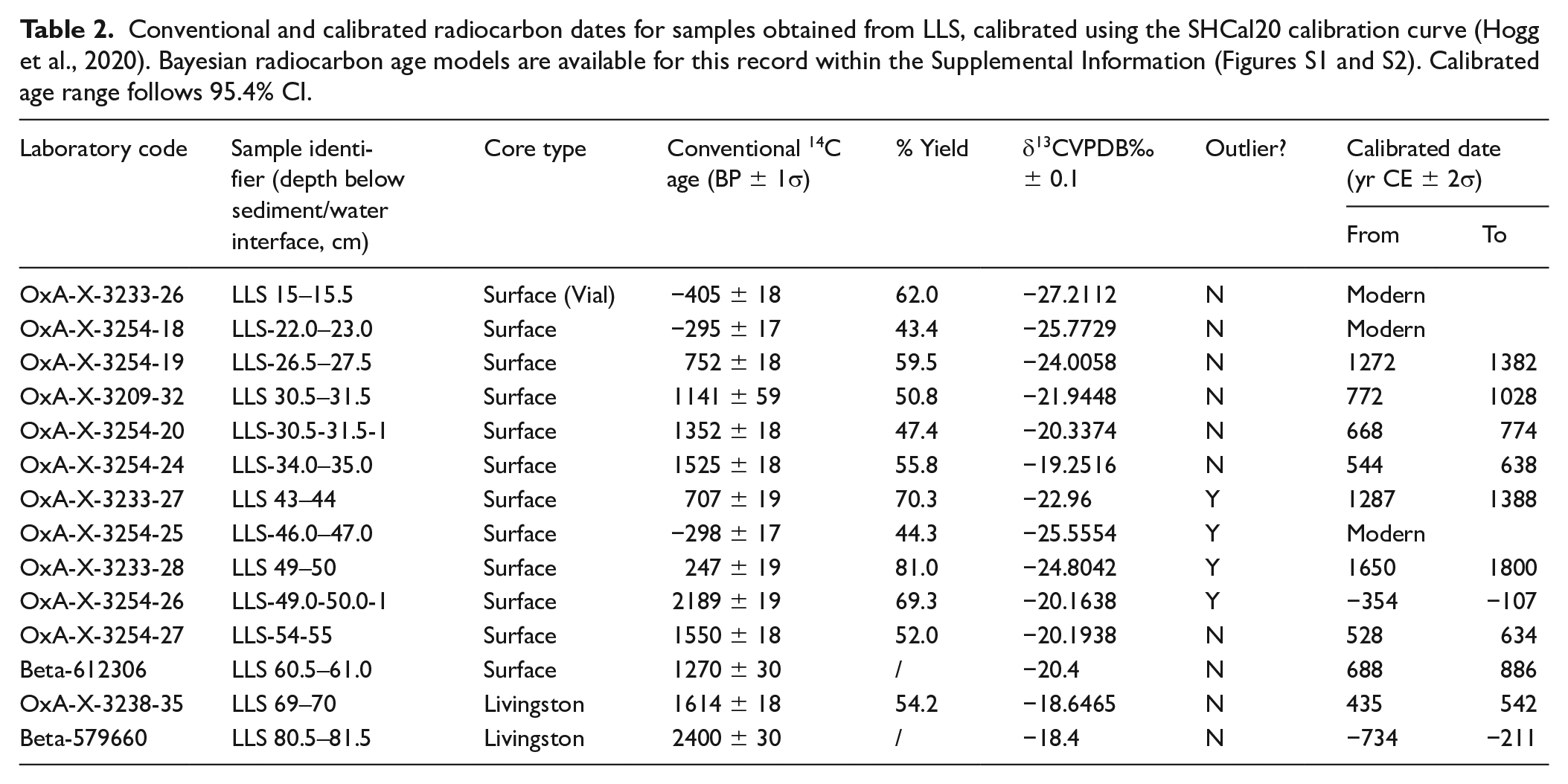
After excluding this sediment unit and its associated dates, it became possible to produce a Bayesian age-depth model for the sequence. With a maximum age of between 734 and 211 BCE, the sedimentation rate at LLS is broadly consistent with other small lakes (Carson et al., 2014; Loughlin et al., 2021) and faster than that of much larger lakes in the LM (Brugger et al., 2016; Whitney et al., 2013). One date (Beta-612306) was flagged as a likely outlier (posterior: 99%), which may be due to radiocarbon analysis being conducted on both the humic and humin fractions of the sample, resulting in an anomalously young date. A large number of dates are concentrated between 400 and 900 CE, indicating an order-of-magnitude increase in sedimentation rate (see Figure S10). Pollen concentrations also increase at this time (Figure 3), which is unusual as one would expect them to decline as the deposited grains become diluted by the additional supply of sediment. Due to this discrepancy, we plot both pollen percentages and charcoal concentration versus depth rather than as influx versus age. For transparency, a summary influx diagram can be found in Figure S5.
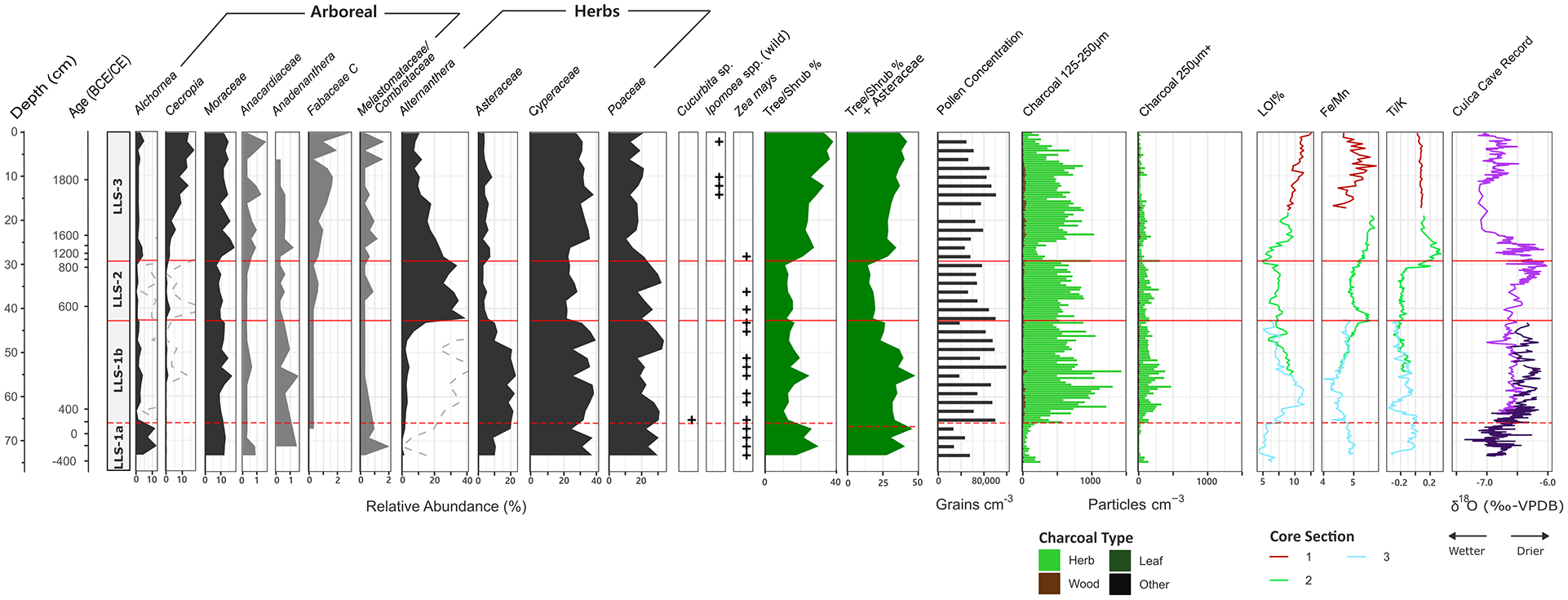
Summary diagram of key pollen types at LLS, including presence/absence data for Zea mays, Cucurbita sp., and Ipomoea sp. grains concentrated using the methodology of Whitney et al. (2012). Dashed curves exaggerate variations in low abundance pollen taxa. The diagram also includes variations in charcoal concentration (125–250 μm and 250+ μm size fractions), loss-on-ignition, and XRF ratio data. Our data are plotted against the δ18O spelaeothem record from Cuica cave (Della Libera et al., 2022). A concentration diagram for this record is provided within the Supplemental Information (Figure S6).
Pollen and charcoal
Pollen preservation is generally reasonable throughout the record (see Figure 3). Although signs of deterioration were observed in the lowermost 12 cm, the pollen was still identifiable. The record is dominated by herbaceous taxa, most notably Cyperaceae (21%–40%), Alternanthera (0%–39%), Poaceae (11%–34%) and Asteraceae (3%–23%). Less common, but still consistently present, are Mimosa (0%–1%) and Amaranthaceae (0%–2%). Common arboreal taxa include Moraceae (7%–18%), Cecropia (0%–17%) and Alchornea (0%–13%), though typical tropical dry forest elements such as Acalypha (0%–1%) and Anadenanthera (0%–2%) were also regularly found. The charcoal record is overwhelmingly dominated by herbaceous particles, which consistently comprise 80%–100% of total charcoal counts.
Our agglomerative clustering algorithm revealed three statistically significant pollen assemblage zones, which we describe below:
LLS-1 (73.5–42.5 cm; 430 cal. yr BCE–590 cal. yr CE)
LLS-1 is characterised by its relatively high proportion of Asteraceae (9%–23%) and Cyperaceae (25%–40%), while Alternanthera (0%–7%) and Cecropia (0%–1%) are virtually absent. Although not statistically significant, LLS-1 can be further divided into two subzones following the abrupt sedimentological transition between the lowermost 12 cm of light olive brown clays and the rest of the record. The lower subzone is distinguished by relatively low overall pollen concentrations and high proportions of Alchornea (4%–12%). By contrast, the upper subzone is marked by the high percentages of Asteraceae, Cyperaceae and Poaceae. The change between these two subzones is driven by an increase in overall pollen concentration; while Alchornea percentages slightly decrease into the upper subzone, no other reductions are observed. A similar two-stage pattern is observed within the charcoal record. Concentrations rarely exceed 300 particles cm−3 within the lower subzone, but quintuple to 1500 particles cm−3 immediately following the transition. This decreases to 1000 particles cm−3 towards the upper zone boundary.
Zea mays is consistently present throughout LLS-1, beginning from the lowest sample analysed. We also record a single Cucurbita grain, though its degraded condition meant that we were unable to determine whether it belonged to a domesticated variety or wild relative.
LLS-2 (42.5–30 cm; 590–840 cal. yr CE)
This zone is marked by an abrupt and substantial increase in Alternanthera (23%–39%), replacing Asteraceae (3%–7%) and to a lesser extent Cyperaceae (21%–28%). Minor reductions are also observed in both Moraceae and Anadenanthera, though note the latter taxon is already rare within the record. Little change is observed in charcoal concentrations from the upper subzone of LLS-1. Zea mays remains present at low abundance throughout the zone, but becomes less consistent towards the surface. It last appears immediately following the transition into LLS-3.
LLS-3 (30–0 cm; 840 cal. yr CE–present)
The transition to LLS-3 is marked by a reduction in Poaceae (27%–17%) and Alternanthera (18%–7%), replaced by Moraceae (10%–15%) and Cyperaceae (18%–30%). A gradual, sustained rise in Cecropia is also observed from 3% to 17% throughout the zone. Increases are recorded in the number and variety of tree/shrub taxa, including in Melastomataceae/Combretaceae, Fabaceae, and Anacardiaceae (e.g. Schinus and Astronium). By contrast, Zea mays is absent and, while we note the presence of Ipomoea, these grains lack the diagnostic bottle-shaped echinae of I. batatas (Bush and Weng, 2007; Roubik and Moreno, 1991) and therefore likely belong to a wild variety. Aside from a temporary reduction at the bottom of LLS-3, charcoal concentrations remain consistent with those in LLS-2, before gradually decreasing towards the surface.
Ordination
Our pollen PCA is estimated to capture 47.4% of the total variance (Figure 4). The samples from LLS are most clearly separated by the first principal axis, which ascribes positive values to samples from LLS-1 and negative values to samples from the other two zones. LLS-1 is shown to exhibit a positive correlation with Asteraceae, Moraceae, Cyperaceae, and Alchornea. Meanwhile, LLS-2 and LLS-3 form a tight cluster that is difficult to disentangle. This cluster is more closely correlated with tree/shrub taxa found in the latter zone (e.g. Cecropia and Anacardiaceae) as well as with Alternanthera. The distinction made by the second principal axis is much less clear, though tree/shrub taxa (e.g. Anadenanthera, Alchornea and Moraceae) are typically ascribed positive loadings, while herbaceous taxa (e.g. Cyperaceae and Poaceae) are negative. However, Asteraceae is assigned a positive score, more closely aligning to arboreal taxa.
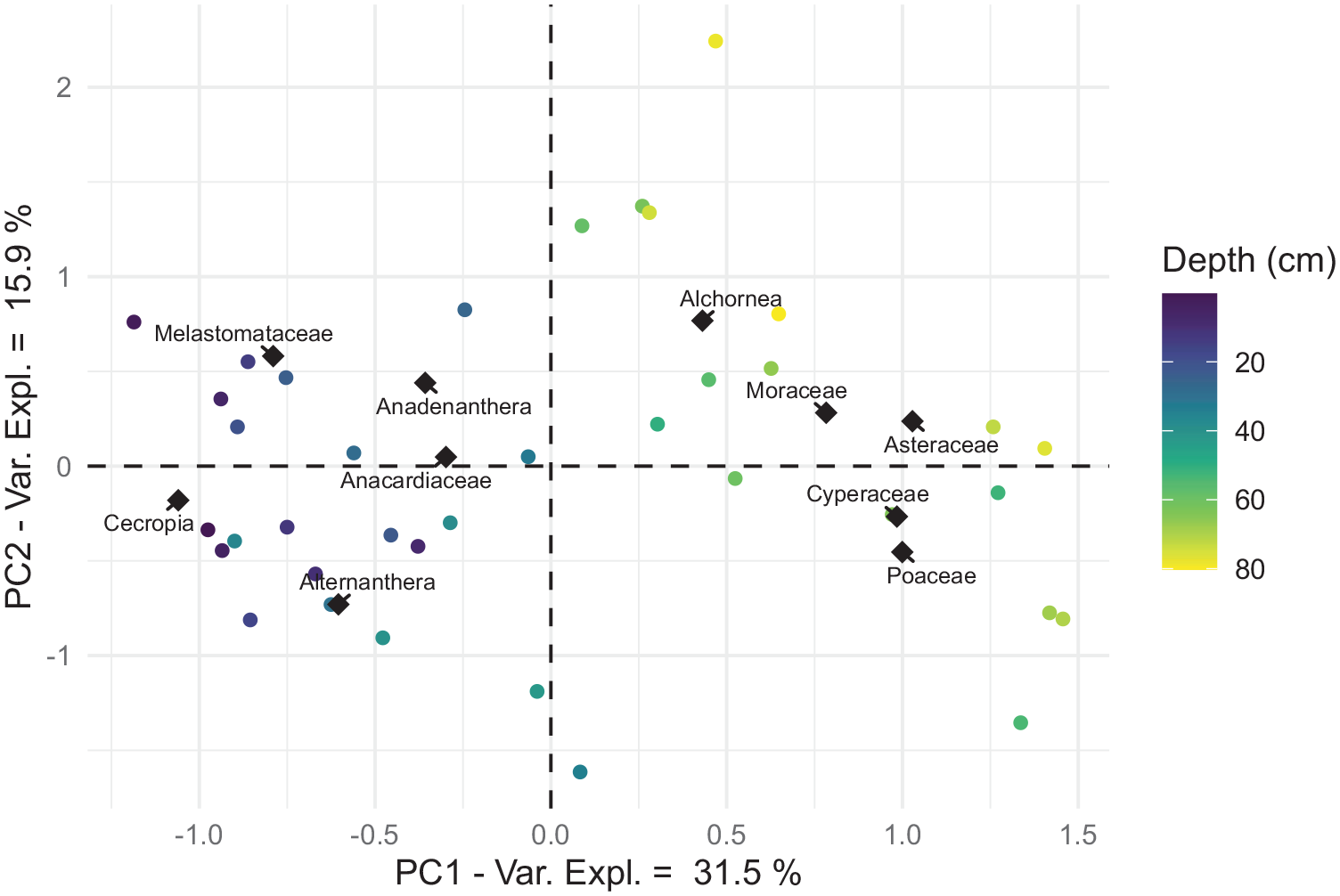
PCA biplot of pollen samples from LLS. Each point reflects a single pollen sample, with colour relating to the depth of the corresponding sample. Position of pollen taxa relates to the contribution and correlation of these taxa towards principal axes. The diamond shapes represent the locations of these taxa projected into sample space.
Lake sediment geochemistry
The first two axes of our geochemical PCA, which together capture 54.4% of total variance, separate the LLS sequence into three clusters (Figure 5). The samples from between 65 and 30 cm are observed to form a group close to the origin, indicating that few changes in geochemical composition occur during this period. However, a subset of this group also displays more negative scores on the second principal axis, which more closely correlates with a group of elements characterised by Mn, Zn and Ca. A second, tighter cluster is made by the sediments pertaining to the light olive brown clays in the lowermost 12 cm of the record. This sediment more closely correlates with Sr, Si and a group of elements characterised by K. A final, more disparate cluster is formed from the samples less than 30 cm in depth, characterised by the closer correlation with Ti, Zr and the Mo ‘coherent’ and ‘incoherent’ (Compton) scattered radiation.
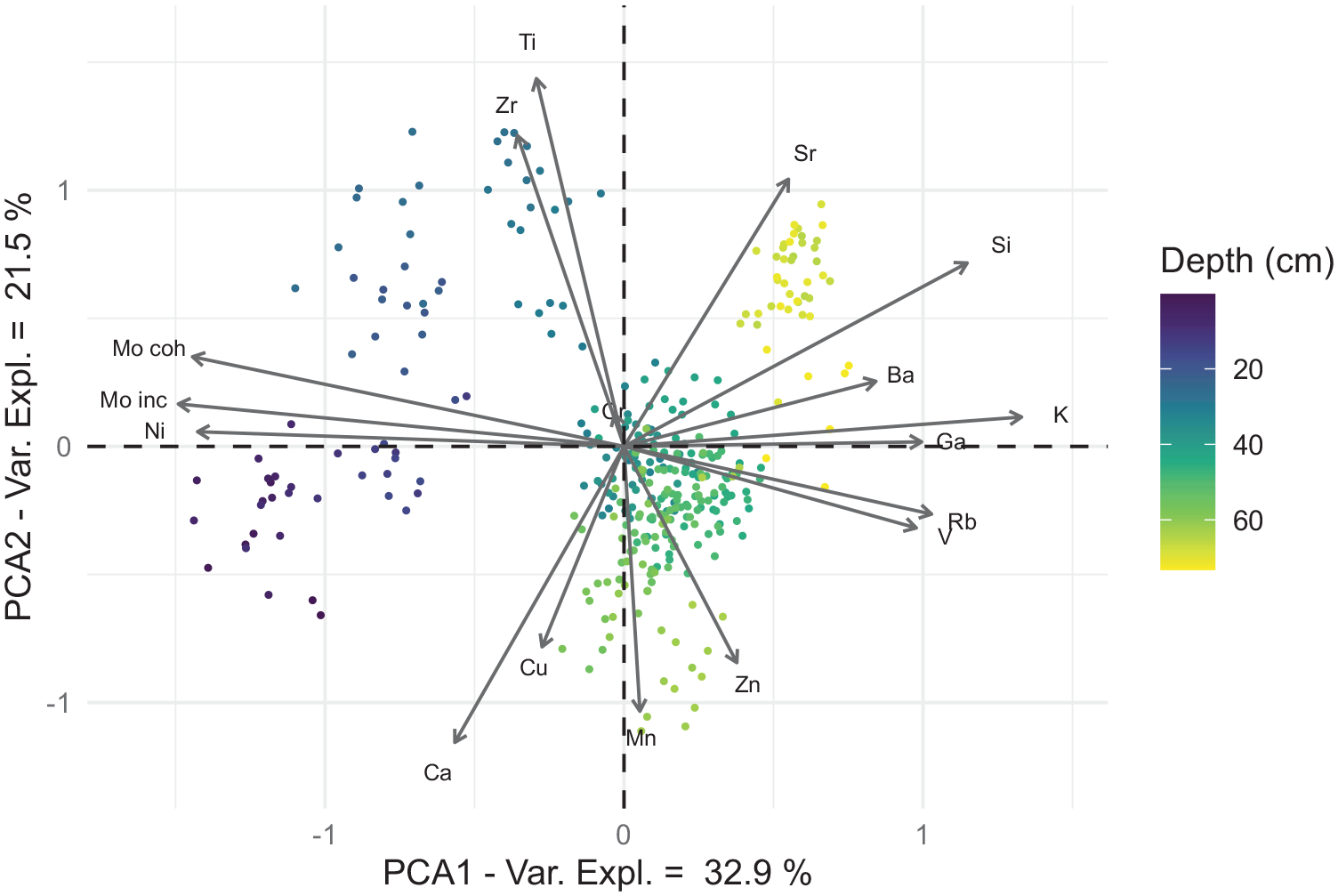
PCA biplot of geochemical samples from LLS. Each point reflects a single geochemical sample, with colour relating to its corresponding depth. Position of chemical elements relates to their contribution and correlation with principal axes.
Interpretation
The complex nature of the sediment record recovered from LLS makes interpretation of its underlying trends challenging. Past changes in pollen and charcoal data are not necessarily aligned to one another, or to changes in sedimentological characteristics. This is further compounded by LLS’s complex local hydrological setting. The sediments deposited within this lake could potentially originate from: (i) the local terrestrial environment surrounding it, (ii) the Rio Ibare, (iii) the Rio Mamoré, by backflow into the channel of the Ibare, as recorded by the hydrological station upstream of LLS at Puerto Almacen (Bourrel et al., 2009) or (iv) any combination of the above. Nonetheless, our multiproxy approach enables us to reconstruct environmental change and human activity here over the past 2.4 millennia, even despite this complexity.
Geochemistry
The first principal axis of our PCA displays a positive correlation between Si, K and Rb (Figure 4), as well as their negative relationship with Mo scattered radiation. K and Rb are commonly found in clay minerals (Zaunbrecher et al., 2015), the primary constituent of the sediment at LLS. This relationship is consistent with local sedimentological studies (Boixadera et al., 2003; May et al., 2015), which indicate that the most common clay type in this area is illite, a K-based mineral that can incorporate Si and Rb into its lattice structure. In contrast, the ratio between Mo ‘incoherent’ and ‘coherent’ scattered radiation is commonly used as a geochemical proxy responding to changes in organic and moisture content, though its sensitivity varies significantly from lake to lake (Woodward and Gadd, 2019). Applying a linear regression to these variables (see Figure S11), we observe that an estimated 80.2% of this ratio’s variance at LLS can be explained through changes in water content (r2 = 0.802, n = 217). On this basis, we interpret the first principal axis of our PCA to reflect variations in the sediment matrix, distinguishing the clay sediment from its moisture content.
The second principal axis emphasises the positive association between Ti, Zr and Sr (Figure 4), as well as their negative correlation with Mn and Ca. Ti and Zr are both considered chemically immobile and thus commonly attributed to the deposition of lithogenic material from the wider lake catchment, particularly in relation to coarser grain sizes (Davies et al., 2015). It is unclear whether this latter interpretation applies at LLS because, while local sedimentological studies corroborate a link between Zr and coarser sediment (Giesche et al., 2020; May et al., 2015), no obvious changes in grain size are displayed within our record. However, the former interpretation is supported by their negative relationship with Mn, an element commonly associated with changes in in-lake redoximorphic conditions (Boyle, 2002; Davison, 1993). Within the LM, Fe and Mn both precipitate as concretions in response to changes in hydromorphic conditions, such as those caused by regular seasonal inundation (Boixadera et al., 2003; May et al., 2015). For this reason, we interpret the second principal axis to represent the balance between material deposited due to localised environmental processes, versus the input of lithogenic material from the wider lake catchment. This interpretation is also consistent with the neutral score of clay-related elements, since illite is found both within the local soils (Boixadera et al., 2003) and as part of the suspended sediment carried by the nearby rivers (Guyot, 1993). Following this interpretation, we utilise the Ti/K ratio below as a proxy for the proportion of lithogenic material being washed into LLS from the wider lake catchment.
Our hypothesis that catchment erosion indicators (peak Ti/K ratio and minimum LOI 550°C values) correlates with the Casarabe occupation/land-use phase (zones LLS 1–2), due to maximum anthropogenic disturbance of the mound, is not supported by our data. Instead, these erosion indicators peak during the lower third of LLS-3, when we predict that forest re-growth following abandonment of the mound would stabilise the slopes and hence lead to reduced, rather than enhanced, catchment erosion (discussed below). We offer no ready explanation for this unexpected finding, and can only infer that these geochemical changes must instead reflect other types of catchment dynamics.
Pollen and charcoal
The pollen sequences recovered from small lakes, such as LLS, typically respond to local-scale changes in vegetation composition (Jacobson and Bradshaw, 1981; Sugita, 1994). However, while the relative abundances of Cecropia, Cyperaceae, Moraceae and Poaceae in our uppermost pollen sample are consistent with lake surface sediment samples previously collected from LLS (Burn et al., 2010), we identified few plants of these taxa during our rapid survey of the local vegetation. We found no members of Cyperaceae and only one member of Poaceae. Moreover, the only member of Moraceae we identified, Ficus, is severely underrepresented in pollen records due to its cleistogamous flowers. Given that we were unable to access the arboreal community on LLS’s eastern shore, it remains possible that Moraceae was more abundant there. However, as Cecropia and Moraceae are prolific pollen producers (Gosling et al., 2009; Jones et al., 2011) and both Cyperaceae and Poaceae dominate the open, seasonally-flooded savannas (Hanagarth, 1993), it is likely that the pollen catchment of LLS contains a significant regional-scale component for these wind-pollinated taxa.
With this in mind, we now present a vegetation reconstruction for each assemblage zone:
LLS-1 (73.5–42.5 cm; 430 cal. yr BCE–590 cal. yr CE)
The pollen assemblage of LLS-1 indicates that the local environment around LLS was a forest-savanna mosaic, with the 9%–14% abundance of Moraceae being significantly below the 40% threshold for closed-canopy rainforest (Gosling et al., 2009). However, the floristic composition of this mosaic was different from that of today, as evidenced by the lower percentage of Moraceae (11%) and higher percentages of Alchornea (10%) and Asteraceae (20%) pollen compared with the surface pollen assemblage (ca. 15%, 5% and 5% respectively). It is difficult to determine the ecosystem(s) to which the latter two taxa belong, given that both are found in multiple vegetation communities near to LLS. Moreover, Alchornea is overrepresented in pollen records (Gosling et al., 2009), and Asteraceae includes both herbaceous and arboreal species. One possibility is that the Asteraceae pollen in this zone originates from herbaceous genera in the open savannas (e.g. Gymnocoronis spilanthoides; Orellana et al., 2004). If true, the lower relative proportion of tree/shrub taxa suggests that the local environment in LLS-1 was more open than today, consistent with lacustrine surface-sediment pollen spectra in the open savannas further east (Jones et al., 2011). However, Asteraceae pollen may have also originated from the riparian forests of the Rio Mamoré, given that this vegetation community contains several Asteraceae tree/shrub species. This includes, Tessaria integrifolia, a pioneer shrub that produces monospecific stands adjacent to the river shoreline, as well as on the beaches of old river channels (Maldonado and Beck, 2004; Sanjinés and Beck, 2004). If the Asteraceae pollen in this zone did originate predominantly from arboreal species, then the percentage forest cover in this zone would be comparable to that of LLS-3. Given this uncertainty over the source(s) of Asteraceae pollen, we plot two alternate tree/shrub pollen sum curves in Figure 3 (i.e. with/without Asteraceae included).
Despite our uncertainty over the extent to which the floristic composition of LLS-1 differed from that of the modern landscape, the presence of Zea mays pollen, and its absence from most of LLS-3, provides clear evidence that land-use practices at this time were quite different from those of today. Zea mays pollen is large (55+ μm diameter) and rarely travels more than several hundred metres from its source plant (Clark, 2007), meaning that maize is typically underrepresented within lake sediments (Lane et al., 2010). Its consistent presence throughout LLS-1 therefore suggests that this crop was grown locally. The single Cucurbita grain also implies that squash was cultivated nearby. If we assume that the Asteraceae pollen signal within our record is indicative of herbaceous species, we infer a minor reduction in the proportion of tree/shrub taxa between 150 and 300 CE. However, this shift is mainly driven by the increased pollen abundance of herbaceous taxa (see Figure S6). Alchornea is the only arboreal taxon to show a reduction and is overrepresented in pollen records, raising doubt regarding its contribution to local forest cover (Gosling et al., 2009; Jones et al., 2011). There is also little evidence to attribute this Alchornea decline to anthropogenic deforestation, given that disturbance indicators, such as Cecropia, are absent. The dominance of herbaceous, rather than woody, charcoal particles is indicative of savanna burning, as opposed to the use of fire for forest clearance.
LLS-2 (42.5–30 cm; 590–840 cal. yr CE)
A shift in floristic composition takes place between LLS-1 and LLS-2. While our CONISS clustering algorithm indicates that LLS-2 still exhibits significant differences from the modern vegetation community, our pollen PCA (Figure 5) shows that it is far more floristically similar to it than that of LLS-1. The most notable difference between the pollen assemblages of LLS-1 and LLS-2 is the replacement of Asteraceae by Alternanthera pollen, respectively. As the latter taxon commonly grows around open savanna lakes, but remains absent from the whitewater riparian vegetation communities of the Rio Mamoré (Maldonado and Beck, 2004; Sanjinés and Beck, 2004), this compositional shift may reflect the encroachment of savanna taxa towards LLS. However, the driver(s) underlying this shift remain unknown, as no marked changes occur in either charcoal concentrations or geochemical composition between the two zones. It is also unclear why Alternanthera would be so much more abundant than today, given that LLS is situated close to the Rio Ibare, which drains the same floodwaters feeding the open savanna lakes.
Charcoal concentrations and maize abundances throughout this zone are similar to those of LLS-1, suggesting that no marked changes took place with respect to human population density or land-use practices, despite the shift in floristic composition. We do record a small reduction in the proportion of tree/shrub pollen taxa if we assume the Asteraceae pollen within our record is primarily arboreal. However, this reduction is driven entirely by Asteraceae and Alchornea, and taxa characteristic of forest disturbance (e.g. Cecropia) remain rare, providing little evidence for large-scale anthropogenic deforestation. If we were instead to treat the Asteraceae pollen as herbaceous, the proportion of tree/shrub taxa remains similar to that of LLS-1, consistent with the interpretation that the vegetation around LLS was more open during this period than it is today (Jones et al., 2011).
LLS-3 (30–0 cm; 840 cal. yr CE–present)
The transition between LLS-2 and LLS-3 is marked by significant changes in both land cover and land-use. Although our pollen PCA indicates that the two zones are floristically similar, Zea mays pollen last appears at the base of LLS-3, suggesting the cessation of maize cultivation around LLS. LLS-3 is further distinguished by a doubling of tree/shrub pollen percentages compared with LLS-2 (from 15% to 30%), signalling increased tropical forest coverage around the lake. Given the concurrent termination of maize cultivation, this rise in tree/shrub taxa may reflect forest recovery following the cessation of anthropogenic disturbance. This disturbance could have manifested through direct forest intervention (e.g. slash-and-mulch agriculture; Denevan, 2001) or by preventing natural forest encroachment, as argued for other palaeoecological records near archaeological sites elsewhere in the LM (e.g. Laguna El Cerrito and Laguna Granja; Carson et al., 2014; Whitney et al., 2014). However, as the arboreal pollen sum increase also broadly coincides with a shift towards a wetter climate in the Cuica cave record (Figure 3), associated with onset of the Little Ice Age (Della Libera et al., 2022), it is possible that it also signifies a response to increasing precipitation.
We interpret the peak in Cecropia pollen in LLS-3 to be consistent with repeated natural disturbances associated with the highly dynamic rivers maintaining these riparian floodplain forests in the early stages of succession, as observed today (Maldonado and Beck, 2004; Navarro and Maldonado, 2002). Similar trends in Cecropia have also been recorded at other lake sites in the LM (Carson et al., 2014; Whitney et al., 2014). However, Cecropia pollen also often signifies anthropogenic land use (Bush et al., 2007, 2021), and we cannot entirely rule this out at LLS, given the surrounding forests experienced low-intensity human intervention during the colonial era (Parada, 2013).
Discussion
Maize cultivation at Laguna Loma Suarez
The consistent presence of Zea mays throughout LLS-1 and LLS-2 indicates that humans inhabited and cultivated maize in the local environment around LLS from at least 280 BCE up until approximately 1130 CE. This age-range places the site well within the time frame of the Casarabe Culture. It also aligns with the ubiquitous presence of maize recorded at other Casarabe Culture sites to the east, distant from major rivers (Bruno, 2010, 2015; Dickau et al., 2012; Lombardo et al., 2025; Whitney et al., 2013), and suggests that they grew this crop in a diverse range of environmental settings. In the majority of samples where maize pollen was found, we only recorded a single grain. While this means we can be confident that the crop was grown locally due to the limited dispersal range of its pollen grains (Lane et al., 2010), maize may not have been grown in the same way, or at the same scale, as at these other sites. It has been shown that the Casarabe Culture used a sophisticated network of artificial canals and ponds to practice large-scale maize-based agriculture in the open savannas to the east (Lombardo et al., 2012, 2025), but no such canal network has been found within the local vicinity of LLS. It is possible that, given these stark contrasts in natural resource availability (presence/absence of a major river) between LLS and the central core region of the Casarabe Culture, quite different strategies were used to cultivate maize around our study site; for example, polyculture agroforestry in the riparian forests (Iriarte et al., 2020) or flood-recession agriculture on the seasonally-exposed banks of the nearby rivers (Denevan, 1996; Erickson, 2008). They could have even practiced slash-and-burn maize cultivation in the forests, similar to modern indigenous groups, though this strategy appears unlikely given that they had no access to metal tools, and even inefficient stone tools were extremely rare due to the absence of rock outcrops in the LM (Denevan, 1992a, 2001; Holmberg, 1969; Piland, 1991). The single, poorly preserved Cucurbita grain also suggests that squash may have been cultivated locally, but this cannot be confirmed; while no wild members of Cucurbita were found in local vegetation inventories (Navarro and Maldonado, 2002; Orellana et al., 2004), the LM is home to the closest wild member to domesticated squash (Cucurbita maxima; Lombardo et al., 2020).
Loma Suarez is located to the west of the fertile Late Holocene lobe of alluvium on top of which most of the Casarabe Culture monumental mounds and canals are concentrated (Figure 1; Lombardo et al., 2012). However, its proximity to the Rio Mamoré – a major whitewater river that transports 64 × 106 tonnes of mineral-rich Andean sediment annually (Guyot et al., 1999) – means that its inhabitants had ready access to an alternative source of rich soils for agriculture, utilising its seasonal flood deposits (Bourrel et al., 2004). This river also regularly backflows into the channel of the Rio Ibare (Bourrel et al., 2009), which runs just 350 m from LLS. Thus, while the soils of the wider LM beyond these alluvial deposits are generally unsuitable for maize cultivation (Lombardo et al., 2015), those around LLS were ideal for maize-based agriculture – whether via agroforestry in the riparian forests, or flood-recession agriculture on the exposed river banks. Proximity to both rivers means that it is also highly likely that the inhabitants of Loma Suarez supplemented this maize with fish protein as an important dietary component (Lauzanne and Loubens, 1985), an interpretation reinforced by the presence of fish remains at other mound sites in the LM (Prestes-Carneiro et al., 2019).
It is noteworthy that Zea mays pollen was found within the lowermost pollen sample analysed, dated to 280 ± 208 BCE. Although maize has been cultivated in the LM since at least 6850 BP (Lombardo et al., 2020), this date still predates the onset of mound construction (400 CE; Jaimes Betancourt, 2012, 2015) by nearly seven centuries. Other evidence for maize cultivation preceding initial mound construction comes from LSJ – close to Lomas Mendoza and Salvatierra – where maize was cultivated since at least 180 CE (Whitney et al., 2013). However, of the 189 settlement mounds that have so far been identified (Prümers et al., 2022), only two (Loma Mendoza and Loma Salvatierra) have robust radiocarbon chronologies (Jaimes Betancourt, 2012, 2015). It is therefore plausible that the Casarabe Culture was modifying the local environment around LLS for maize cultivation significantly before 400 CE.
Anthropogenic burning
The relatively stable trends in charcoal concentrations indicate that fire played a consistent role in the local vegetation communities around LLS over the past 2.4 millennia. This stability persists despite a shift in floristic composition being recorded by our pollen PCA, signifying that floristic turnover had little influence over local fire activity during this period. Regional spelaeothem records (Apaéstegui et al., 2018; Novello et al., 2016), the nearest of which is Cuica cave 500 km to the northeast (Figure 1; Della Libera et al., 2022), also document multiple shifts in climate over the past two millennia, but the absence of any corresponding change in charcoal abundance suggests that fire activity was primarily driven by human activity rather than climate. Fire plays a major role in the cultivation practices of many modern Amazonian indigenous groups (Denevan, 2001; Levis et al., 2018), and the sudden increase in charcoal concentration between 150 and 450 CE at LLS broadly aligns with the earliest known mound occupation dates, pointing to a human-fire linkage (400 CE; Jaimes Betancourt, 2012, 2015). Assuming that the LLS charcoal record is largely driven by human activity, then the overwhelming dominance of herbaceous charcoal particles (Figure 3) shows that the Casarabe Culture primarily utilised fire in the open savannas, rather than in the riparian forests. This corroborates findings by Whitney et al. (2013) from the much larger LSJ in the heart of the Casarabe Culture to the east.
However, despite the consistent inferences of savanna burning at LLS and LSJ, the charcoal records do differ between these two sites; vis-a-vis the relatively constant values at LLS between 400 and 900 CE, whereas LSJ shows a progressive increase in charcoal abundance over the same period (Whitney et al., 2013). As airborne charcoal particles larger than 125 μm rarely travel more than 7 km from their source (Whitlock and Larsen, 2001), and these two lakes are more than 50 km apart, this divergence may reflect spatial differences in the Casarabe Culture’s utilisation of fire.
Pre-Columbian deforestation and landscape impact
While our pollen record shows that the Casarabe Culture regularly practiced maize cultivation around LLS, it remains unclear precisely where that cultivation took place. The inhabitants of Loma Suarez could have grown maize in the open savannas, as practiced ca. 50 km to the east in the heart of the Casarabe Culture region (Lombardo et al., 2025) and by the raised-field culture to the north (Whitney et al., 2014), or in the forests, either through slash-and-burn agriculture or as part of a mixed agroforestry system (Levis et al., 2018). Growing maize within the local riparian forests would have forced the Casarabe Culture to practice at least selective deforestation, as the crop is shade intolerant (Gao et al., 2020). However, the only perceptible reduction in tree/shrub pollen observed in the LLS record, which occurs between 150 and 500 CE, is primarily driven by changes in Alchornea and Asteraceae and assumes that the latter is derived from arboreal taxa. Alchornea is overrepresented in southwestern Amazonian pollen records (Gosling et al., 2009; Jones et al., 2011) but accounts for, at most, just 12% of the overall pollen assemblage, implying that it contributes little to local forest cover. Meanwhile, Asteraceae possesses both herbaceous and arboreal species in multiple vegetation communities near to LLS today (Hanagarth, 1993; Navarro and Maldonado, 2002). Thus, while this reduction in tree/shrub pollen overlaps with the earliest known period of mound occupation (Jaimes Betancourt, 2012, 2015) and could therefore potentially be linked with human activity, there is no conclusive evidence to indicate that the Casarabe Culture practiced significant local deforestation around LLS during the past 2400 years.
Assuming that the Asteraceae pollen within our record is primarily herbaceous, the tree/shrub pollen sum in zones LLS-1 and LLS-2 is markedly lower than that of LLS-3. This reduced level more closely resembles the modern pollen rain of lakes in the open savannas (Jones et al., 2011), implying that the local environment around LLS was more open during this time. Given that the southeastern LM has been occupied by humans since the early Holocene (Lombardo et al., 2013) and that we find evidence of maize cultivation in our lowermost pollen sample, it is possible that humans deforested the local landscape around LLS before our record began at 300 BCE, with the Casarabe Culture subsequently maintaining the open environment. Under this scenario, the doubling of tree/shrub pollen percentages at the onset of zone LLS-3 could signify forest recovery following the demise/collapse of the Casarabe Culture. However, it is also plausible that no prior forest clearance took place. As with Laguna Granja and Laguna Oricore in the eastern LM (Carson et al., 2014, 2016), the Casarabe Culture may have settled and constructed Loma Suarez in a relatively open savanna landscape, which has since become more heavily forested. Nonetheless, regardless of whether reduced tree cover was the product of anthropogenic deforestation, the relative proportion of tree/shrub taxa remained constant between 200 BCE and 1000 CE. This implies that, while the Casarabe Culture might have maintained a locally open landscape, it also did not expand the scale of deforestation during this period.
We cannot exclude the possibility that climate change may have played a role in the forest expansion recorded in zone LLS-3, given that it broadly aligns (at least in part) with the transition to a wetter climate at Cuica cave (Figure 3; Della Libera et al., 2022), as well as in other regional palaeoclimate records (Apaéstegui et al., 2018; Novello et al., 2016) associated with the onset of the Little Ice Age. However, forest-savanna responses to climate change in these seasonally-flooded landscapes are likely to be complex and may in fact be opposite to expectation; perhaps accounting for the lack of more robust vegetation-climate linkages. For example, while higher rainfall often favours rainforest expansion in terra firme ecotonal landscapes (Burbridge et al., 2004; Carson et al., 2014; Mayle et al., 2000), increased precipitation in the seasonally-flooded LM may instead promote the expansion of open savanna, given the intolerance of woody plants to prolonged flooding (Mayle et al., 2007). Instead, drier climatic conditions (i.e. reduced inundation) may favour the replacement of seasonally-flooded savanna with forest, as has been hypothesised by Mayle et al. (2007) and subsequently supported by phytolith evidence (Lombardo et al., 2019).
Notwithstanding uncertainties over the potential influence of climate change associated with the Mediaeval Climate Anomaly and/or the Little Ice Age, and the attribution of Asteraceae pollen to arboreal versus herbaceous taxa, we argue that the most parsimonious interpretation of the LLS record is the maintenance of a relatively open landscape and maize cultivation by the Casarabe Culture, with subsequent forest encroachment following its collapse and depopulation.
Conclusions
The multiproxy palaeoecological record from Laguna Loma Suarez (LLS) reveals that the pre-Columbian Casarabe Culture practiced maize cultivation not only in savanna-dominated landscapes in the heart of the monumental mound region of Amazonian Bolivia – where maize formed the basis of an agrarian economy (Bruno, 2010, 2015; Dickau et al., 2012; Lombardo et al., 2025; Whitney et al., 2013) – but also in riparian forest settings, where we expected aquatic and riparian food resources to take precedence. This does not imply that they employed the same agricultural practices as at these other sites; indeed, differences in the patterns of anthropogenic burning between LLS and Laguna San Jose (LSJ; Whitney et al., 2013) indicate that there were spatial variations in how the Casarabe Culture utilised the forest-savanna mosaic landscapes of the Bolivian Llanos de Moxos (LM). Nonetheless, there are some key similarities between these sites, such as anthropogenic fire use being primarily restricted to the open savannas.
Notwithstanding the highly variable distribution of maize pollen grains (Lane et al., 2010), their disappearance alongside an increase in tree/shrub pollen after 1130 cal. CE strongly suggests that the Casarabe Culture maintained a more open landscape around LLS, by preventing forest growth, until after the nearby settlement mound (Loma Suarez) was abandoned. This is consistent with findings in quite different environmental settings at the opposite (eastern) end of the LM (Carson et al., 2014, 2016).
Although we attribute forest expansion after 1130 cal. CE largely to forest recovery following settlement site abandonment, it is possible that climate change may have been a contributing driver (Della Libera et al., 2022). However, the mechanism by which this occurred in this seasonally-flooded mosaic landscape is by no means clear (Lombardo et al., 2019; Mayle et al., 2007).
To conclude, we find that maize was an important cultivar in pre-Columbian times across the seasonally-flooded savannas of Amazonian Bolivia, despite major differences in environmental setting (fluvial riparian forest vs open savanna wetland vs terra firme forest/savanna) and different types of culture (Monumental Mound vs Ring Ditch (Carson et al., 2014, 2016) vs raised-field cultures (Whitney et al., 2014)).
Supplemental Material
sj-docx-1-hol-10.1177_09596836251332794 – Supplemental material for Localised land-use and maize agriculture by the pre-Columbian Casarabe Culture in Lowland Bolivia
Supplemental material, sj-docx-1-hol-10.1177_09596836251332794 for Localised land-use and maize agriculture by the pre-Columbian Casarabe Culture in Lowland Bolivia by Joseph Hirst, Marco Raczka, Umberto Lombardo, Ezequiel Chavez, Lorena Becerra-Valdivia, McKenzie Bentley, Christopher Bronk Ramsey, Miros Stavros James Charidemou, Suzanne Maclachlan and Francis E Mayle in The Holocene
Footnotes
Acknowledgements
We thank the following: the Museo de Historia Natural ‘Noel Kempff Mercado’, Santa Cruz, Bolivia, for logistical support during fieldwork; two anonymous reviewers whose useful comments improved the manuscript; Ian Croudace for his help with interpretation of the XRF data.
Author contribution(s)
Declaration of conflicting interests
The authors declare that Francis Mayle serves on the editorial board for The Holocene and therefore request another editorial board member is chosen to manage the review of this manuscript. There are no other potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Funding
The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This research was funded by the SCENARIO NERC doctoral training partnership grant (NE/S007261/1). Funding for the 14 radiocarbon dates was provided by the ‘Human Environment Relationships in pre-Columbian Amazonia’ (HERCA) project (AHRC grant number: AH/S001662/1: FAPESP grant number: 2019/07794-9; co-PIs: F. Mayle, E.G. Neves; co-I: C. Ramsey).
Ethical considerations
It was not necessary to obtain ethical approval in order to conduct this research.
Data availability
Supplemental material
Supplemental material for this article is available online.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.